A Mass Spectrometry-Based Study Shows that Volatiles Emitted by Arthrobacter agilis UMCV2 Increase the Content of Brassinosteroids in Medicago truncatula in Response to Iron Deficiency Stress


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
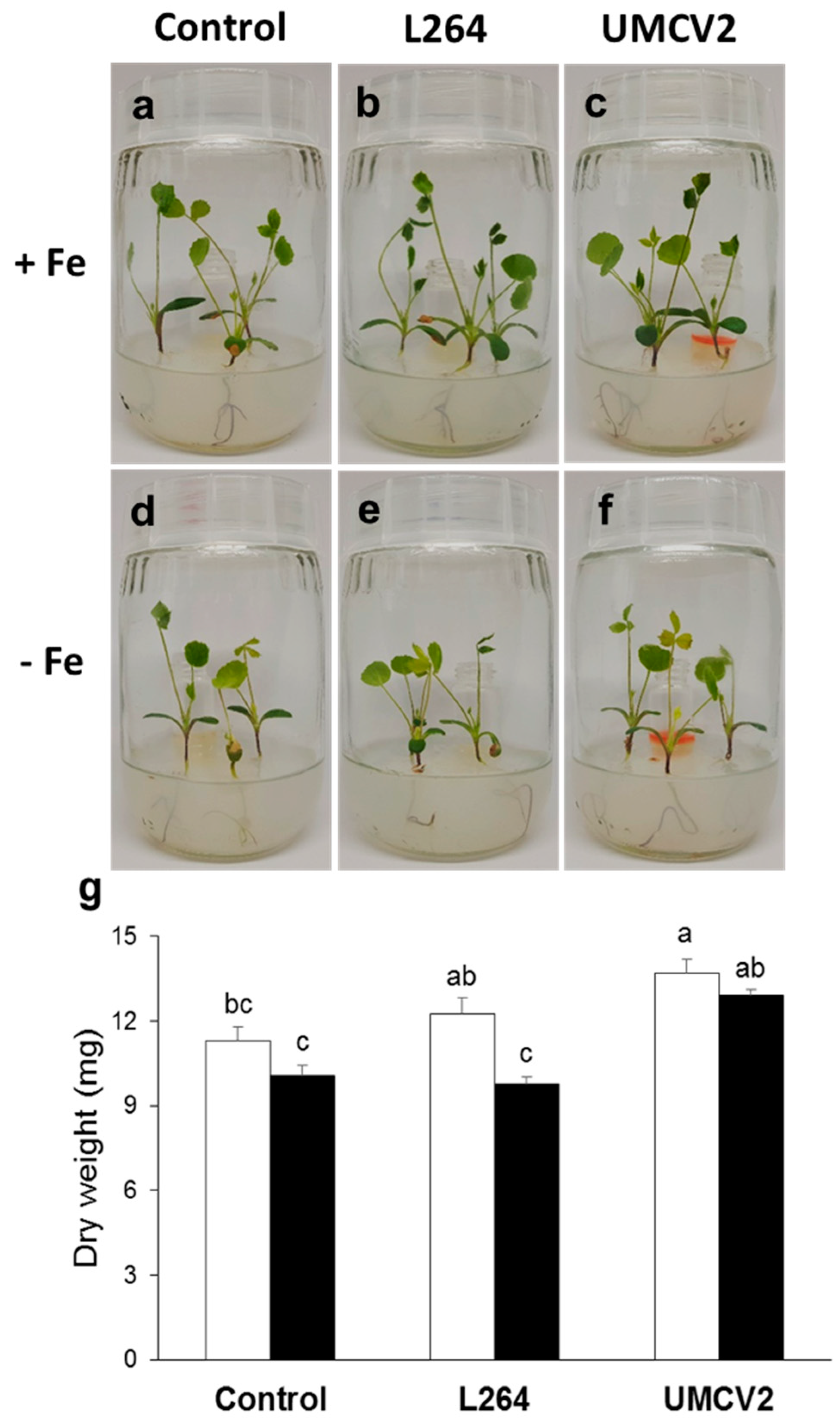
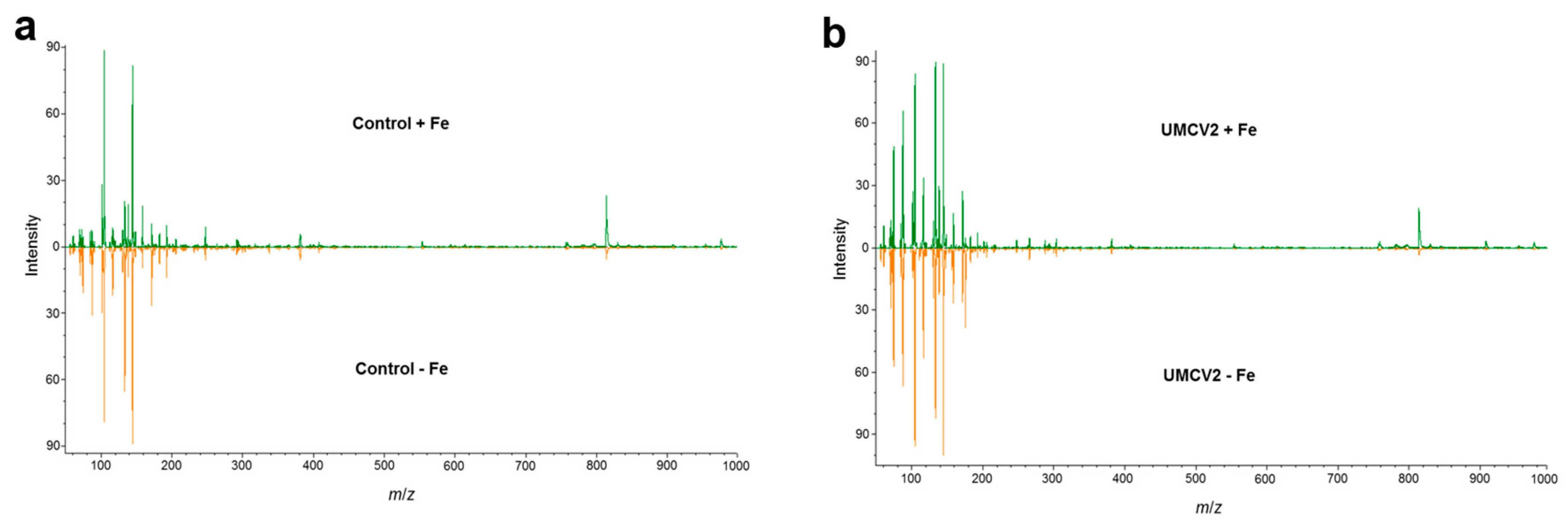
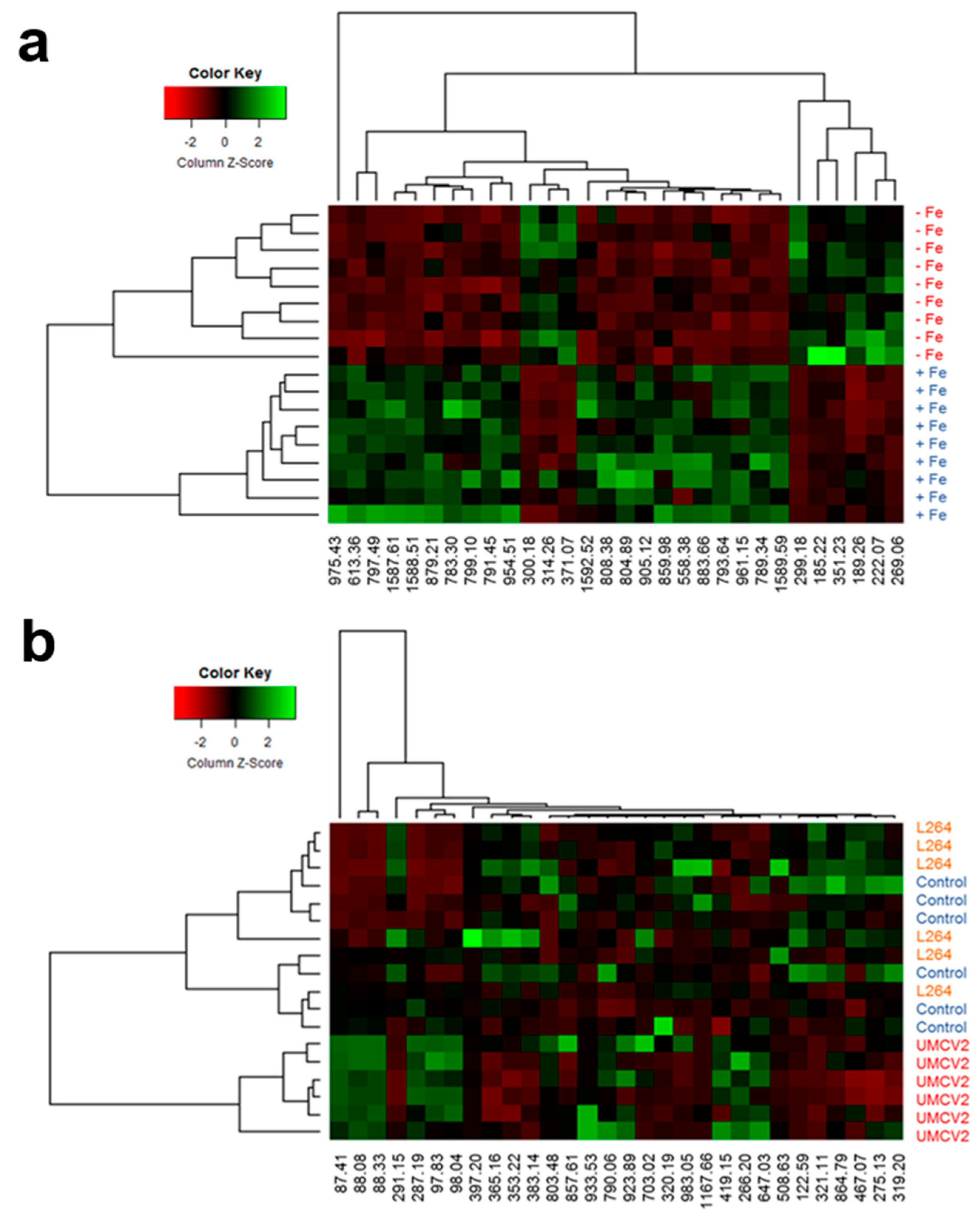
