Hybridization of Particulate Methane Monooxygenase by Methanobactin-Modified AuNPs

Abstract
:1. Introduction
2. Results and Discussions
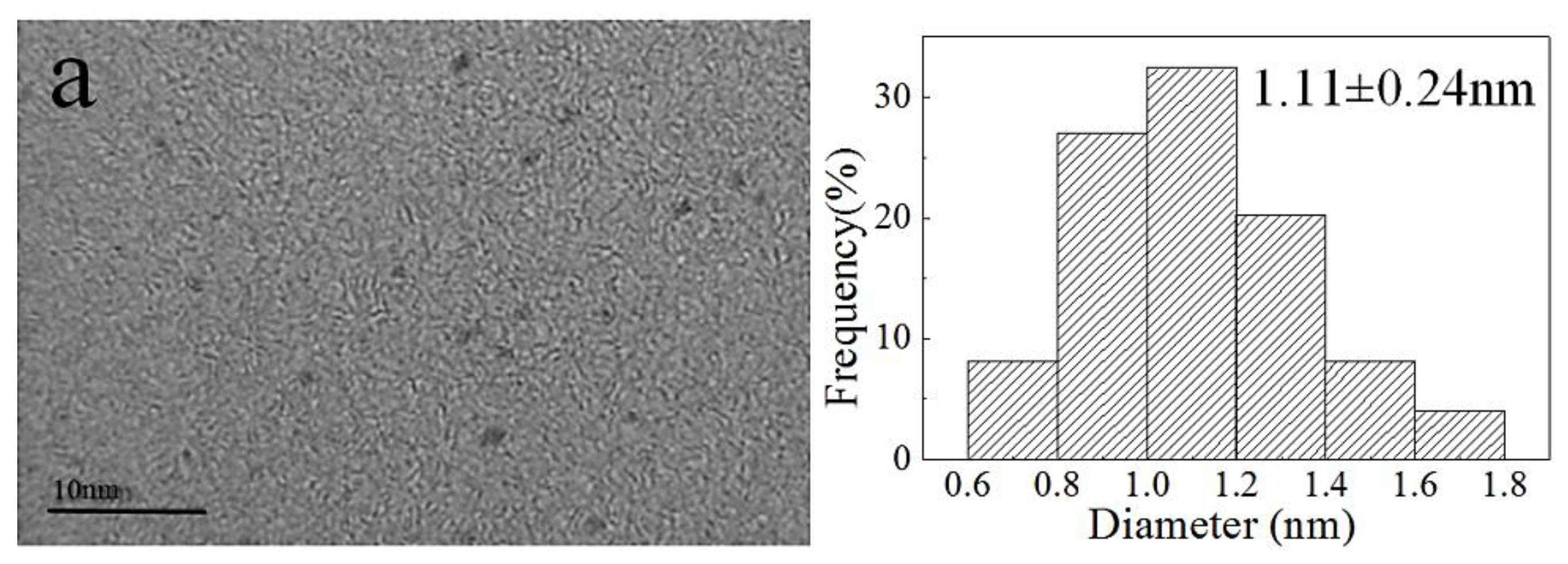
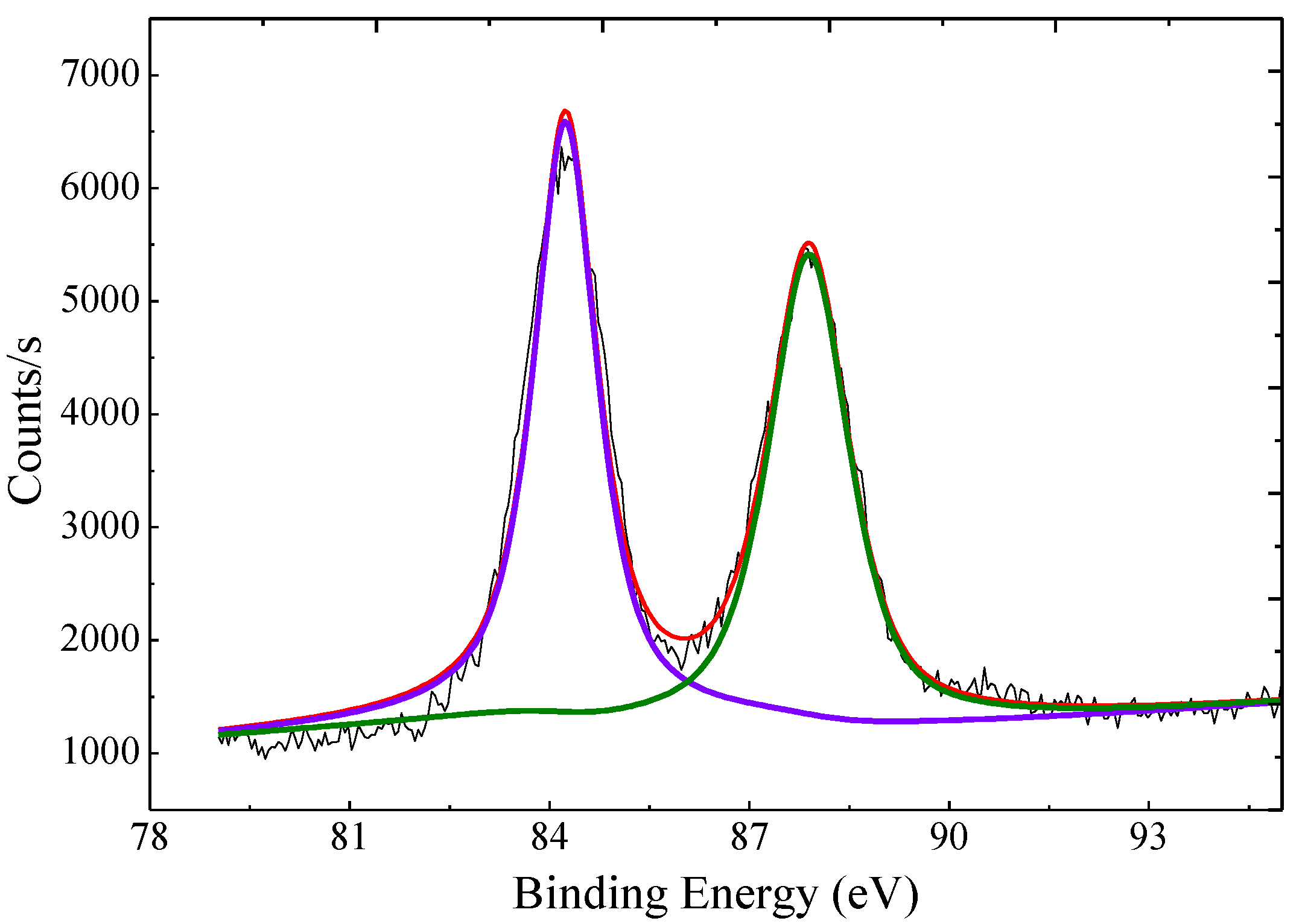
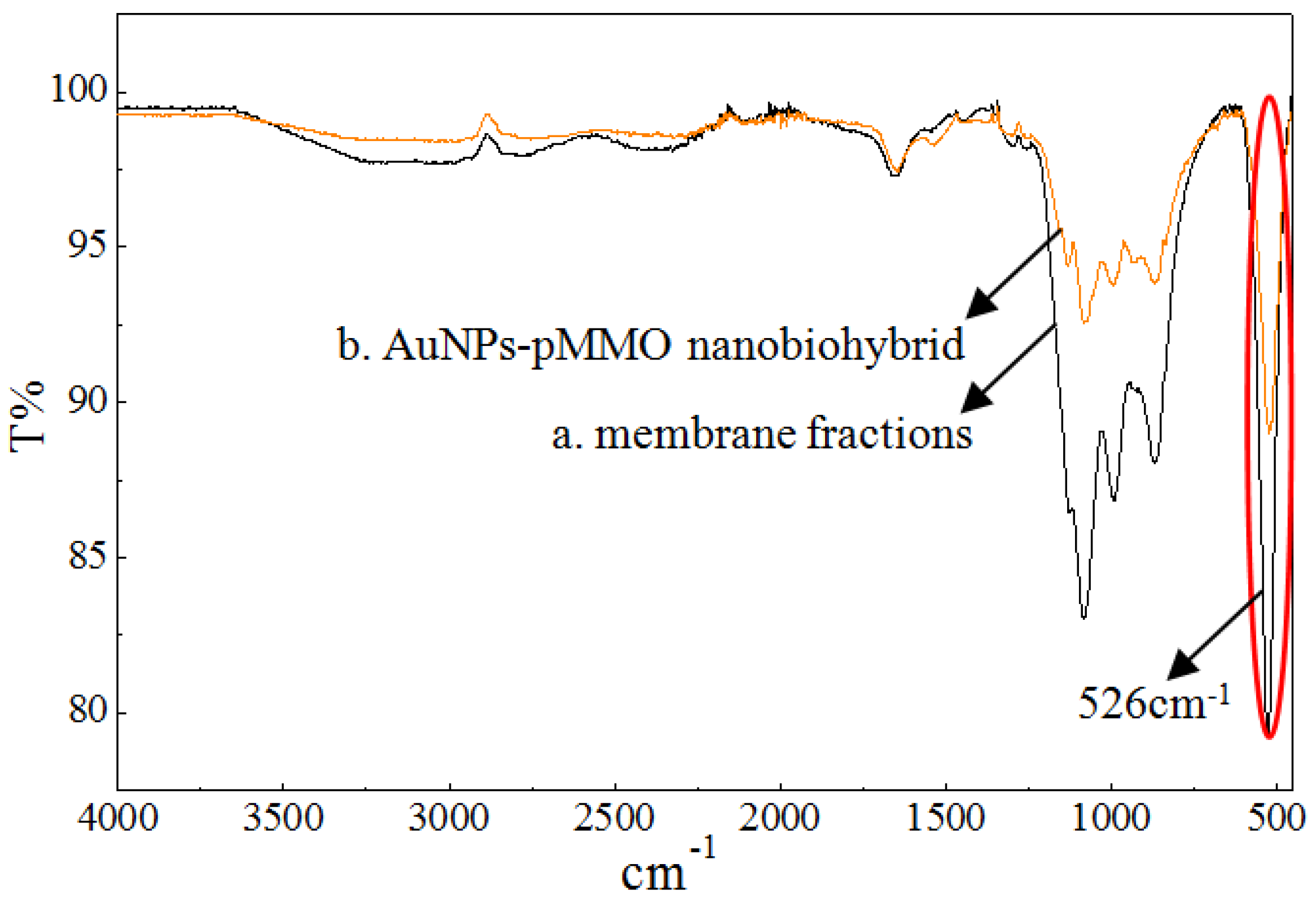
2.1. Preparation and Characterization of AuNP–pMMO Nanobiohybrids
2.2. MMO Activity of AuNP–pMMO Nanobiohybrids
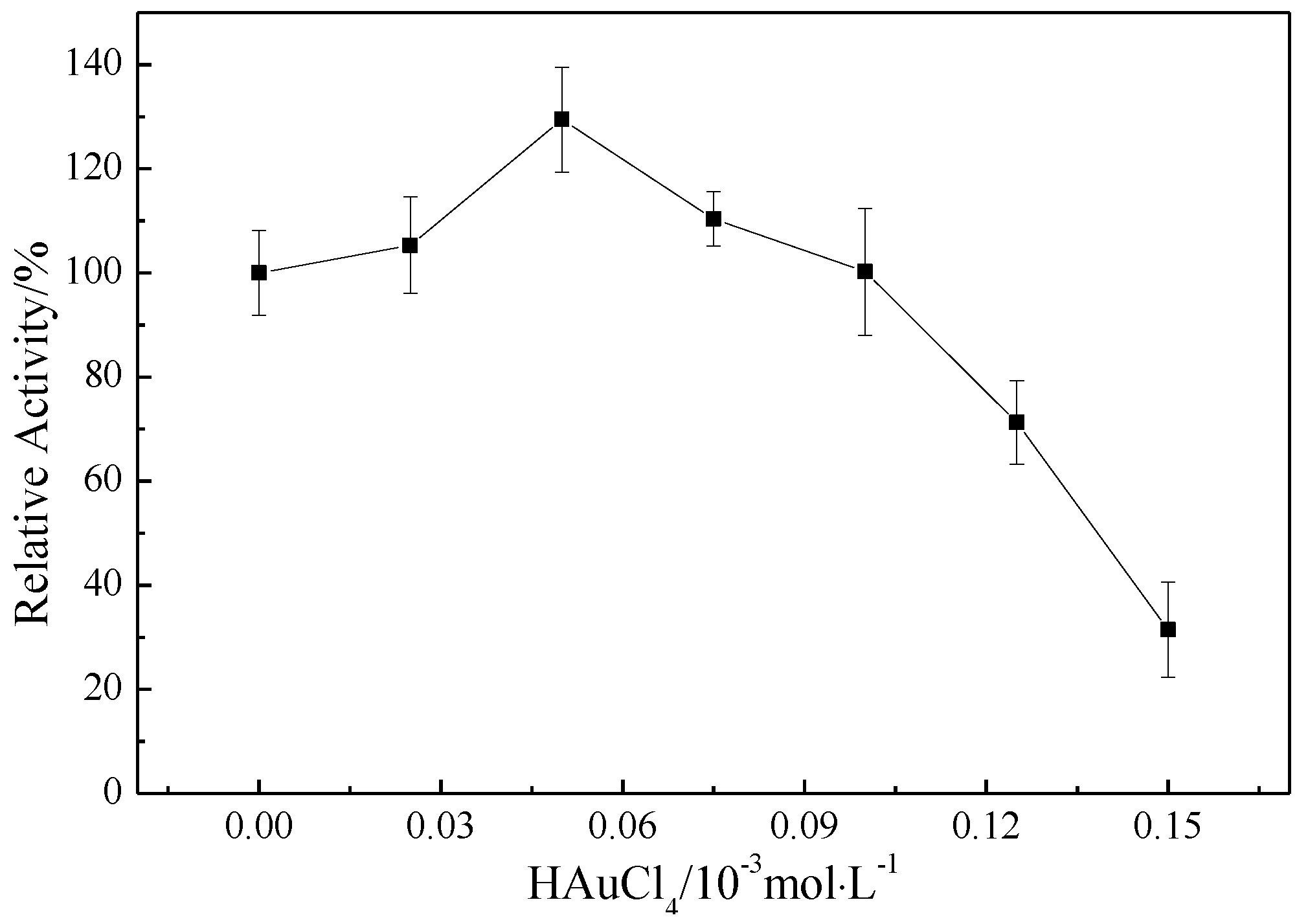
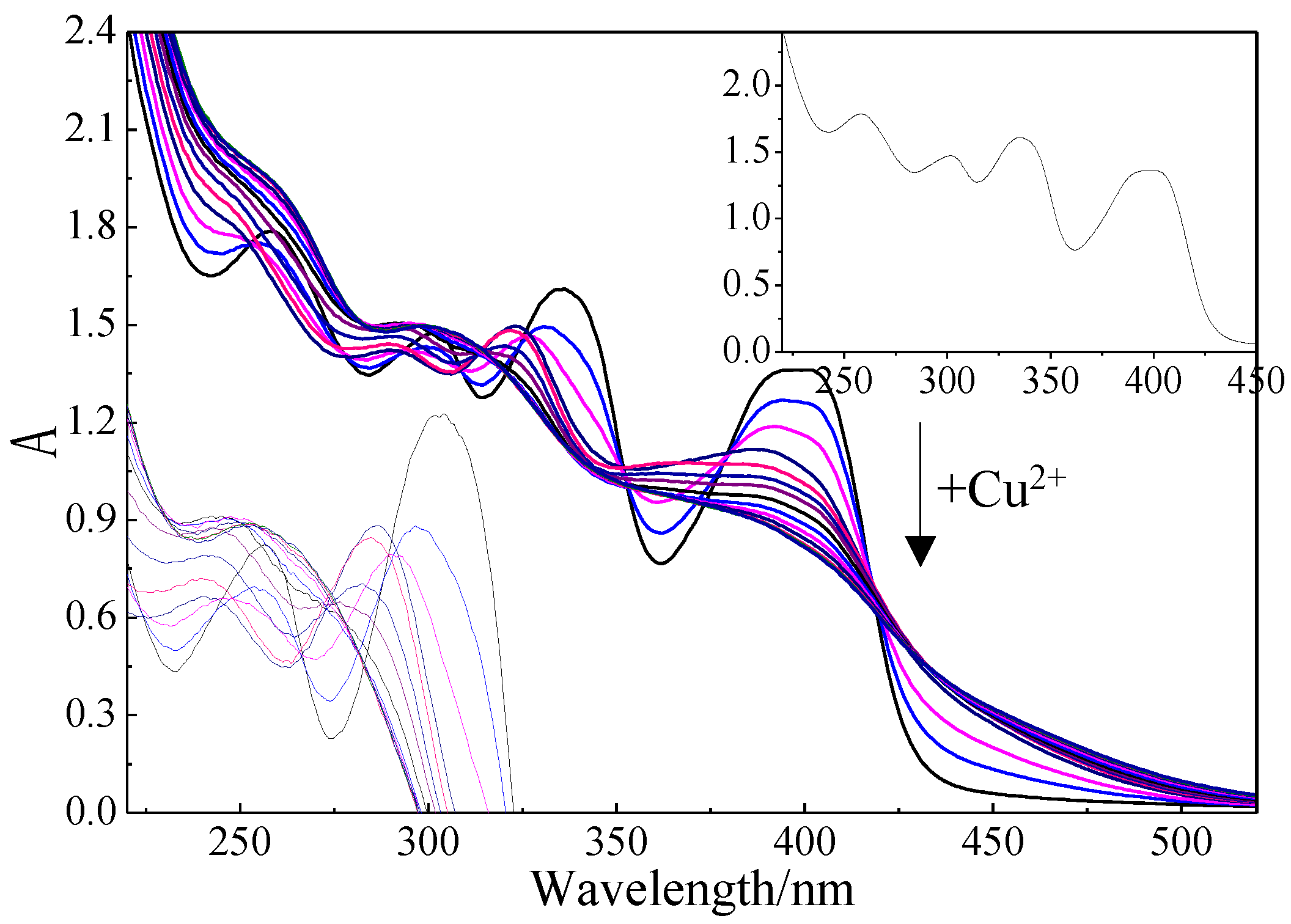
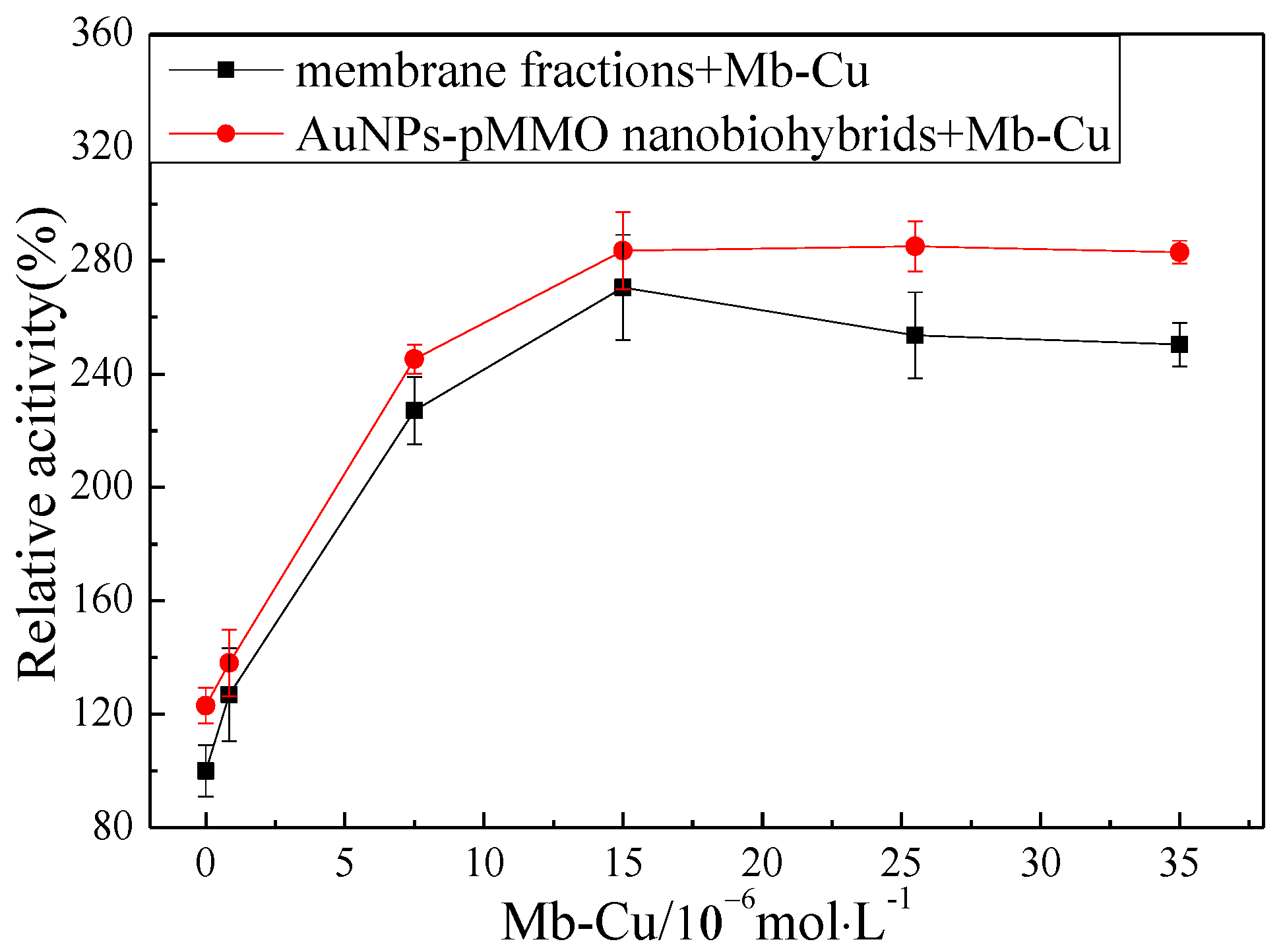
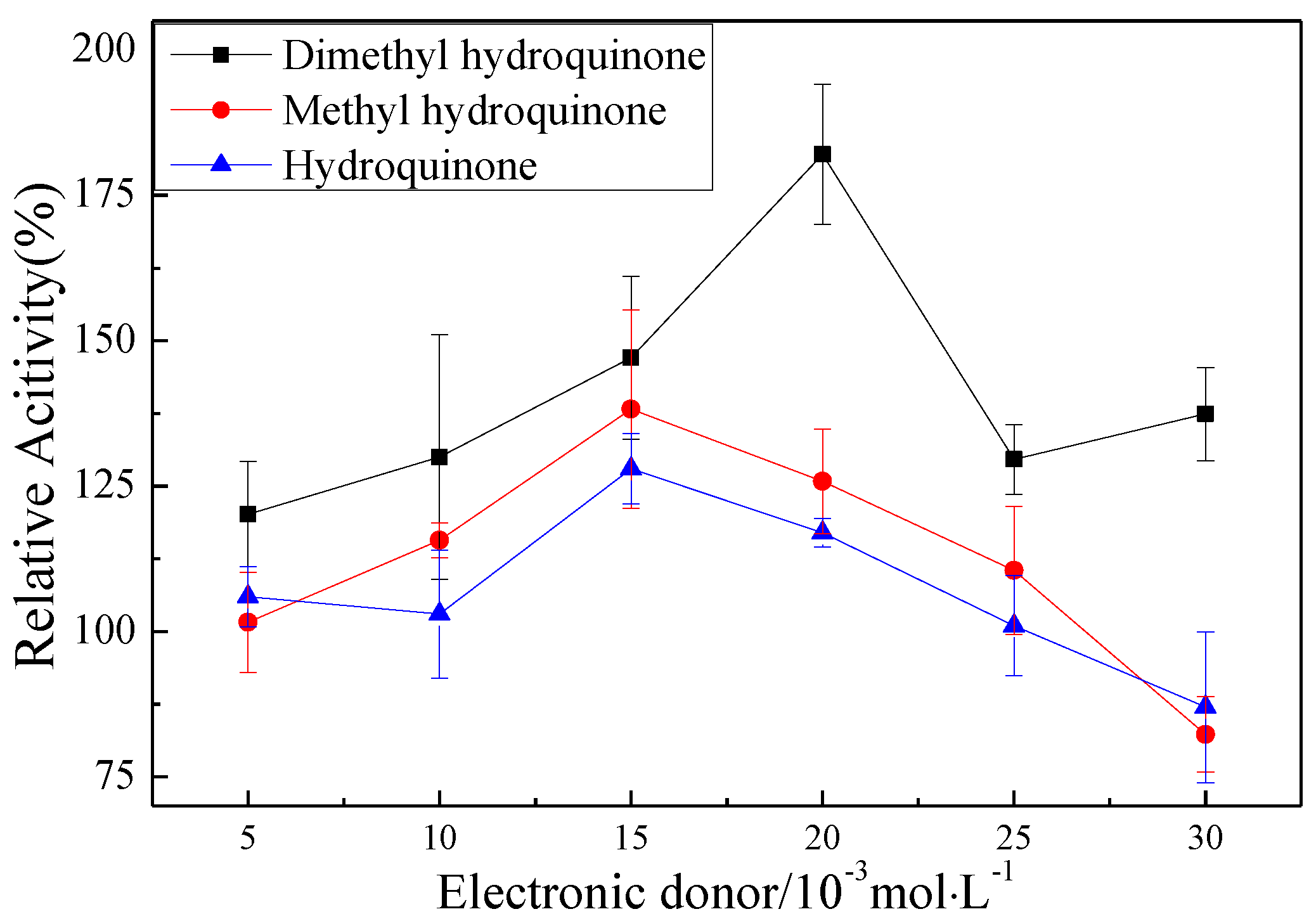
2.3. Effect of Mb–Cu-Modified AuNP–pMMO Nanobiohybrids on the Catalytic Activity of pMMO
2.4. Reuse of Mb–Cu-Modified AuNP–pMMO Nanobiohybrids
3. Materials and Methods
3.1. Chemicals and Apparatus
3.2. Microorganism and Culture Conditions
3.3. Isolation and Quantification of Mb and Mb–Cu
3.4. Preparation of pMMO-Enriched Membrane Fraction
3.5. Activity Assay of pMMO
3.6. Assay of Peroxidase-Like Activity
3.7. Synthesis of AuNP–pMMO Nanobiohybrids
3.8. Characterization of AuNP–pMMO Nanobiohybrids
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sirajuddin, S.; Rosenzweig, A.C. Enzymatic oxidation of methane. Biochemistry 2015, 54, 2283–2294. [Google Scholar] [CrossRef] [PubMed]
- Semrau, J.D.; Dispirito, A.A.; Yoon, S. Methanotrophs and copper. FEMS Microbiol. Rev. 2010, 34, 496–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Mccormick, M.S.; Lippard, S.J.; Cho, U.S. Control of substrate access to the active site in methane monooxygenase. Nature 2013, 494, 380–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakemian, A.S.; Rosenzweig, A.C. The biochemistry of methane oxidation. Annu. Rev. Biochem. 