Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation

Abstract
:1. Introduction
2. Results
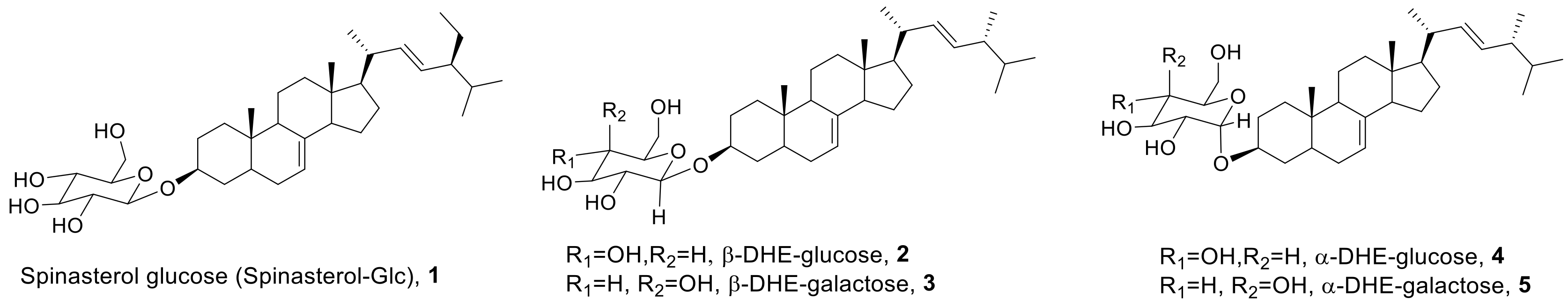
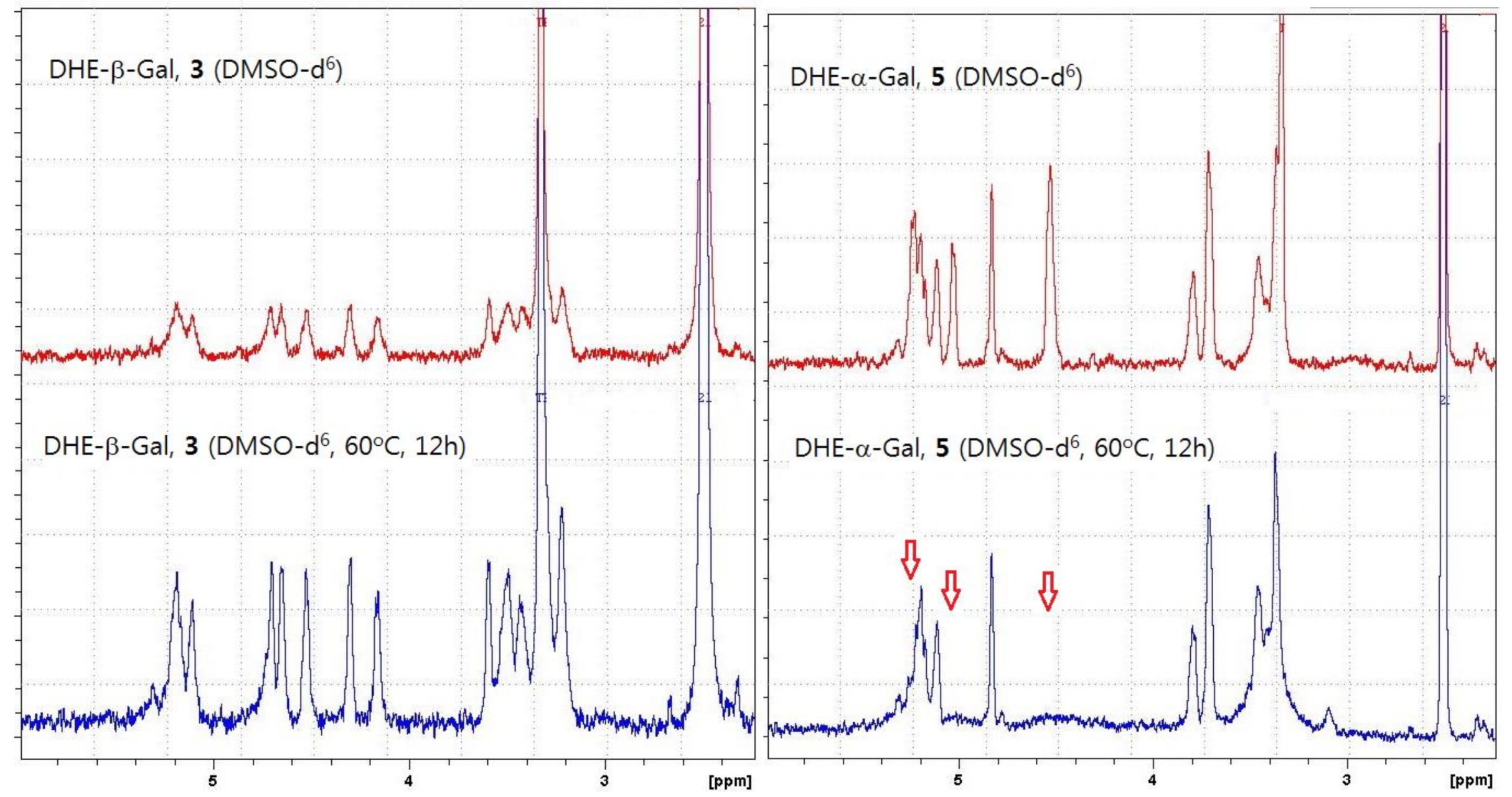
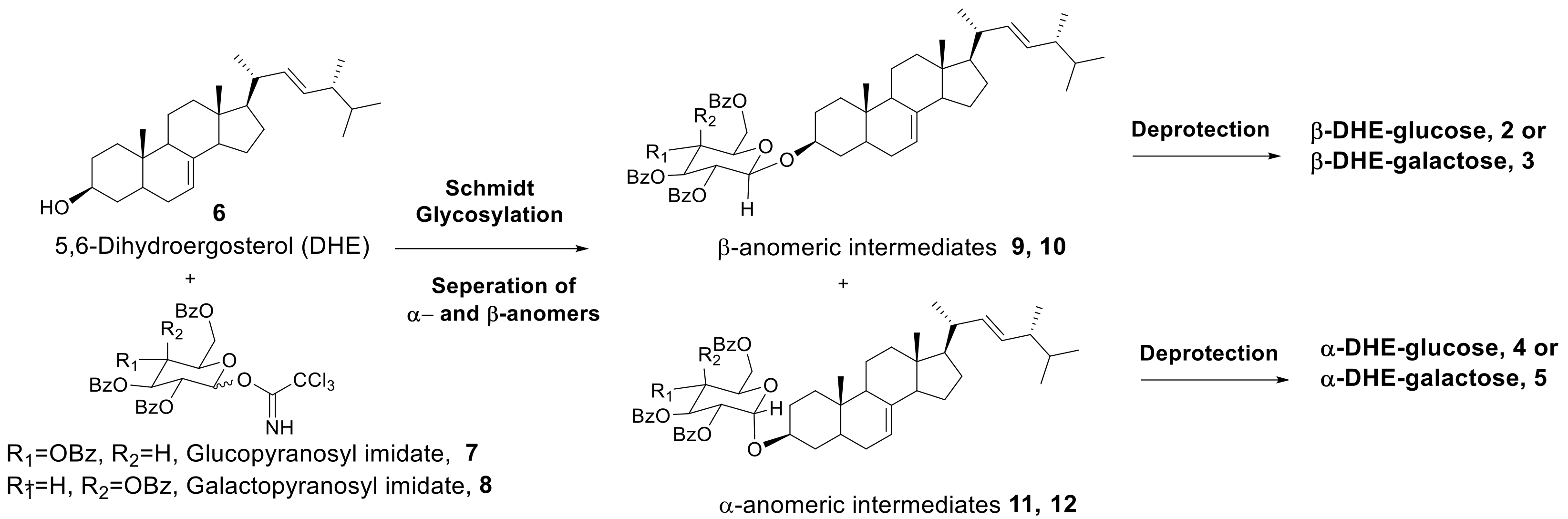
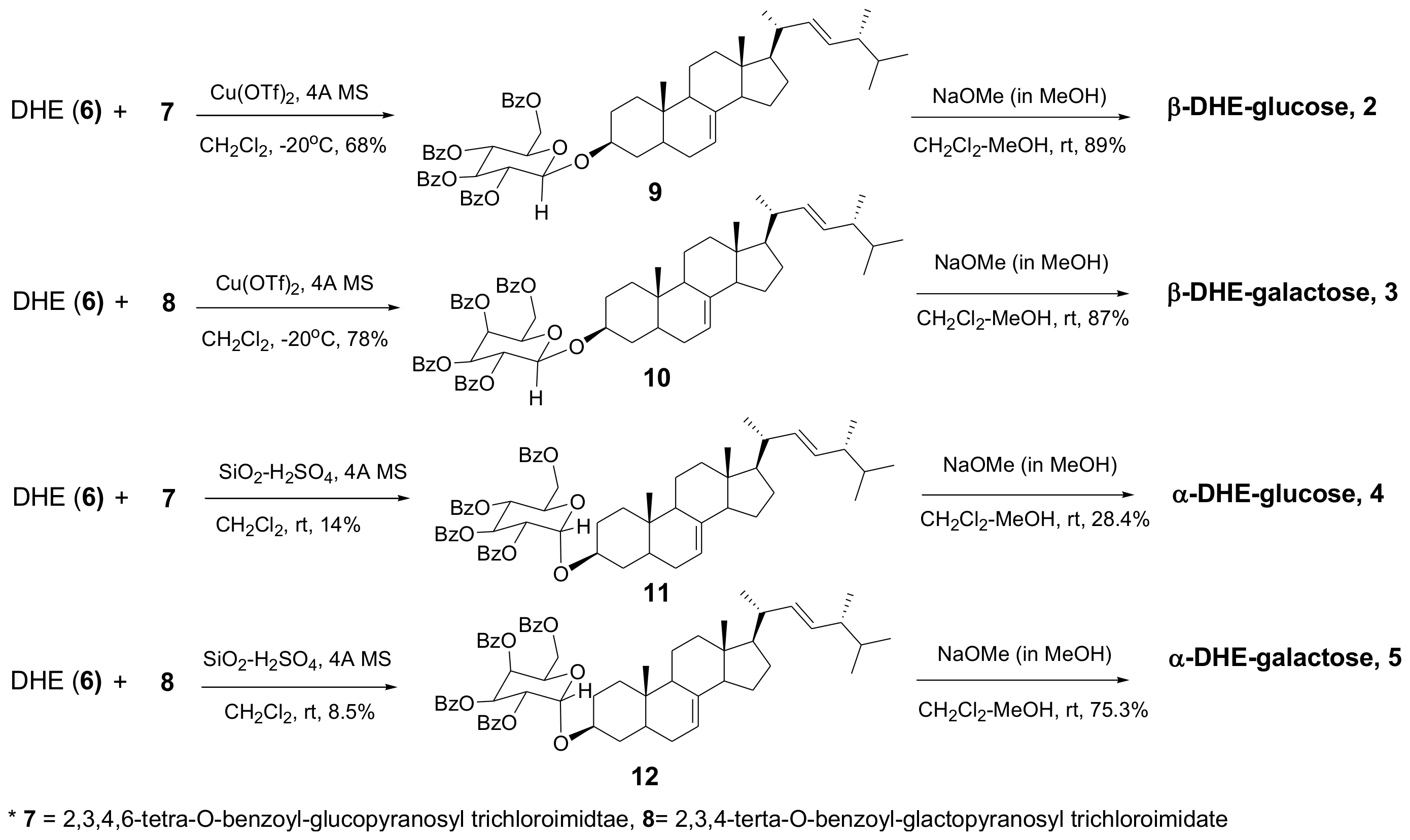
2.1. Synthesis of α- and β-Anomers of 5,6-Dihydroergosterol-Glycosides (DHE-Glycosides)
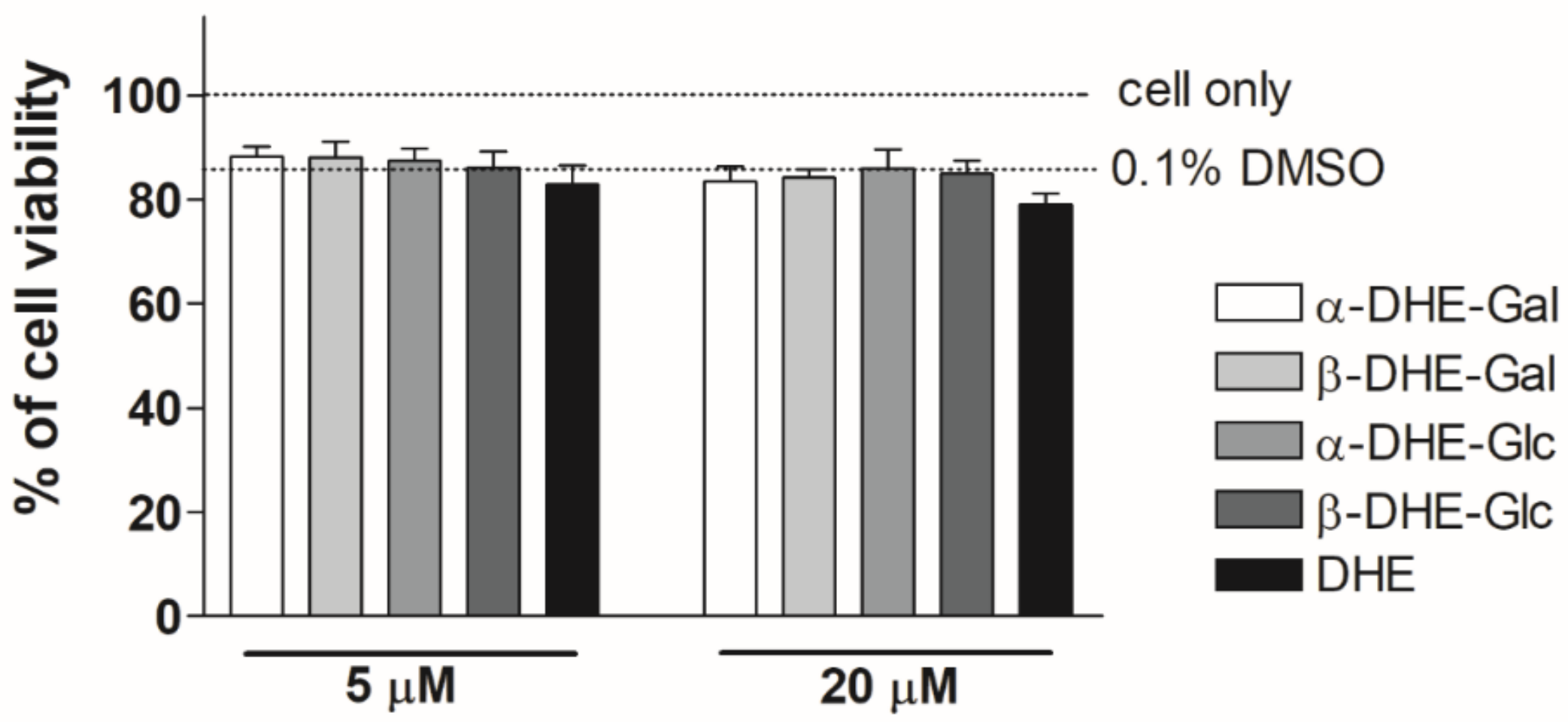
2.2. Effect of α- and β-Anomers of 5,6-Dihydroergosterol(DHE)-Glycosides on Cell Viability
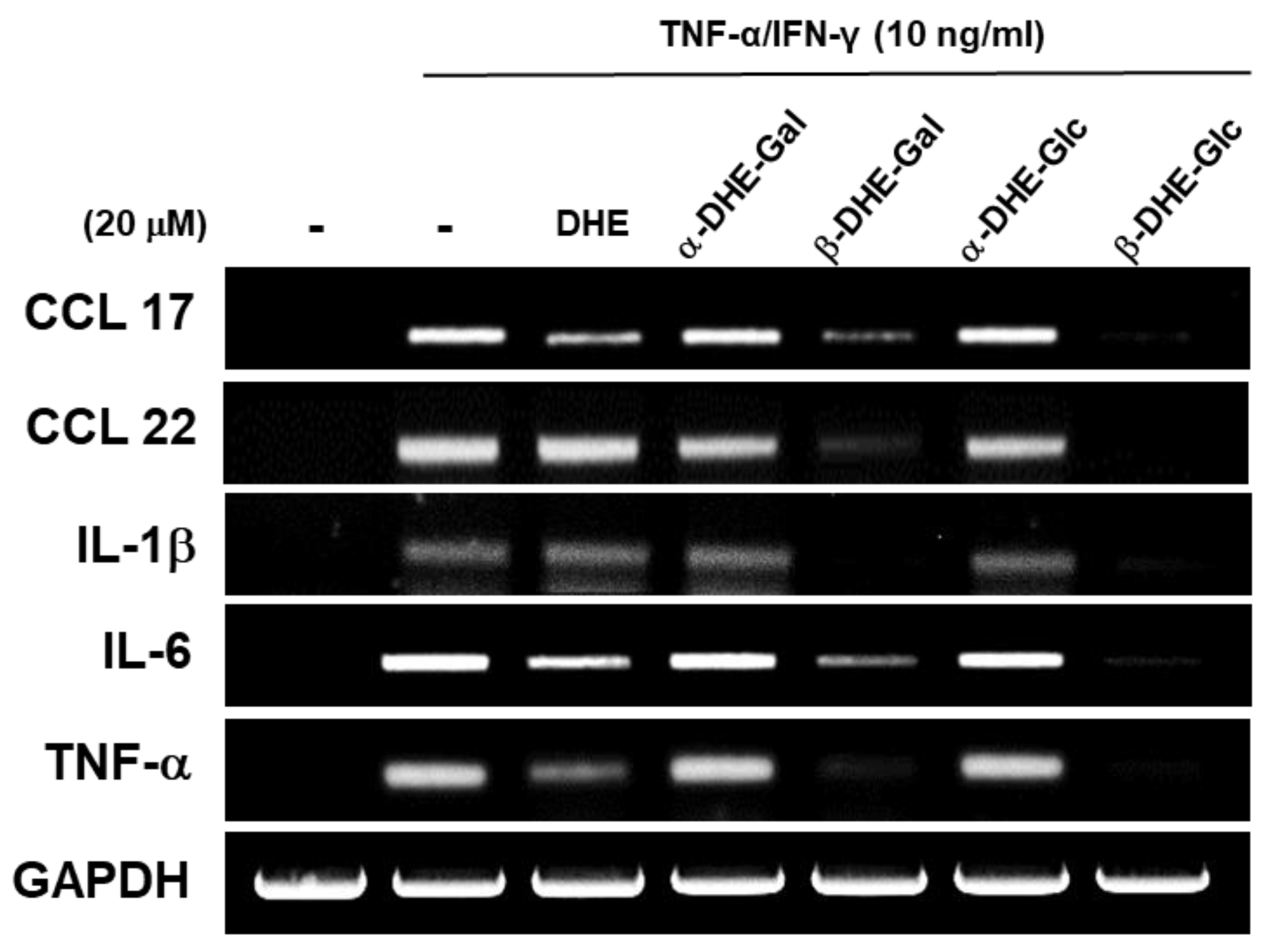
2.3. Effects of DHE-Glycoside α- and β-Anomers on mRNA Expression Levels of Pro-Inflammatory Cytokines and Chemokines in TNF-α/IFN-γ Induced HaCaT Cells
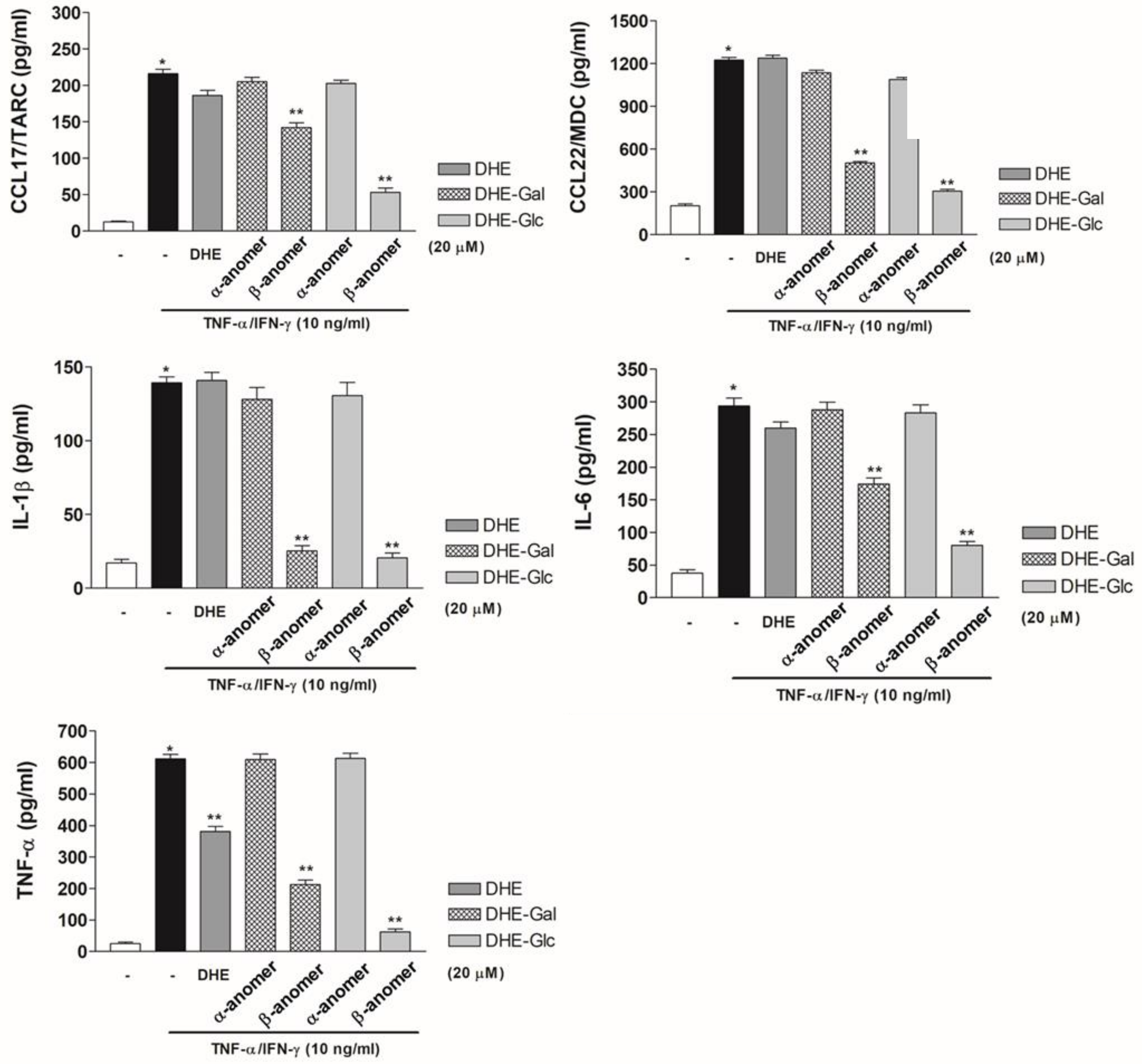
2.4. Effects of DHE-Glycoside α- and β-Anomers on Pro-Inflammatory Cytokine and Chemokine Protein Expression Levels in TNF-α/IFN-γ-Induced HaCaT Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Instruments
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) Analysis
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Fiore, C.; Eisenhut, M.; Ragazzi, E.; Zanchin, G.; Armanini, D. A history of the therapeutic use of liquorice in Europe. J. Ethnopharmacol. 2005, 99, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Sparg, S.G.; Light, M.E.; van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.R.; Lee, T.H.; Bang, M.H.; Kim, H.; Son, Y.; Chung, D.K.; Kim, J. Suppression