2.1. DLI-ESI-MS in the Analysis of Fe Deficiency in Medicago truncatula Seedlings and Response to Bacterial Volatiles
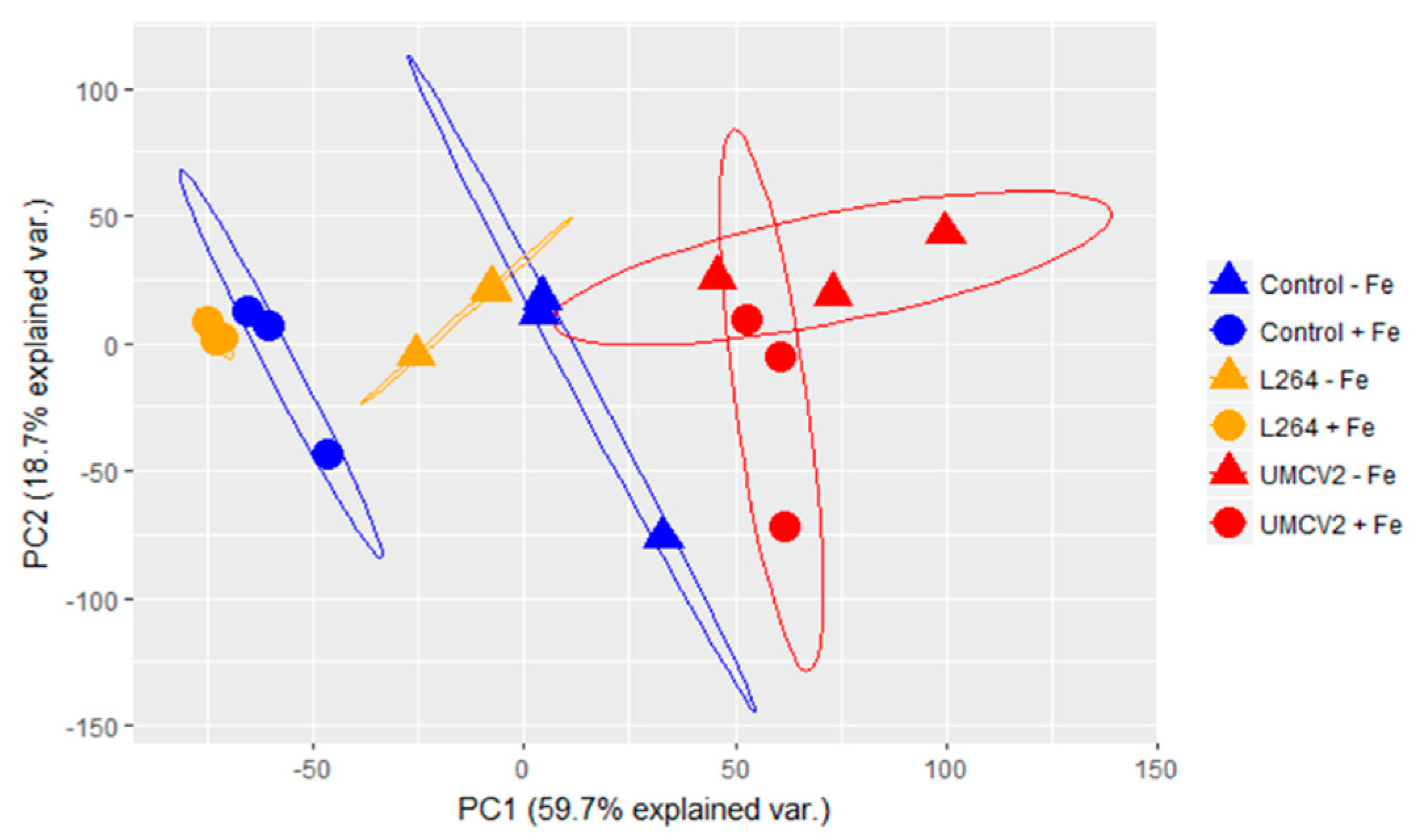
2.2. Classification of Random Forest Model for Differentiating Plants Grown under Two Different Fe Conditions and Rhizobacterial Inoculation
3. Discussion
4. Materials and Methods
4.1. Biological Material and Growth Conditions
4.2. Plant-Bacteria Interaction through the Emission of Volatiles
4.3. Metabolite Extraction from M. truncatula Leaves
4.4. Non-Targeted Metabolic Profiling by DLI-ESI Mass Spectrometry
4.5. Identification of Significant Ions
4.6. Brassinosteroids Determination
Author Contributions
Funding
Conflicts of Interest
References
- Analytical Methods Committee AMCTB No. 81. A “periodic table” of mass spectrometry instrumentation and acronyms. Anal. Methods 2017, 9, 5086–5090.