2007, 76, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.S.-F.; Chen, K.H.-C.; Tseng, M.Y.-H.; Wang, Y.-S.; Tseng, C.-F.; Chen, Y.-J.; Huang, D.-S.; Chan, S.I. Production of high-quality particulate methane monooxygenase in high yields from Methylococcus capsulatus (Bath) with a hollow-fiber membrane bioreactor. J. Bacteriol. 2003, 185, 5915–5924. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.C.-C.; Maji, S.; Chen, P.P.-Y.; Lee, H.K.; Yu, S.S.-F.; Chan, S.I. Alkane oxidation: Methane monooxygenases, related enzymes, and their biomimetics. Chem. Rev. 2017, 117, 8574–8621. [Google Scholar] [CrossRef]
- Choi, D. Methanobactin and the Membrane-Associated Methane Monooxygenase in Methanotrophy: A Tale of Two Proteins. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2007. [Google Scholar]
- Ul-haque, M.F.; Kalidass, B.; Vorobev, A.; Baral, B.S.; Dispirito, A.A.; Semrau, J.D. Methanobactin from Methylocystis sp. strain SB2 affects gene expression and methane monooxygenase activity in methylosinus trichosporium OB3b. Appl. Environ. Microbiol. 2015, 81, 2466–2473. [Google Scholar] [CrossRef]
- Miyaji, A.; Suzuki, M.; Baba, T.; Kamachi, T.; Okura, I. Hydrogen peroxide as an effecter on the inactivation of particulate methane monooxygenase under aerobic conditions. J. Mol. Catal. B Enzym. 2009, 57, 211–215. [Google Scholar] [CrossRef]
