Identification of New Antifungal Agents Targeting Chitin Synthesis by a Chemical-Genetic Method

Abstract
:1. Introduction
2. Results
2.1. To Screen Agents that Are More Toxic to Yeast Glucan Synthase Mutants
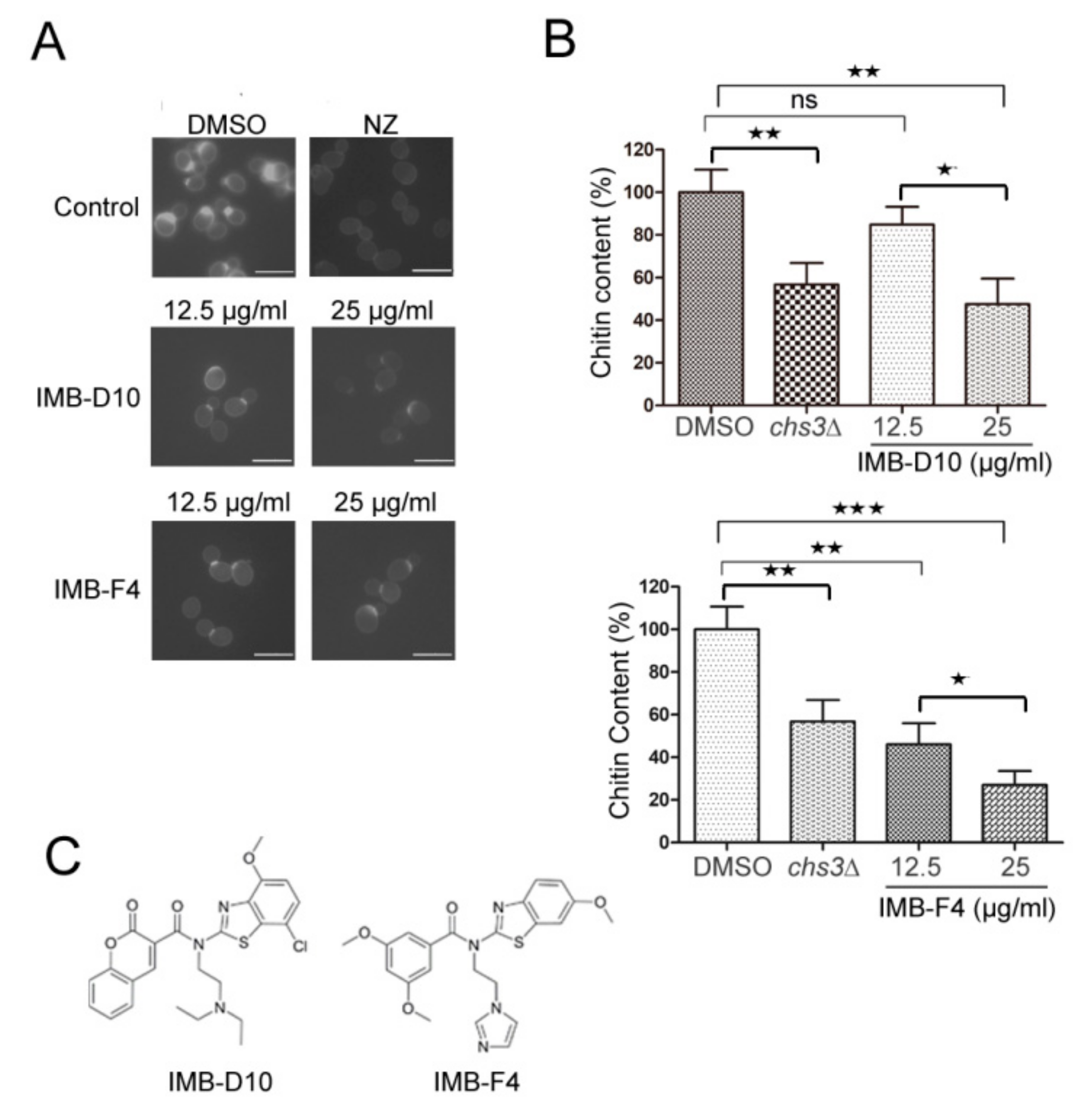
2.2. The Effect of the Candidate Compounds on Yeast Chitin Levels
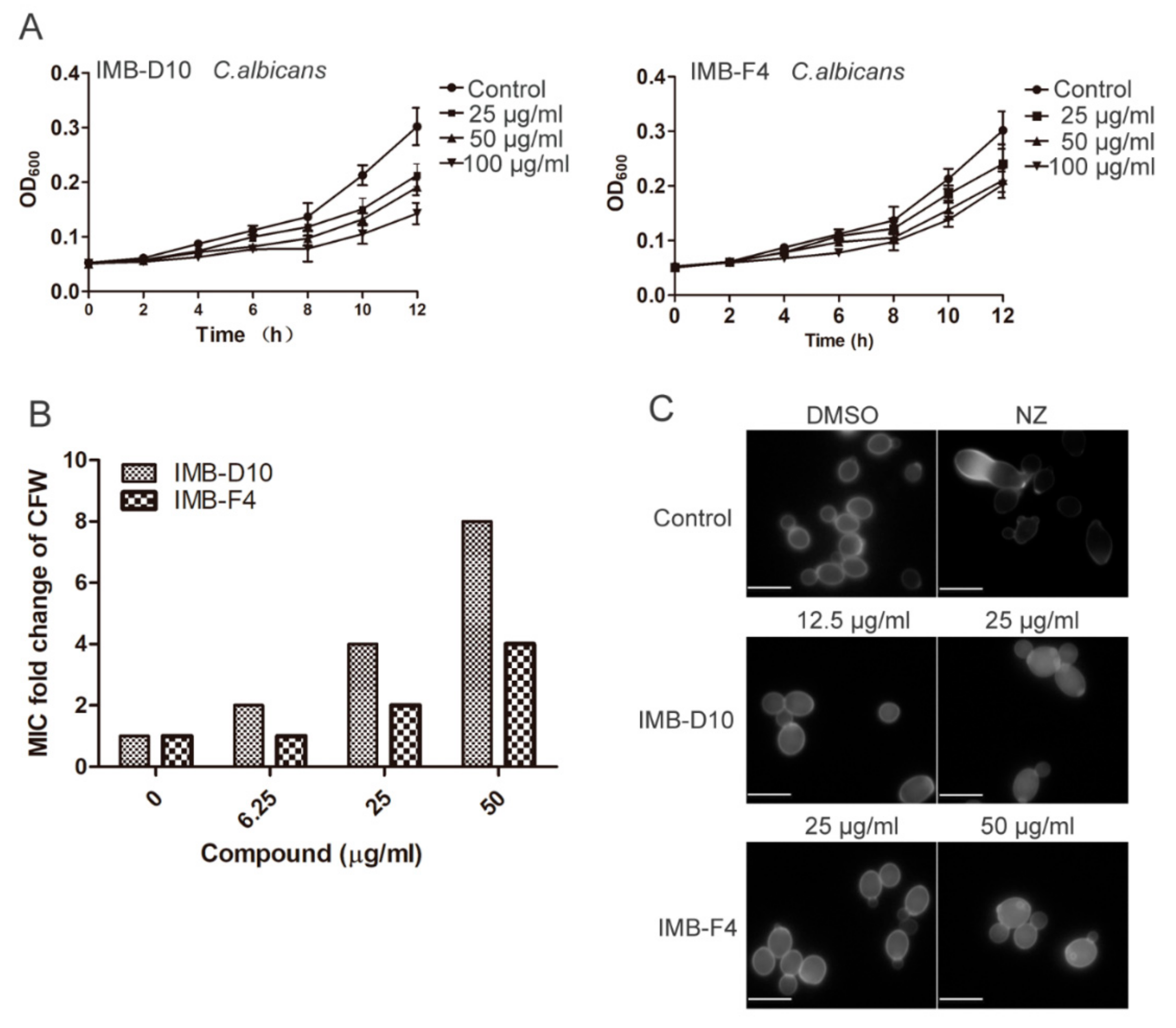
2.3. IMB-D10 and IMB-F4 Inhibit Fungal Growth at High Concentrations without Toxicity to Human Cells
2.4. The Effect of IMB-D10 and IMB-F4 on the Activity of Antifungal Agents Targeting Cell Wall
2.5. Inhibition of Chitin Synthase Activity by IMB-D10 and IMB-F4
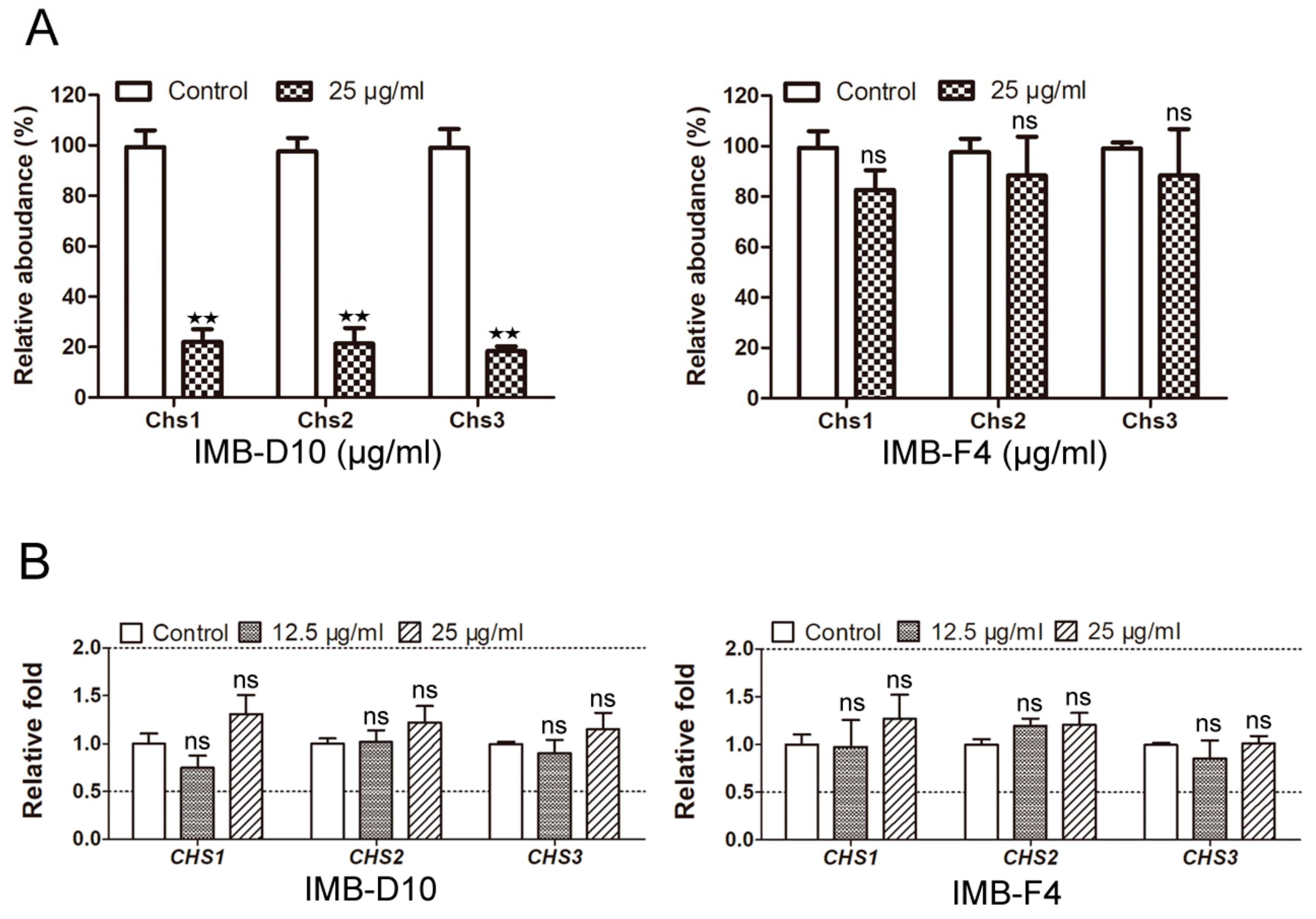
2.6. IMB-D10 Reduces the Protein Level of Three Chitin Synthases
2.7. IMB-D10 and IMB-F4 Inhibit the Growth of Candida albicans
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Compound Library Screening
4.3. Light and Fluorescence Microscopy
4.4. Analysis of Chitin Synthases Activity
4.5. Chitin Content Assay
4.6. Quantitative Real-Time PCR (qRT-PCR)
4.7. Use Isobaric Tags for Relative and Absolute Quantification (iTRAQ) to Determine the Protein Levels of Chitin Synthases
4.8. Analysis of Synergistic and Antagonistic Effect
4.9. Cytotoxicity Assay
4.10. Growth Curve Assay
4.11. The Growth Inhibition of S. cerevisiae and C. albicans by Compounds IMB-D10 and IMB-F4
4.12. Statistic Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Köhler, J.R.; Hube, B.; Puccia, R.; Casadevall, A.; Perfect, J.R. Fungi that Infect Humans. Microbiol. Spectr. 2017, 5, 28597822. [Google Scholar]
- Limper, A.H.; Adenis, A.; Le, T.; Harrison, T.S. Fungal infections in HIV/AIDS. Lancet Infect. Dis. 2017, 17, e334–e343. [Google Scholar] [CrossRef]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Sanglard, D. Emerging threats in antifungal-resistant fungal pathogens. Front. Med. 2016, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Holan, Z.; Pokorný, V.; Beran, K.; Gemperle, A.; Tuzar, Z.; Baldrián, J. The glucan-chitin complex in Saccharomyces cerevisiae. Arch. Microbiol. 1981, 130, 312–318. [Google Scholar] [CrossRef]
