
Astilbin Inhibits the Activity of Sortase A from Streptococcus mutans

Abstract
:1. Introduction
2. Results
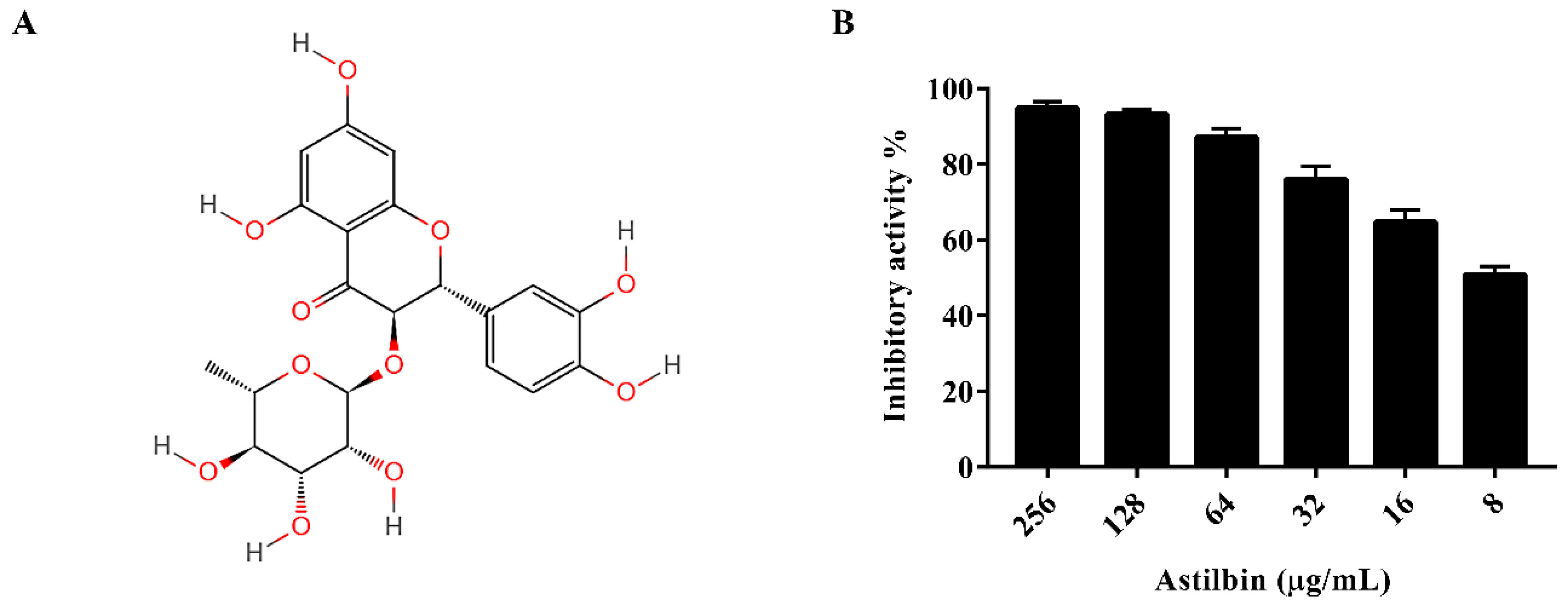
2.1. Inhibition of S. mutans SrtA by Astilbin
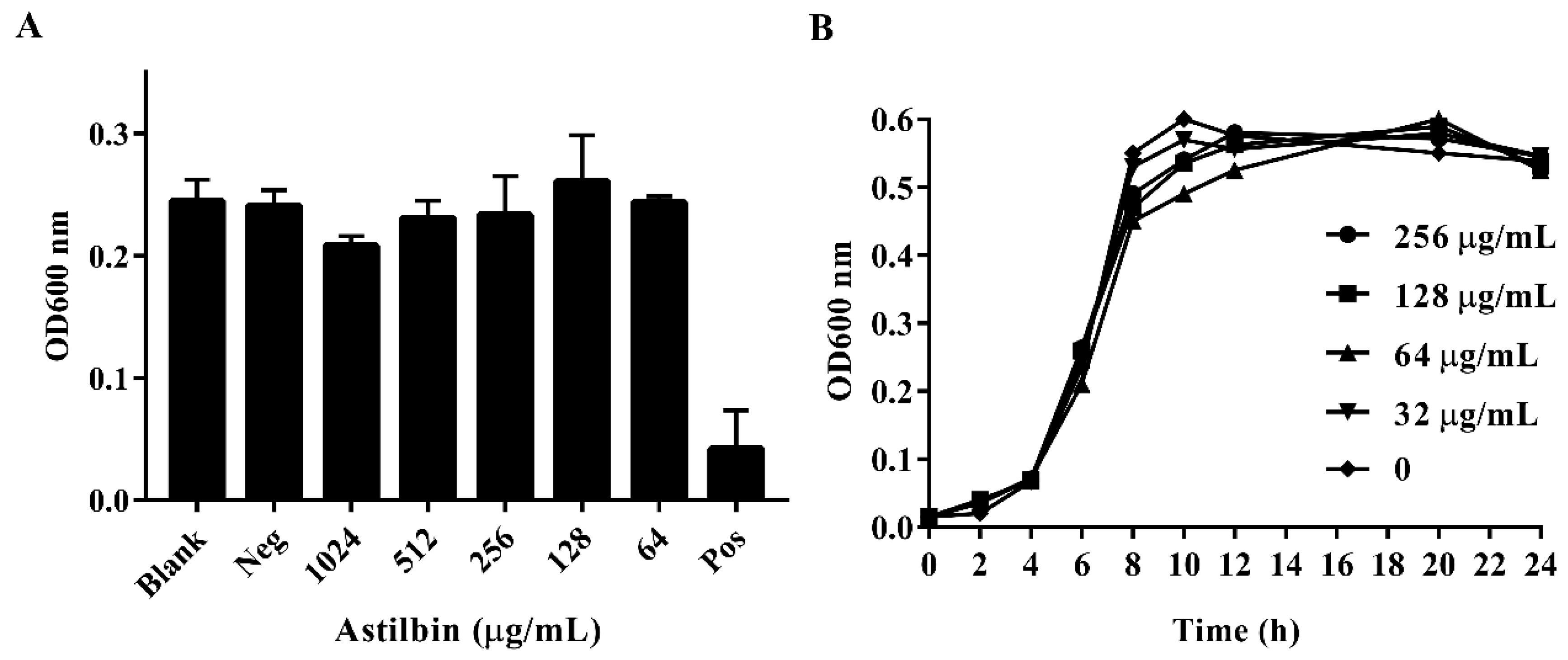
2.2. Antibacterial Activity of Astilbin
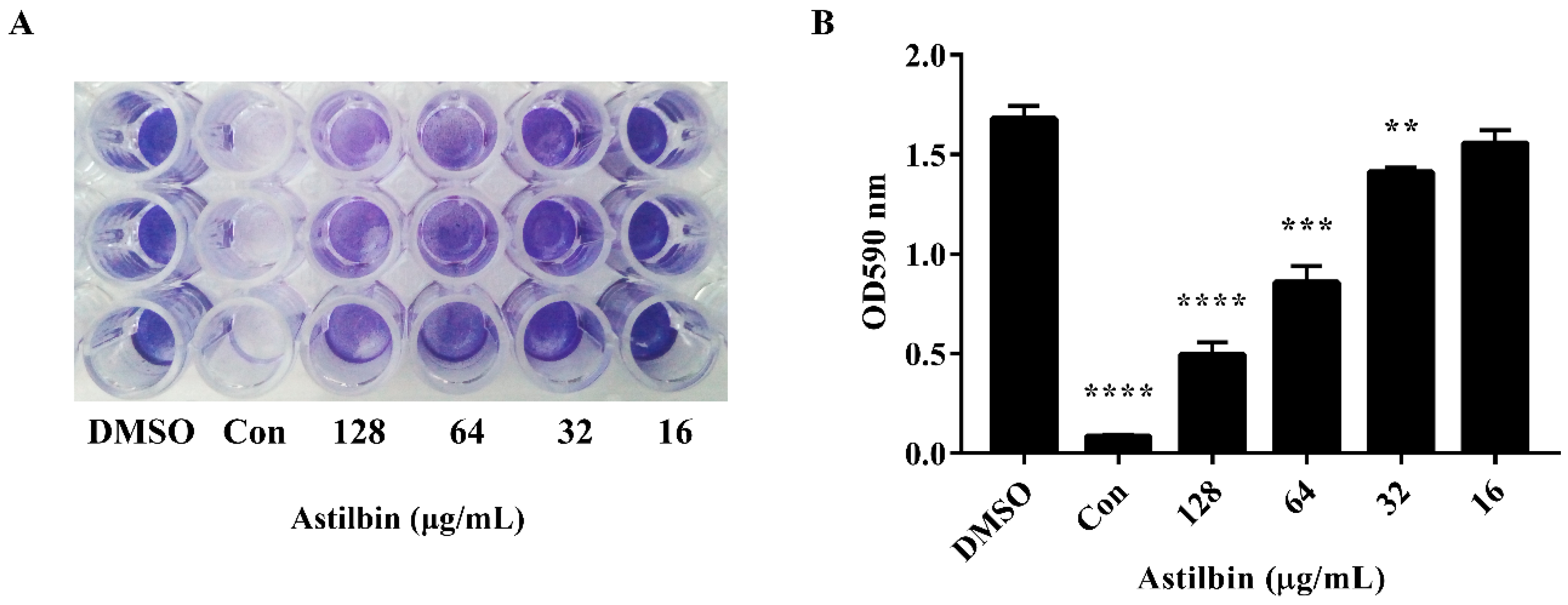
2.3. Inhibition of Biofilm Formation by Astilbin
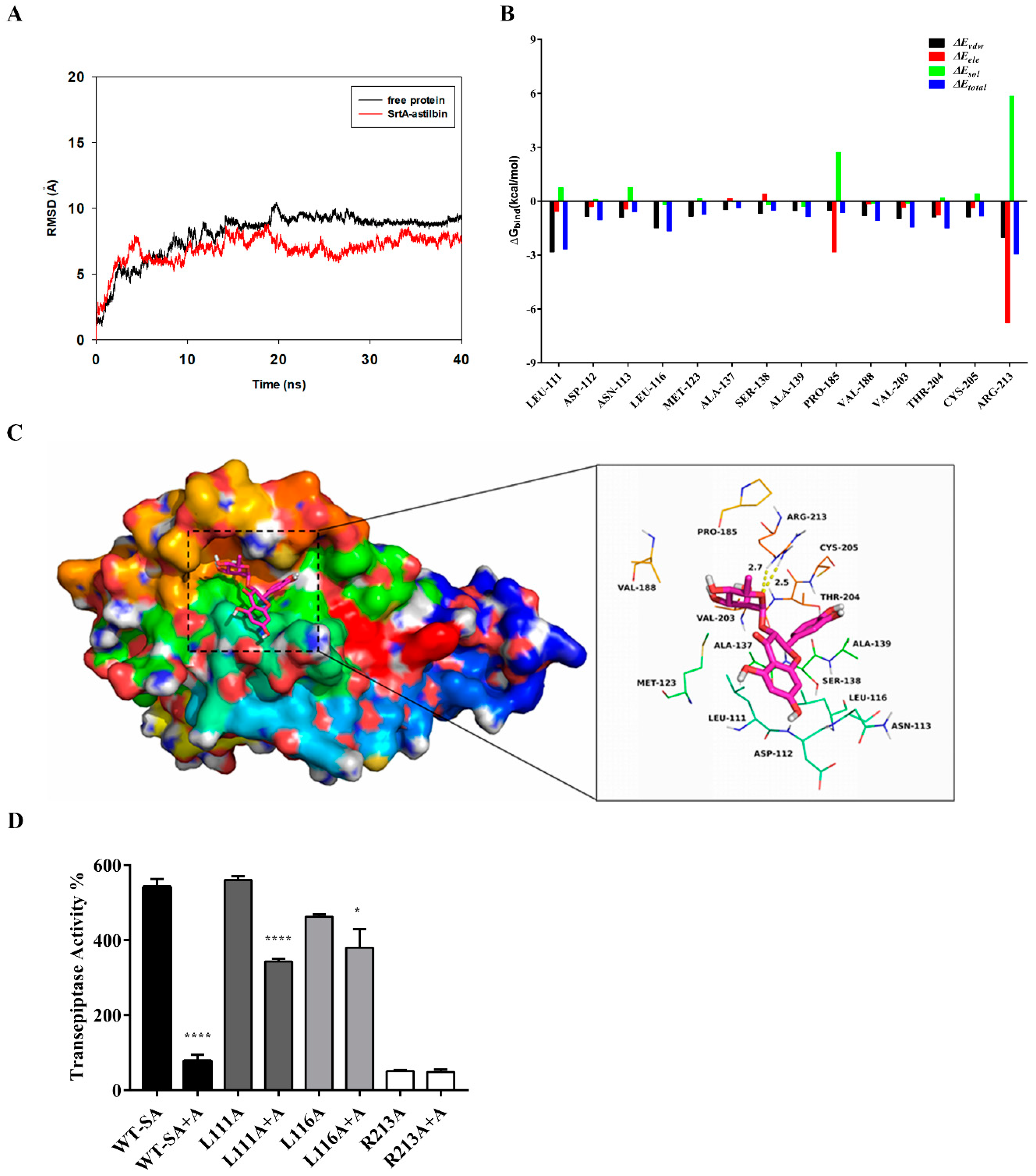
2.4. Determination of the Molecular Mechanism
3. Discussion
4. Materials and Methods
4.1. Bacteria, Chemicals, and Growth Conditions
4.2. Cloning and Expression of the SrtA and Its Mutants
4.3. Determination of MIC
4.4. Growth Curves of S. mutans
4.5. SrtA Activity Assay
4.6. Crystal Violet Biofilm Assay
4.7. Molecular Docking and Molecular Dynamics Simulation
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Van Houte, J. Role of micro-organisms in caries etiology. J. Dent. Res. 1994, 73, 672. [Google Scholar] [CrossRef] [PubMed]
- Kreth, J.; Merritt, J.; Qi, F. Bacterial and host interactions of oral streptococci. DNA Cell Biol. 2009, 28, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Nyvad, B. The