of thymus- and activation-regulated chemokine (TARC/CCL17) production by 3-O-β-d-glucopyanosylspinasterol via blocking NF-κB and STAT1 signaling pathways in TNF-α and IFN-γ-induced HaCaT keratinocytes. Biochem. Biophys. Res. Commun. 2012, 427, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Giroux, S.; Corey, E.J. Stereocontrolled synthesis of dafachronic acid A, the ligand for the DAF-12 nuclear receptor of Caenorhabditis elegans. J. Am. Chem. Soc. 2007, 129, 9866–9867. [Google Scholar] [CrossRef] [PubMed]
- Badreddine, A.; Mostafa, K.E.; Zarrouk, A.; Nury, T.; Kharassi, Y.E.; Nasser, B.; Malki, M.C.; Lizard, G.; Samadi, M. An expeditious synthesis of spinasterol and schottenol, two phytosterols present in argan oil and in cactus pear seed oil, and evaluation of their biological activities on cells of the central nervous system. Steroids 2015, 99, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Park, H.G.; Lee, T.H.; Chang, F.; Kwon, H.J.; Kim, J.Y.; Kim, H.W. Synthesis of ergosterol and 5,6-dihydroergosterol glycosides and their inhibitory activities on lipopolysaccharide-induced nitric oxide production. Bull. Korean Chem. Soc. 2013, 34, 1339–1344. [Google Scholar] [CrossRef]
- Jung, M.; Lee, T.H.; Oh, H.J.; Kim, H.; Son, Y.; Lee, E.H.; Kim, J. Inhibitory effect of 5,6-dihydroergosteol-glucoside on atopic dermatitis-like skin lesions via suppression of NF-κB and STAT activation. J. Dermatol. Sci. 2015, 79, 252–261. [Google Scholar] [CrossRef]