- Orlean, P. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef]
- Cabib, E.; Duran, A. Synthase III-dependent chitin is bound to different acceptors depending on location on the cell wall of budding yeast. J. Biol. Chem. 2005, 280, 9170–9179. [Google Scholar] [CrossRef]
- Cabib, E. Two novel techniques for determination of polysaccharide cross-links show that Crh1p and Crh2p attach chitin to both β(1–6)-and β(1–3)glucan in the Saccharomyces cerevisiae cell wall. Eukaryot. Cell 2009, 8, 1626–1636. [Google Scholar] [CrossRef]
- Cabib, E.; Silverman, S.J.; Shaw, J.A. Chitinase and chitin synthase 1: Counterbalancing activities in cell separation of Saccharomyces cerevisiae. Microbiology 1992, 138, 97–102. [Google Scholar] [CrossRef]
- Shaw, J.A.; Mol, P.C.; Bowers, B.; Silverman, S.J.; Valdivieso, M.H.; Durán, A.; Cabib, E. The function of chitin synthases 2 and 3 in the Saccharomyces cerevisiae cell cycle. J. Cell Biol. 1991, 114, 111–123. [Google Scholar] [CrossRef]
- Cabib, E.; Schmidt, M. Chitin synthase III activity, but not the chitin ring, is required for remedial septa formation in budding yeast. FEMS. Microbiol. Lett. 2003, 224, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Bowers, B.; Varma, A.; Roh, D.H.; Cabib, E. In budding yeast, contraction of the actomyosin ring and formation of the primary septum at cytokinesis depend on each other. J. Cell Sci. 2002, 115, 293–302. [Google Scholar] [PubMed]
- Bulawa, C.E. CSD2, CSD3, and CSD4, genes required for chitin synthesis in Saccharomyces cerevisiae: The CSD2 gene product is related to chitin synthases and to developmentally regulated proteins in Rhizobium species and Xenopus laevis. Mol. Cell Biol. 1992, 12, 1764–1776. [Google Scholar] [CrossRef] [PubMed]