- Choi, D.W.; Semrau, J.D.; Antholine, W.E.; Hartsel, S.C.; Anderson, R.C.; Carey, J.N.; Dreis, A.M.; Kenseth, E.M.; Renstrom, J.M.; Scardino, L.L. Oxidase, superoxide dismutase, and hydrogen peroxide reductase activities of methanobactin from types I and II methanotrophs. J. Inorg. Biochem. 2008, 102, 1571–1580. [Google Scholar] [CrossRef]
- Lieberman, R.L.; Shrestha, D.B.; Doan, P.E.; Hoffman, B.M.; Stemmler, T.L.; Rosenzweig, A.C. Purified particulate methane monooxygenase from Methylococcus capsulatus (Bath) is a dimer with both mononuclear copper and a copper-containing cluster. Proc. Natl. Acad. Sci. USA 2003, 100, 3820–3825. [Google Scholar] [CrossRef]
- Culpepper, M.A.; Rosenzweig, A.C. Architecture and active site of particulate methane monooxygenase. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.I.; Nguyen, H.-H.T.; Chen, K.H.-C.; Yu, S.S.-F. Overexpression and purification of the particulate methane monooxygenase from Methylococcus capsulatus (Bath). Methods Enzymol. 2011, 495, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Kunz, R.C.; Boyd, E.S.; Semrau, J.D.; Antholine, W.E.; Han, J.I.; Zahn, J.A.; Boyd, J.M.; Arlene, M.; Dispirito, A.A. The membrane-associated methane monooxygenase (pMMO) and pMMO-NADH: Quinone oxidoreductase complex from Methylococcus capsulatus Bath. J. Bacteriol. 2003, 185, 5755–5764. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.W.; Hammond, L.; Porter, E.G. The solubility of hydrocarbon gases in lipid bilayers. Chem. Phys. Lipids 1977, 20, 229–241. [Google Scholar] [CrossRef]