- García-Flores, M.; Juárez-Colunga, S.; García-Casarrubias, A.; Trachsel, S.; Winkler, R.; Tiessen, A. Metabolic profiling of plant extracts using direct-injection electrospray ionization mass spectrometry allows for high-throughput phenotypic characterization according to genetic and environmental effects. J. Agric. Food Chem. 2015, 63, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Becerra, R.; Montero-Vargas, J.; Martínez-Jarquín, S.; Gálvez-Ponce, E.; Moreno-Pedraza, A.; Winkler, R. Rapid classification of coffee products by data mining models from direct electrospray and plasma-based mass spectrometry analyses. Food Anal. Methods 2016, 10, 1359–1368. [Google Scholar] [CrossRef]
- González-Domínguez, R.; Sayago, A.; Fernández-Recamales, A. High-throughput mass-spectrometry based-metabolomics to characterize metabolite fingerprints associated with Alzheimer’s disease pathogenesis. Metabolites 2018, 8, 52. [Google Scholar] [CrossRef] [PubMed]
- Montero-Vargas, J.; Casarrubias-Castillo, K.; Martínez-Gallardo, N.; Ordaz-Ortiz, J.; Délano-Frier, J.; Winkler, R. Modulation of steroidal glycoalkaloid biosynthesis in tomato (Solanum lycopersicum) by jasmonic acid. Plant. Sci. 2018, 277, 155–165. [Google Scholar] [CrossRef] [PubMed]
- García-Flores, M.; Juárez-Colunga, S.; Montero-Vargas, J.M.; López-Arciniega, J.A.I.; Chagolla, A.; Tiessen, A.; Winkler, R. Evaluating the physiological state of maize (Zea mays L.) plants by direct-injection electrospray mass spectrometry (DIESI-MS). Mol. BioSyst. 2012, 8, 1658–1660. [Google Scholar] [CrossRef] [PubMed]
- Rendón-Anaya, M.; Montero-Vargas, J.M.; Saburido-Álvarez, S.; Vlasova, A.; Capella-Gutierrez, S.; Ordaz-Ortiz, J.J.; Aguilar, O.M.; Vianello-Brondani, R.P.; Santalla, M.; Delaye, L.; et al. Genomic history of the origin and domestication of common bean unveils its closest sister species. Genome Biol. 2017, 18, 60. [Google Scholar] [CrossRef]
- González-Vallejo, E.B.; Susín, S.; Abadía, A.; Abadía, J. Changes in sugar beet leaf plasma membrane Fe(III)-chelate reductase activities mediated by Fe-deficiency, assay buffer composition, anaerobiosis and the presence of flavins. Protoplasma 1998, 205, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Salama, Z.; El-Beltagi, H.; El-Hariri, D.M. Effect of Fe deficiency on antioxidant system in leaves of three flax cultivars. Not. Bot. Hort. Agrobot. Cluj. 2009, 37, 122–128. [Google Scholar]
- Jin, C.W.; Li, G.X.; Yu, X.H.; Zheng, S.J. Plant Fe status affects the composition of siderophore-secreting microbes in the rhizosphere. Ann. Bot. 2010, 105, 835–841. [Google Scholar] [CrossRef]
- Rodríguez-Celma, J.; Lattanzio, G.; Grusak, M.A.; Abadía, A.; Abadía, J.; López-Millán, A.F. Root responses of Medicago truncatula plants grown in two different iron deficiency conditions: Changes in root protein profile and riboflavin biosynthesis. J. Proteome Res. 2011, 10, 2590–2601. [Google Scholar] [CrossRef] [PubMed]
- Fariduddin, Q.; Varshney, P.; Yusuf, M.; Ahmad, A. Polyamines: Potent modulators of plant responses to stress. J. Plant. Interact. 2013, 8, 1–16. [Google Scholar] [CrossRef]
- Ravet, K.; Pilon, M. Copper and iron homeostasis in plants: The challenges of oxidative stress. Antioxid Redox Signal. 2013, 19, 919–932. [Google Scholar] [CrossRef]
- Baba, S.A.; Malik, A.H.; Wani, Z.A.; Mohiuddin, T.; Shah, Z.; Abbas, N.; Ashraf, N. Phytochemical analysis and antioxidant activity of different tissue types of Crocus sativus and oxidative stress alleviating potential of saffron extract in plants, bacteria, and yeast. S. Afr. J. Bot. 2015, 99, 80–87. [Google Scholar] [CrossRef]