- Sanchatjate, S.; Schekman, R. Chs5/6 complex: A multiprotein complex that interacts with and conveys chitin synthase III from the trans-Golgi network to the cell surface. Mol. Biol. Cell 2006, 17, 4157–4166. [Google Scholar] [CrossRef] [PubMed]
- Bernard, M.; Latgé, J.P. Aspergillus fumigatus cell wall: Composition and biosynthesis. Med. Mycol. 2001, 39, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. Bioessays 2006, 28, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Drgonova, J.; Tomás, D.; Tanaka, K.; Kollar, R.; Chen, G.C.; Ford, R.A.; Cabib, E.; Chan, C.S.; Takai, Y. Rho1p, a yeast protein at the interface between cell polarization and morphogenesis. Science 1996, 272, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Tong, A.H.Y.; Lesage, G.; Bader, G.D.; Ding, H.; Xu, H.; Xin, X.; Chen, Y. Global mapping of the yeast genetic interaction network. Science 2004, 303, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Nix, D.E.; Swezey, R.R.; Hector, R.; Galgiani, J.N. Pharmacokinetics of nikkomycin Z after single rising oral doses. Antimicrob. Agents Chemother. 2009, 53, 2517–2521. [Google Scholar] [CrossRef]
- Sucher, A.J.; Chahine, E.B.; Balcer, H.E. Echinocandins: The newest class of antifungals. Ann. Pharmacother. 2009, 43, 1647–1657. [Google Scholar] [CrossRef]
- Cheung, Y.Y.; Hui, M. Effects of echinocandins in combination with nikkomycin Z against invasive Candida albicans bloodstream isolates and the fks mutants. Antimicrob. Agents Chemother. 2017, 61, e00619–e00717. [Google Scholar] [CrossRef] [PubMed]
- Lucero, H.A.; Kuranda, M.J.; Bulik, D.A. A nonradioactive, high throughput assay for chitin synthase activity. Anal. Biochem. 2002, 305, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Shedletzky, E.; Unger, C.; Delmer, D.P. A microtiter-based fluorescence assay for (1,3)-beta-glucan synthases. Anal. Biochem. 