- Takeguchi, M.; Miyakawa, K.; Okura, I. Purification and properties of particulate methane monooxygenase from Methylosinus trichosporium OB3b. J. Mol. Catal. A Chem. 1998, 132, 145–153. [Google Scholar] [CrossRef]
- Zahn, J.A.; Dispirito, A.A. Membrane-associated methane monooxygenase from Methylococcus capsulatus (Bath). J. Bacteriol. 1996, 178, 1018–1029. [Google Scholar] [CrossRef] [Green Version]
- Dispirito, A.A.; Zahn, J.A.; Graham, D.W.; Kim, H.J.; Larive, C.K.; Derrick, T.S.; Cox, C.D.; Taylor, A. Copper-binding compounds from Methylosinus trichosporium OB3b. J. Bacteriol. 1998, 180, 3606–3613. [Google Scholar]
- El Ghazouani, A.; Baslé, A.; Gray, J.; Graham, D.W.; Firbank, S.J.; Dennison, C. Variations in methanobactin structure influences copper utilization by methane-oxidizing bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 8400–8404. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.I.; Chen, K.H.-C.; Yu, S.S.-F.; Chen, C.-L.; Kuo, S.S.-J. Toward delineating the structure and function of the particulate methane monooxygenase from methanotrophic bacteria. Biochemistry 2004, 43, 4421–4430. [Google Scholar] [CrossRef]
- Choi, D.W.; Antholine, W.E.; Do, Y.S.; Semrau, J.D.; Kisting, C.J.; Kunz, R.C.; Campbell, D.; Rao, V.; Hartsel, S.C.; Dispirito, A.A. Effect of methanobactin on the activity and electron paramagnetic resonance spectra of the membrane-associated methane monooxygenase in Methylococcus capsulatus Bath. Microbiology 2005, 151, 3417–3426. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Graham, D.W.; Dispirito, A.A.; Alterman, M.A.; Galeva, N.; Larive, C.K.; Asunskis, D.; Sherwood, P.M. Methanobactin, a copper-acquisition compound from methane-oxidizing bacteria. Science 2004, 305, 1612–1615. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Galeva, N.; Larive, C.K.; Alterman, M.; Graham, D.W. Purification and physical− chemical properties of methanobactin: A chalkophore from Methylosinus trichosporium OB3b. Biochemistry 2005, 44, 5140–5148. [Google Scholar] [CrossRef] [PubMed]
- El Ghazouani, A.; Basle, A.; Firbank, S.J.; Knapp, C.W.; Gray, J.; Graham, D.W.; Dennison, C. Copper-binding properties and structures of methanobactins from Methylosinus trichosporium OB3b. Inorg. Chem. 2011, 50, 1378–1391. [Google Scholar] [CrossRef]
- Dispirito, A.A.; Semrau, J.D.; Murrell, J.C.; Gallagher, W.H.; Dennison, C.; Vuilleumier, S. Methanobactin and the link between copper and bacterial methane oxidation. Microbiol. Mol. Biol. Rev. 2016, 80, 387–409. [Google Scholar] [CrossRef] [PubMed]
- Behling, L.A.; Hartsel, S.C.; Lewis, D.E.; Dispirito, A.A.; Choi, D.W.; Masterson, L.R.; Veglia, G.; Gallagher, W.H. NMR, mass spectrometry and chemical evidence reveal a different chemical structure for methanobactin that contains oxazolone rings. J. Am. Chem. Soc. 2008, 130, 12604–12605. [Google Scholar] [CrossRef] [PubMed]
- Murrell, J.C.; Mcdonald, I.R.; Gilbert, B. Regulation of expression of methane monooxygenases by copper ions. Trends Microbiol. 2000, 8, 221–225. [Google Scholar] [CrossRef]
- Kenney, G.E.; Rosenzweig, A.C. Chemistry and biology of the copper chelator methanobactin. ACS Chem. Biol. 2011, 7, 260–268. [Google Scholar] [CrossRef]