- Kasraie, S.; Werfel, T. Role of macrophages in the pathogenesis of atopic dermatitis. Mediators Inflamm. 2013, 2013. [Google Scholar] [CrossRef]
- Yamanaka, K.; Mizutani, H. The role of cytokines/chemokines in the pathogenesis of atopic dermatitis. Curr. Probl. Dermatol. 2011, 41, 80–92. [Google Scholar]
- Caproni, M.; Torchia, D.; Cardinali, C.; Volpi, W.; Del Bianco, E.; D’Agata, A.; Fabbri, P. Infiltrating cells, related cytokines and chemokine receptors in lesional skin of patients with dermatomyositis. Br. J. Dermatol. 2004, 151, 784–791. [Google Scholar] [CrossRef]
- Mu, Z.; Zhao, Y.; Chang, C.; Zhang, J. Molecular biology of atopic dermatitis. Clin. Rev. Allergy Immunol. 2014, 47, 193–218. [Google Scholar] [CrossRef] [PubMed]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Albanesi, C.; Scarponi, C.; Sebastiani, S.; Cavani, A.; Federici, M.; Sozzani, S.; Girolomoni, G. A cytokine-to-chemokine axis between T lymphocytes and keratinocytes can favor Th1 cell accumulation in chronic inflammatory skin diseases. J. Leukoc. Biol. 2001, 70, 617–623. [Google Scholar] [PubMed]
- Scheu, S.; Ali, S.; Ruland, C.; Arolt, V.; Alferink, J. The C-C Chemokines CCL17 and CCL22 and Their Receptor CCR4 in CNS Autoimmunity. Int. J. Mol. Sci. 2017, 18, 2306. [Google Scholar] [CrossRef] [PubMed]