1997, 249, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Kingsbury, J.M.; Heitman, J.; Pinnell, S.R. Calcofluor White combination antifungal treatments for Trichophyton rubrum and Candida albicans. PLoS ONE 2017, 7, e39405. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.W.; Martínez-Márquez, J.Y.; Javed, F.T.; Duncan, M.C. A simple and inexpensive quantitative technique for determining chemical sensitivity in Saccharomyces cerevisiae. Sci. Rep. 2018, 8, 11919. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M. Survival and cytokinesis of Saccharomyces cerevisiae in the absence of chitin. Microbiology 2004, 150, 3253–3260. [Google Scholar] [CrossRef] [PubMed]
- Lesage, G.; Shapiro, J.; Specht, C.A.; Sdicu, A.M.; Ménard, P.; Hussein, S.; Bussey, H.; Boone, C. An interactional network of genes involved in chitin synthesis in Saccharomyces cerevisiae. BMC Genet. 2005, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.A.; Munro, C.A.; De Bruijn, I.; Lenardon, M.D.; McKinnon, A.; Gow, N.A. Gow Stimulation of chitin synthesis rescues Candida albicans from echinocandins. PLoS Pathog. 2008, 4, e1000040. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P.; Morin, N.; Baginsky, W.; El-Sherbeini, M.; Clemas, J.A.; Nielsen, J.B.; Foor, F. Differential expression and function of two homologous subunits of yeast 1,3-β-glucan synthase. Mol. Cell. Biol. 1995, 15, 5671–5681. [Google Scholar] [CrossRef]
- Katiyar, S.K.; Alastruey-Izquierdo, A.; Healey, K.R.; Johnson, M.E.; Perlin, D.S.; Edlind, T.D. Fks1 and Fks2 are functionally redundant but differentially regulated in Candida glabrata: Implications for echinocandin resistance. Antimicrob. Agents Chemother. 2012, 56, 6304–6309. [Google Scholar] [CrossRef]