- Jeffery, C.J. Moonlighting proteins: Old proteins learning new tricks. Trends Genet. 2003, 19, 415–417. [Google Scholar] [CrossRef]
- Dmoore, B. Bifunctional and moonlighting enzymes: Lighting the way to regulatory control. Trends Plant. Sci. 2004, 9, 221–228. [Google Scholar] [CrossRef]
- Lu, Y.J.; Hung, M.C.; Chang, B.T.A.; Lee, T.L.; Lin, Z.H.; Tsai, I.K.; Chen, Y.S.; Chang, C.S.; Tsai, Y.F.; Chen, K.H.C.; et al. The PmoB subunit of particulate methane monooxygenase (pMMO) in Methylococcus capsulatus (Bath): The CuI sponge and its function. J. Inorg. Biochem. 2019, 196, 110691. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.W.; Do, Y.S.; Zea, C.J.; McEllistrem, M.T.; Lee, S.W.; Semrau, J.D.; Pohl, N.L.; Kisting, C.J.; Scardino, L.L.; Hartsel, S.C.; et al. Spectral and thermodynamic properties of Ag (I), Au (III), Cd (II), Co (II), Fe (III), Hg (II), Mn (II), Ni (II), Pb (II), U (IV), and Zn (II) binding by methanobactin from Methylosinus trichosporium OB3b. J. Inorg. Biochem. 2006, 100, 2150–2161. [Google Scholar] [CrossRef]
- Xin, J.Y.; Cheng, D.D.; Zhang, L.X.; Lin, K.; Fan, H.-C.; Wang, Y.; Xia, C.G. Methanobactin-mediated one-step synthesis of gold nanoparticles. Int. J. Mol. Sci. 2013, 14, 21676–21688. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.-H.T.; Nakagawa, K.H.; Hedman, B.; Hedman, B.; Elliott, S.J.; Lidstrom, M.E.; Hodgson, K.O.; Chan, S.I. X-ray absorption and EPR studies on the copper ions associated with the particulate methane monooxygenase from Methylococcus capsulatus (Bath). Cu (I) ions and their implications. J. Am. Chem. Soc. 1996, 118, 12766–12776. [Google Scholar] [CrossRef]