- Wang, B.; Li, G.; Zhang, W.H. Brassinosteroids are involved in Fe homeostasis in rice (Oryza sativa L.). J. Exp. Bot. 2015, 66, 2749–2761. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Lan, P. The understanding of the plant iron deficiency responses in strategy I plants and the role of ethylene in this process by omic approaches. Front. Plant. Sci. 2017, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.D.R.; Barros Junior, U.O.; Batista, B.L.; da Silva Lobato, A.K. Brassinosteroids mitigate iron deficiency improving nutritional status and photochemical efficiency in Eucalyptus urophylla plants. Trees 2018, 32, 1681–1694. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; Alías-Villegas, C.; Bellogín, R.A.; del Cerro, P.; Espuny, M.R.; Jiménez-Guerrero, I.; López-Baena, F.J.; Ollero, F.J.; Cubo, T. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Pare, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. 2003, 100, 4927–4932. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, Y.; Xie, X.; Kim, M.S.; Dowd, S.E.; Paré, P.W. A soil bacterium regulates plant acquisition of iron via deficiency-inducible mechanisms. Plant. J. 2009, 58, 568–577. [Google Scholar] [CrossRef]
- Valencia-Cantero, E.; Hernández-Calderón, E.; Velázquez-Becerra, C.; López-Meza, J.E.; Alfaro-Cuevas, R.; López-Bucio, J. Role of dissimilatory fermentative iron-reducing bacteria in Fe uptake by common bean (Phaseolus vulgaris L.) plants grown in alkaline soil. Plant. Soil. 2007, 291, 263–273. [Google Scholar] [CrossRef]
- Velázquez-Becerra, C.; Macías-Rodríguez, L.I.; López-Bucio, J.; Altamirano-Hernández, J.; Flores-Cortez, I.; Valencia-Cantero, E. A volatile organic compound analysis from Arthrobacter agilis identifies dimethylhexadecylamine, an amino-containing lipid modulating bacterial growth and Medicago sativa morphogenesis in vitro. Plant. Soil. 2011, 339, 329–340. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.C.; Velázquez-Becerra, C.; Macías-Rodríguez, L.I.; Santoyo, G.; Flores-Cortez, I.; Alfaro-Cuevas, R.; Valencia-Cantero, E. Arthrobacter agilis UMCV2 induces iron acquisition in Medicago truncatula (strategy I plant) in vitro via dimethylhexadecylamine emission. Plant. Soil. 2013, 362, 51–66. [Google Scholar] [CrossRef]
- Castulo-Rubio, D.Y.; Alejandre-Ramírez, N.A.; Orozco-Mosqueda, M.C.; Santoyo, G.; Macías-Rodríguez, L.I.; Valencia-Cantero, E. Volatile organic compounds produced by the rhizobacterium Arthrobacter agilis UMCV2 modulate Sorghum bicolor (strategy II plant) morphogenesis and SbFRO1 transcription in vitro. J. Plant. Growth Regul. 2015, 34, 611–623. [Google Scholar] [CrossRef]
- Montejano-Ramírez, V.; Martínez-Cámara, R.; García-Pineda, E.; Valencia-Cantero, E. Rhizobacterium Arthrobacter agilis UMCV2 increases organ-specific expression of FRO genes in conjunction with genes associated with the systemic resistance pathways of Medicago truncatula. Acta Physiol. Plant. 2018, 40, 138. [Google Scholar] [CrossRef]
- Shimojima, M.; Ohta, H. Critical regulation of galactolipid synthesis controls membrane differentiation and remodeling in distinct plant organs and following environmental changes. Prog. Lipid Res. 2011, 50, 258–266. [Google Scholar] [CrossRef]
- Aronsson, H.; Schottler, M.A.; Kelly, A.A.; Sundqvist, C.; Dormann, P.; Karim, S.; Jarvis, P. Monogalactosyldiacylglycerol deficiency in Arabidopsis affects pigment composition in the prolamellar body and impairs thylakoid membrane energization and photoprotection in leaves. Plant. Physiol. 2008, 148, 580–592. [Google Scholar] [CrossRef]