- Osabe, M.; Tajika, T.; Tohkin, M. Allopurinol suppresses expression of the regulatory T-cell migration factors TARC/CCL17 and MDC/CCL22 in HaCaT keratinocytes via restriction of nuclear factor-κB activation. J. Appl. Toxicol. 2018, 38, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Miyano, K.; Matsushita, S.; Tsuchida, T.; Nakamura, K. Inhibitory effect of a histamine 4 receptor antagonist on CCL17 and CCL22 production by monocyte-derived Langerhans cells in patients with atopic dermatitis. J. Dermatol. 2016, 43, 1024–1029. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.S.; Oh, S.H.; Wu, W.H.; Bae, B.G.; Lee, H.J.; Lee, M.G.; Lee, K.H. CC chemokines as potential immunologic markers correlated with clinical improvement of atopic dermatitis patients by immunotherapy. Exp. Dermatol. 2010, 19, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Narbutt, J.; Lesiak, A.; Sysa-Jedrzeiowska, A.; Zakrzewski, M.; Bogaczewicz, J.; Stelmach, I.; Kuna, P. The imbalance in serum concentration of Th-1- and Th-2-derived chemokines as one of the factors involved in pathogenesis of atopic dermatitis. Mediators Inflamm. 2009, 2009. [Google Scholar] [CrossRef] [PubMed]
- Wuts, P.G.M.; Green, T.W. Protective Group in Organic Synthesis, 4th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 102–120. [Google Scholar]