- Baixench, M.T.; Aoun, N.; Desnos-Ollivier, M.; Garcia-Hermoso, D.; Bretagne, S.; Ramires, S.; Piketty, C.; Dannaoui, E. Acquired resistance to echinocandins in Candida albicans: Case report and review. J. Antimicrob. Chemother. 2007, 59, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Niimi, K.; Monk, B.C.; Hirai, A.; Hatakenaka, K.; Umeyama, T.; Lamping, E.; Uehara, Y.; Maki, K.; Tanabe, K.; Kamimura, T.; et al. Clinically significant micafungin resistance in Candida albicans involves modification of a glucan synthase catalytic subunit GSC1 (FKS1) allele followed by loss of heterozygosity. J. Antimicrob. Chemother. 2010, 65, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Soulié, M.C.; Piffeteau, A.; Choquer, M.; Boccara, M.; Vidal-Cros, A. Disruption of Botrytis cinerea calss I chitin synthase gene Bcchs 1 results in cell wall weakening and reduced virulence. Fungal Genet. Biol. 2003, 40, 38–46. [Google Scholar] [CrossRef]
- Faurobert, M.; Pelpoir, E.; Chaïb, J. Phenol extraction of proteins for proteomic studies of recalcitrant plant tissues. Methods Mol. Biol. 2007, 355, 9–14. [Google Scholar] [PubMed]
- Elias, J.E.; Gygi, S.P. Target-decoy search strategy for increased confidence in large-scale protein identifications by mass spectrometry. Nat. Methods 2008, 4, 207–214. [Google Scholar] [CrossRef]
- Gan, C.S.; Chong, P.K.; Pham, T.K.; Wright, P.C. Technical, experimental, and biological variations in isobaric tags for relative and absolute quantitation (iTRAQ). J. Proteome Res. 2007, 6, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Unwin, R.D.; Griffiths, J.R.; Whetton, A.D. Simultaneous analysis of relative protein expression levels across multiple samples using iTRAQ isobaric tags with 2D nano LC-MS/MS. Nat. Protoc. 2010, 5, 1574–1582. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Bolden, C.B.; Iqbal, N.; Kuykendall, R.J. Validation of 24-hour flucytosine MIC determination by comparison with 48-hour determination by the Clinical and Laboratory Standards Institute M27-A3 broth microdilution reference method. J. Clin. Microbiol. 2011, 49, 4322–4325. [Google Scholar] [CrossRef]