- McNeil, S.D.; Nuccio, M.L.; Ziemak, M.J.; Hanson, A.D. Enhanced synthesis of choline and glycine betaine in transgenic tobacco plants that overexpress phosphoethanolamine N-methyltransferase. Proc. Natl. Acad. Sci. 2001, 98, 10001–10005. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Aly, A.A. Iron Deficiency Stimulated Some Enzymes Activity, Lipid Peroxidation and free radicals production in Borage officinalis induced in vitro. Int. J. Agri. Biol. 2004, 6, 179–184. [Google Scholar]
- Sperotto, R.A.; Boff, T.; Duarte, G.L.; Fett, J.P. Increased senescence-associated gene expression and lipid peroxidation induced by iron deficiency in rice roots. Plant. Cell Rep. 2008, 27, 183–195. [Google Scholar] [CrossRef]
- Vigani, G.; Zocchi, G. The fate and the role of mitochondria in Fe-deficient roots of Strategy I plants. Plant. Signal. Behav. 2009, 4, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitrián, M.; Zarza, X.; Altabella, T.; Tiburcio, A.F.; Alcázar, R. Polyamines under abiotic stress: Metabolic crossroads and hormonal crosstalks in plants. Metabolites 2012, 2, 516–528. [Google Scholar] [CrossRef]
- Voss, I.; Sunil, B.; Scheibe, R.; Raghavendra, A.S. Emerging concept for the role of photorespiration as an important part of abiotic stress response. Plant. Biol. 2013, 15, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, R.; Zhang, Y.M.; Rock, C.O.; Jackowski, S. Coenzyme A: Back in action. Prog Lipid Res. 2005, 44, 125–153. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Yokota, T. Biosynthesis and metabolism of brassinosteroids. Annu Rev. Plant. Biol. 2003, 54, 137–164. [Google Scholar] [CrossRef] [PubMed]
- Bishop, G.J.; Nomura, T.; Yokota, T.; Harrison, K.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Jones, J.D.G.; Kamiya, Y. The tomato DWARF enzyme catalyses C-6 oxidation in brassinosteroid biosynthesis. Proc. Natl. Acad. Sci. 1999, 96, 1761–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, T.; Sato, T.; Bishop, G.J.; Kamiya, Y.; Takatsuto, S.; Yokota, T. Acumulation of 6-deoxocathasterone and 6-deoxocatasterone in Arabidopsis, pea and tomato is suggestive of common rate limiting steps in brassinosteroid biosynthesis. Phytochemistry 2001, 57, 171–178. [Google Scholar] [CrossRef]
- Clouse, S.D. Brassinosteroids. Arab. Book 2002, 1, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Goyer, A. Thiamine in plants: Aspects of its metabolism and functions. Phytochemistry 2010, 71, 1615–1624. [Google Scholar] [CrossRef]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant. Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef]
- He, N.W.; Zhao, Y.; Guo, L.; Shang, J.; Yang, X.B. Antioxidant, antiproliferative, and pro-apoptotic activities of a saponin extract derived from the roots of Panax notoginseng (Burk.) F.H. Chen. J. Med. Food. 2012, 15, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Wasson, A.P.; Pellerone, F.I.; Mathesius, U. Silencing the flavonoid pathway in Medicago truncatula inhibits root nodule formation and prevents auxin transport regulation by Rhizobia. Plant. Cell 2006, 18, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Maj, D.; Wielbo, J.; Marek-Kozaczu, M.; Skorupska, A. Response to flavonoids as a factor influencing competitiveness and symbiotic activity of Rhizobium leguminosarum. Microbiol. Res. 2010, 165, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Lateif, K.; Bogusz, D.; Hocher, V. The role of flavonoids in the establishment of plant roots endosymbioses with arbuscular mycorrhiza fungi, rhizobia and Frankia bacteria. Plant. Signal. Behav. 2012, 7, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Meyerowitz, E.M.; Somerville, C.R. Arabidopsis; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1994; p. 1270. [Google Scholar]