- Lee, T.H.; Kwak, H.B.; Kim, H.H.; Lee, Z.H.; Chung, D.K.; Baek, N.I.; Kim, J. Methanol extracts of Stewartia koreana inhibit cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) gene expression by blocking NF-kappaB transactivation in LPS-activated RAW264.7 cells. Mol. Cells 2007, 23, 398–404. [Google Scholar]
- Lee, T.H.; Jung, M.; Bang, M.H.; Chung, D.K.; Kim, J. Inhibitory effects of a spinasterol glycoside on lipopolysaccharide-induced production of nitric oxide and proinflammatory cytokines via down-regulating MAP kinase pathways and NF-kB activation in RAW264.7 macrophage cells. Int. Immunopharmacol. 2012, 13, 264–270. [Google Scholar] [CrossRef]
- Lee, K.S.; Chun, S.Y.; Lee, M.G.; Kim, S.; Jang, T.J.; Nam, K.S. The prevention of TNF-α/IFN-γ mixture-induced inflammation in human keratinocyte and atopic dermatitis-like skin lesions in Nc/Nga mice by mineral-balanced deep sea water. Biomed. Pharmacother. 2018, 97, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Mazanet, M.M.; Neote, K.; Hughes, C.C. Expression of IFN-inducible T cell alpha chemoattractant by human endothelial cells is cyclosporin A-resistant and promotes T cell adhesion: Implications for cyclosporin A-resistant immune inflammation. J. Immunol. 2000, 164, 5383–5388. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Sugaya, M.; Suga, H.; Oka, T.; Kawaguchi, M.; Miyagaki, T.; Fujita, H.; Sato, S. Thymic Stromal Chemokine TSLP Acts through Th2 Cytokine Production to Induce Cutaneous T-cell Lymphoma. Cancer Res. 2016, 76, 6241–6252. [Google Scholar] [CrossRef] [PubMed]