- Kuykendall, R.J.; Lockhart, S.R. Microbroth dilution susceptibility testing of Candida species. Methods Mol. Biol. 2016, 1356, 173–181. [Google Scholar]
Sample Availability: Samples of the compounds IMB-D10 and IMB-F4 are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | chs3∆ | fks1∆ | |
|---|---|---|---|
| Nikkomycin IMB-D10 | >200 50 | >200 100 | 25 25 |
| IMB-F4 | >100 | >100 | 50 |
| Compound | MIC (μg/mL) | MIC of Caspofungin (μg/mL) | FICI | |
|---|---|---|---|---|
| Alone | Combination | Combination | ||
| Nikkomycin | >100 | 12.5 | 0.0075 | <0.5 |
| IMB-D10 | 50 | 12.5 | 0.0075 | 0.5 |
| IMB-F4 | >100 | 25 | 0.0075 | <0.5 |
| Caspofungin | 0.03 | - | - | - |
| Compound | MIC (μg/mL) | MIC of Caspofungin(μg/mL) | FICI | |
|---|---|---|---|---|
| Alone | Combination | Combination | ||
| Nikkomycin | 12.5 | 3.125 | 0.00625 | <0.5 |
| IMB-D10 | 100 | 12.5 | 0.00625 | 0.5 |
| IMB-F4 | >100 | 12.5 | 0.003125 | <0.5 |
| Caspofungin | 0.025 | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Sun, H.; Zhu, X.; Bian, C.; Wang, Y.; Si, S. Identification of New Antifungal Agents Targeting Chitin Synthesis by a Chemical-Genetic Method. Molecules 2019, 24, 3155. https://doi.org/10.3390/molecules24173155
Li Y, Sun H, Zhu X, Bian C, Wang Y, Si S. Identification of New Antifungal Agents Targeting Chitin Synthesis by a Chemical-Genetic Method. Molecules. 2019; 24(17):3155. https://doi.org/10.3390/molecules24173155
Chicago/Turabian StyleLi, Yan, Hongmin Sun, Xiaohong Zhu, Cong Bian, Yanchang Wang, and Shuyi Si. 2019. "Identification of New Antifungal Agents Targeting Chitin Synthesis by a Chemical-Genetic Method" Molecules 24, no. 17: 3155. https://doi.org/10.3390/molecules24173155
APA StyleLi, Y., Sun, H., Zhu, X., Bian, C., Wang, Y., & Si, S. (2019). Identification of New Antifungal Agents Targeting Chitin Synthesis by a Chemical-Genetic Method. Molecules, 24(17), 3155. https://doi.org/10.3390/molecules24173155