- Pollard, M.; Beisson, F.; Li, Y.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant. Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Garay, B.; Phillips, G.C.; Kuehn, G.D. Detection of norspermidine and norspermine in Medicago sativa L. (alfalfa). Plant. Physiol. 1989, 89, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.F.; Wang, B.; Song, W.F.; Zheng, S.J.; Shen, R.F. Putrescine alleviates iron deficiency via NO-dependent reutilization of root cell-wall Fe in Arabidopsis. Plant. Physiol. 2016, 170, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Bourgaud, F.; Hehn, A.; Larbat, R.; Doerper, S.; Gontier, E.; Kellner, S.; Matern, U. Biosynthesis of coumarins in plants: A major pathway still to be unravelled for cytochrome P450 enzymes. Phytochem. Rev. 2006, 5, 293–308. [Google Scholar] [CrossRef]
- Ingle, R.A. Histidine biosynthesis. Arab. Book 2011, 9, 1–9. [Google Scholar] [CrossRef]
- Velazquez, E.; Silva, L.R.; Peix, A. Legumes: A healthy and ecological source of flavonoids. Curr. Nutr. Food Sci. 2010, 6, 109–144. [Google Scholar] [CrossRef]
- Wang, B.; Li, Y.; Zhang, W.H. Brassinosteroids are involved in response of cucumber (Cucumis sativus) to iron deficiency. Ann. Bot. 2012, 110, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Boisson-Dernier, A.; Andriankaja, A.; Chabaud, M.; Niebel, A.; Journet, E.P.; Barker, D.G.; de Carvalho-Niebel, F. MtENOD11 Gene activation during rhizobial infection and mycorrhizal arbuscule development requires a common AT-rich-containing regulatory sequence. MPMI 2005, 18, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Luna, F.M.; López-Bucio, J.; Altamirano-Hernández, J.; Valencia-Cantero, E.; Reyes-de la Cruz, H.; Macías-Rodríguez, L. Plant growth-promoting rhizobacteria modulate root-system architecture in Arabidopsis thaliana through volatile organic compound emission. Symbiosis 2010, 51, 75–83. [Google Scholar] [CrossRef]
- Hernández-Calderón, E.; Aviles-Garcia, M.E.; Castulo-Rubio, D.Y.; Macías-Rodríguez, L.; Montejano-Ramírez, V.; Santoyo, G.; López-Bucio, J.; Valencia-Cantero, E. Volatile compounds from beneficial or pathogenic bacteria differentially regulate root exudation, transcription of iron transporters, and defense signaling pathways in Sorghum bicolor. Plant. Mol. Biol. 2018, 96, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Strohalm, M.; Hassman, M.; Košata, B.; Kodíček, M. mMass data miner: An open source alternatative for mass spectrometric data analysis. Rapid Commun. Mass Spectrom. 2008, 22, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Gibb, S.; Strimmer, K. MALDIquant: A versatile R package for the analysis of mass spectrometry data. Bioinformatics 2012, 28, 2270–2271. [Google Scholar] [CrossRef]
- Williams, G.J. Data Mining with Rattle and R: The Art of Excavating Data for Knowledge Discovery; Springer: New York, NY, USA, 2011; p. 374. [Google Scholar]
- Winkler, R. SpiderMass: Semantic database creation and tripartite metabolite identification strategy. J. Mass Spectrom. 2015, 50, 538–541. [Google Scholar] [CrossRef]
- Ikekawa, N.; Takatsuto, S. Microanalysis of brassinosteroids in plants by gas chromatography/mass spectrometry. J. Mass Spectrom. Soc. Jpn. 1984, 32, 55–70. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m/z | Monoisotopic Mass (Da) | Ionization Mode | Compound Name | Function |
|---|---|---|---|---|
| 189.26 | 188.20 | [M + H]+ | Norspermine | Stress |
| 185.22 | 184.01 | [M + H]+ | 1-Deoxy-l-glycero-tetrulose 4-phosphate | Riboflavin biosynthesis |
| 222.07 | 199.03 | [M + Na]+ | l-Histidinol-phosphate | Histidine biosynthesis |
| 269.06 | 246.09 | [M + Na]+ | (+)-Marmesin | Stress |
| 300.18 | 299.15 | [M + H]+ | (S)-N-methylcoclaurine | Stress |
| 613.36 | 612.22 | [M + H]+ | Protochlorophyllide a | Chlorophyll biosynthesis |