- Mollanazar, N.K.; Smith, P.K.; Yosipovitch, G. Mediators of Chronic Pruritus in Atopic Dermatitis: Getting the Itch Out? Clin. Rev. Allergy Immunol. 2016, 51, 263–292. [Google Scholar] [CrossRef] [PubMed]
- Rebane, A.; Zimmermann, M.; Aab, A.; Baurecht, H.; Koreck, A.; Karelson, M.; Abram, K.; Metsalu, T.; Pihlap, M.; Meyer, N.; et al. Mechanisms of IFN-γ-induced apoptosis of human skin keratinocytes in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2012, 129, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Byrne, A.M.; Goleva, E.; Chouiali, F.; Kaplan, M.H.; Hamid, Q.A.; Leung, D.Y. Induction of GITRL expression in human keratinocytes by Th2 cytokines and TNF-α: Implications for atopic dermatitis. Clin. Exp. Allergy 2012, 42, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Giustizieri, M.L.; Mascia, F.; Frezzolini, A.; De Pità, O.; Chinni, L.M.; Giannetti, A.; Girolomoni, G.; Pastore, S. Keratinocytes from patients with atopic dermatitis and psoriasis show a distinct chemokine production profile in response to T cell-derived cytokines. J. Allergy Clin. Immunol. 2001, 107, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, M.S.; Jeong, G.S.; Yoon, J. Xanthii fructus extract inhibits TNF-α/IFN-γ-induced Th2-chemokines production via blockade of NF-κB, STAT1 and p38-MAPK activation in human epidermal keratinocytes. J. Ethnopharmacol. 