- Xin, J.; Dou, B.; Wang, Z.; Wang, Y.; Xia, C.; Liu, Z. Direct Electrochemistry of Methanobactin Functionalized Gold Nanoparticles on Au Electrode. J. Nanosci. Nanotechnol. 2018, 18, 4805–4813. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.N.; Lee, J.Y.; Wang, D.I. Uncovering the design rules for peptide synthesis of metal nanoparticles. J. Am. Chem. Soc. 2010, 132, 5677–5686. [Google Scholar] [CrossRef]
- Jv, Y.; Li, B.; Cao, R. Positively-charged gold nanoparticles as peroxidiase mimic and their application in hydrogen peroxide and glucose detection. Chem. Commun. 2010, 46, 8017–8019. [Google Scholar] [CrossRef]
- Xin, J.Y.; Li, C.Y.; Zhang, S.; Wang, Y.; Zhang, W.; Xia, C.G. Cu-induced assembly of methanobactin-modified gold nanoparticles and its peroxidase mimic activity. IET Nanobiotechnology 2018, 12, 915–921. [Google Scholar] [CrossRef]
- Xin, J.-Y.; Cui, J.-R.; Hu, X.-X.; Li, S.-B.; Xia, C.-G.; Zhu, L.-M.; Wang, Y.-Q. Particulate methane monooxygenase from Methylosinus trichosporium is a copper-containing enzyme. Biochem. Biophys. Res. Commun. 2002, 295, 182–186. [Google Scholar] [CrossRef]
- Müh, F.; Glöckner, C.; Hellmich, J.; Zouni, A. Light-induced quinone reduction in photosystem II. BBA-Bioenergetics 2012, 1817, 44–65. [Google Scholar] [CrossRef] [Green Version]