| 299.18 | 276.11 | [M + Na]+ | 5-Amino-6-(d-ribitylamino) uracil | Riboflavin biosynthesis |
| 859.98 | 837.16 | [M + Na]+ | Butanoyl-CoA | Fatty acid beta oxidation |
| 797.49 | 774.53 | [M + Na]+ | 1-18:3-2-18:3-Monogalactosyldiacylglycerol | Chloroplast membrane lipid |
| 314.26 | 313.24 | [M + H]+ | 9,10-Epoxy-18-hydroxystearate | Cutin biosynthesis |
| 371.07 | 348.12 | [M + Na]+ | Chelerythrine | Stress |
| 808.38 | 785.16 | [M + Na]+ | Dioleoylphosphatidylcholine | Membranes lipid |
| 351.23 | 328.17 | [M + Na]+ | Crocetin | Stress |
| m/z | Monoisotopic Mass (Da) | Ionization Mode | Compound Name | Function |
|---|---|---|---|---|
| 88.08 | 87.01 | [M + H]+ | Pyruvate | Energy |
| 266.20 | 265.11 | [M + H]+ | Thiamine | Stress |
| 383.14 | 360.14 | [M + Na]+ | 7-Deoxyloganate | Stress |
| 933.53 | 932.53 | [M + H]+ | Notoginsenoside R1 | Stress |
| 319.20 | 296.31 | [M + Na]+ | Phytol | Constituent of chlorophyll |
| 365.17 | 342.12 | [M + Na]+ | Galactinol | Stress |
| 98.34 | 75.07 | [M + Na]+ | N-Monomethylethanolamine | Choline biosynthesis |
| 291.15 | 268.07 | [M + Na]+ | Formononetin | Stress |
| 353.22 | 352.18 | [M + H]+ | 16-Hydroxytabersonine | Indole alkaloid biosynthesis |
| 397.20 | 396.34 | [M + H]+ | 5-Dehydroepisterol | Brassinosteroid biosynthesis |
| 287.19 | 286.05 | [M + H]+ | Kaempferol | Stress |
| 97.83 | 75.01 | [M + Na]+ | Glycolate | Photorespiration |
| 467.07 | 466.37 | [M + H]+ | 6-Hydroxycastasterone | Brassinosteroid biosynthesis |
| 321.11 | 320.09 | [M + H]+ | 4-Coumaroylshikimate | Flavonoid and phenylpropanoid biosynthesis |
| 790.06 | 767.12 | [M + Na]+ | Coenzyme A | Fatty acid beta oxidation |
| 275.13 | 274.08 | [M + H]+ | Fustin | Stress |
| 320.20 | 297.24 | [M + Na]+ | 18-Hydroxyoleate | Cutin, suberin and wax biosynthesis |
| 88.33 | 87.07 | [M + H]+ | 4-Aminobutanal | Stress |
| 87.41 | 86.07 | [M + H]+ | 3-Pentanone | Stress |
| 419.15 | 418.38 | [M + H]+ | 6-Deoxocathasterone | Brassinosteroid biosynthesis |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Cortez, I.; Winkler, R.; Ramírez-Ordorica, A.; Elizarraraz-Anaya, M.I.C.; Carrillo-Rayas, M.T.; Valencia-Cantero, E.; Macías-Rodríguez, L. A Mass Spectrometry-Based Study Shows that Volatiles Emitted by Arthrobacter agilis UMCV2 Increase the Content of Brassinosteroids in Medicago truncatula in Response to Iron Deficiency Stress. Molecules 2019, 24, 3011. https://doi.org/10.3390/molecules24163011
Flores-Cortez I, Winkler R, Ramírez-Ordorica A, Elizarraraz-Anaya MIC, Carrillo-Rayas MT, Valencia-Cantero E, Macías-Rodríguez L. A Mass Spectrometry-Based Study Shows that Volatiles Emitted by Arthrobacter agilis UMCV2 Increase the Content of Brassinosteroids in Medicago truncatula in Response to Iron Deficiency Stress. Molecules. 2019; 24(16):3011. https://doi.org/10.3390/molecules24163011
Chicago/Turabian StyleFlores-Cortez, Idolina, Robert Winkler, Arturo Ramírez-Ordorica, Ma. Isabel Cristina Elizarraraz-Anaya, María Teresa Carrillo-Rayas, Eduardo Valencia-Cantero, and Lourdes Macías-Rodríguez. 2019. "A Mass Spectrometry-Based Study Shows that Volatiles Emitted by Arthrobacter agilis UMCV2 Increase the Content of Brassinosteroids in Medicago truncatula in Response to Iron Deficiency Stress" Molecules 24, no. 16: 3011. https://doi.org/10.3390/molecules24163011
APA StyleFlores-Cortez, I., Winkler, R., Ramírez-Ordorica, A., Elizarraraz-Anaya, M. I. C., Carrillo-Rayas, M. T., Valencia-Cantero, E., & Macías-Rodríguez, L. (2019). A Mass Spectrometry-Based Study Shows that Volatiles Emitted by Arthrobacter agilis UMCV2 Increase the Content of Brassinosteroids in Medicago truncatula in Response to Iron Deficiency Stress. Molecules, 24(16), 3011. https://doi.org/10.3390/molecules24163011