2015, 171, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Classen, A.; Kalali, B.N.; Schnopp, C.; Andres, C.; Aguilar-Pimentel, J.A.; Ring, J.; Ollert, M.; Mempel, M. TNF receptor I on human keratinocytes is a binding partner for staphylococcal protein A resulting in the activation of NF kappa B, AP-1, and downstream gene transcription. Exp. Dermatol. 2011, 20, 48–52. [Google Scholar] [CrossRef]
- Usabiaga, I.; González, J.; León, I.; Arnaiz, P.F.; Cocinero, E.J.; Ferńandez, J.A. Influence of the anomeric conformation in the intermolecular interactions of glucose. J. Phys. Chem. Lett. 2017, 8, 1147–1151. [Google Scholar] [CrossRef]
- Davis, A.P.; Wareham, R.S. Carbohydrate Recognition through Noncovalent Interactions: A Challenge for Biomimetic and Supramolecular Chemistry. Angew. Chem. Int. Ed. 1999, 38, 2978–2996. [Google Scholar] [CrossRef]
- Mbadugha, B.N.S.; Menger, F.M. Sugar/Steroid/Sugar conjugates: Sensitivity of lipid binding to sugar structure. Org. Lett. 2003, 5, 4041–4044. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Y.; Yao, Q.; Liu, Y.; Si, C.L.; Chen, J.; Jiu, N.; Lian, H.Y.; Ding, L.N.; Zhang, Y.M. Highly efficient synthesis and antitumor activity of monosaccharide saponins mimicking components of Chinese folk medicine Cordyceps sinensis. J. Asian Nat. Pro. Res. 2012, 14, 429–435. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the α-and β-anomers of DHE-glycosides are available from the authors upon request. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Entry | Acid Catalyst a | Temp. | Time (h) | Yield (%) b | |
|---|---|---|---|---|---|
| α-anomer, 11 | β-anomer, 9 | ||||
| 1 | Cellulose-HClO4 | R.T. | 48 | Trace | 58% |
| 2 | SiO2-H2SO4 | R.T. | 24 | 10% | 55% |
| 3 | TMSOTf | −20 °C | 24 | Trace | 77% |
| 4 | Cu(OTf)2 | −20 °C | 24 | Trace | 68% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.K.; Cho, Y.K.; Park, H.; Lee, T.H.; Kim, H. Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation. Molecules 2019, 24, 371. https://doi.org/10.3390/molecules24020371
Kim TK, Cho YK, Park H, Lee TH, Kim H. Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation. Molecules. 2019; 24(2):371. https://doi.org/10.3390/molecules24020371
Chicago/Turabian StyleKim, Tae Kyun, Young Kyoung Cho, HoonGyu Park, Tae Hoon Lee, and Hakwon Kim. 2019. "Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation" Molecules 24, no. 2: 371. https://doi.org/10.3390/molecules24020371
APA StyleKim, T. K., Cho, Y. K., Park, H., Lee, T. H., & Kim, H. (2019). Comparison of the Inhibitory Activities of 5,6-Dihydroergosterol Glycoside α- and β-Anomers on Skin Inflammation. Molecules, 24(2), 371. https://doi.org/10.3390/molecules24020371