- Han, R.; Rempfer, K.; Zhang, M.; Dobbek, H.; Zouni, A.; Dau, H.; Luber, S. Investigating the Structure and Dynamics of Apo-Photosystem II. ChemCatChem 2019, 11, 4072–4080. [Google Scholar] [CrossRef]
- Mengele, A.K.; Seibold, G.M.; Eikmanns, B.J.; Rau, S. Coupling Molecular Photocatalysis to Enzymatic Conversion. ChemCatChem 2017, 9, 4369–4376. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Specific Activity (nmol/min·mg) | |
|---|---|---|
| Propene | Methane | |
| Membrane fractions of pMMO | 3.16 ± 0.600 | 2.65 ± 0.770 |
| pMMO–AuNP nanobiohybrids | 4.62 ± 1.033 | 3.37 ± 0.880 |
| Mb–Cu-modified pMMO–AuNP nanobiohybrids | 8.48 ± 0.579 | 6.44 ± 0.837 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xin, J.-Y.; Sun, L.-R.; Lin, H.-Y.; Zhang, S.; Xia, C.-G. Hybridization of Particulate Methane Monooxygenase by Methanobactin-Modified AuNPs. Molecules 2019, 24, 4027. https://doi.org/10.3390/molecules24224027
Xin J-Y, Sun L-R, Lin H-Y, Zhang S, Xia C-G. Hybridization of Particulate Methane Monooxygenase by Methanobactin-Modified AuNPs. Molecules. 2019; 24(22):4027. https://doi.org/10.3390/molecules24224027
Chicago/Turabian StyleXin, Jia-Ying, Li-Rui Sun, Hui-Ying Lin, Shuai Zhang, and Chun-Gu Xia. 2019. "Hybridization of Particulate Methane Monooxygenase by Methanobactin-Modified AuNPs" Molecules 24, no. 22: 4027. https://doi.org/10.3390/molecules24224027
APA StyleXin, J. -Y., Sun, L. -R., Lin, H. -Y., Zhang, S., & Xia, C. -G. (2019). Hybridization of Particulate Methane Monooxygenase by Methanobactin-Modified AuNPs. Molecules, 24(22), 4027. https://doi.org/10.3390/molecules24224027