role of bacteria in the caries process: Ecological perspectives. J. Dent. Res. 2011, 90, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Loesche, W.J. Role of Streptococcus mutans in human dental decay. Microbiol. Rev. 1986, 50, 353–380. [Google Scholar]
- Gendron, R.; Grenier, D.; Maheurobert, L. The oral cavity as a reservoir of bacterial pathogens for focal infections. Microbes Infect. 2000, 2, 897–906. [Google Scholar] [CrossRef]
- Banas, J.A.; Vickerman, M.M. Glucan-binding proteins of the oral streptococci. Crit. Rev. Oral. Biol. Med. 2003, 14, 89. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, H.F.; Demuth, D.R. Structure, function and immunogenicity of streptococcal antigen I/II polypeptides. Mol. Microbiol. 2010, 23, 183–190. [Google Scholar] [CrossRef]
- Ma, J.K.; Hunjan, M.; Smith, R.; Kelly, C.; Lehner, T. An investigation into the mechanism of protection by local passive immunization with monoclonal antibodies against Streptococcus mutans. Infect. Immun. 1990, 58, 3407–3414. [Google Scholar]
- AvilãS-Reyes, A.; Miller, J.H.; Lemos, J.A.; Abranches, J. Collagen-binding proteins of Streptococcus mutans and related streptococci. Mol. Oral Microbiol. 2017, 32, 89–106. [Google Scholar] [CrossRef]
- Haas, W.; Banas, J.A. Ligand-binding properties of the carboxyl-terminal repeat domain of Streptococcus mutans glucan-binding protein A. J. Bacteriol. 2000, 182, 728. [Google Scholar] [CrossRef]
- Sato, Y.; Yamamoto, Y.; Kizaki, H. Cloning and sequence analysis of the gbpC gene encoding a novel glucan-binding protein of Streptococcus mutans. Infect. Immun. 1997, 65, 668–675. [Google Scholar] [PubMed]
- Cossart, P.; Jonquières, R. Sortase, a universal target for therapeutic agents against Gram-positive bacteria? Proc. Natl. Acad. Sci. USA 2000, 97, 5013–5015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonthat, H.; Liu, G.; Mazmanian, S.K.; Faull, K.F.; Schneewind, O. Purification and characterization of sortase, the transpeptidase that cleaves surface proteins of Staphylococcus aureus at the LPXTG motif. Proc. Natl. Acad. Sci. USA 1999, 96, 12424–12429. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.F.; Boran, T.L. Roles of sortase in surface expression of the major protein adhesin P1, saliva-induced aggregation and adherence, and cariogenicity of Streptococcus mutans. Infect. Immun. 2003, 71, 676. [Google Scholar] [CrossRef] [PubMed]
- Maresso, A.W.; Ruiying, W.; Kern, J.W.; Rongguang, Z.; Dorota, J.; Missiakas, D.M.; Mark-Eugene, D.; Andrzej, J.; Olaf, S. Activation of inhibitors by sortase triggers irreversible modification of the active site. J. Biol. Chem. 2007, 282, 23129–23139. [Google Scholar] [CrossRef] [PubMed]
- Frankel, B.A.; Matthew, B.; Kruger, R.G.; Mccafferty, D.G. Vinyl sulfones: Inhibitors of SrtA, a transpeptidase required for cell wall protein anchoring and virulence in Staphylococcus aureus. J. Am. Chem. Soc. 2004, 126, 3404–3405. [Google Scholar] [CrossRef]
- Scott, C.J.; Andrew, M.D.; S Lorraine, M.; Lynas, J.F.; Koen, V.; Brian, W. Irreversible inhibition of the bacterial cysteine protease-transpeptidase sortase (SrtA) by substrate-derived affinity labels. Biochem. J. 2002, 366, 953–958. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, H.; Pan, J.; Dong, J.; Zhou, X.; Niu, X.; Deng, X. Oligopeptide Targeting Sortase A as Potential Anti-infective Therapy forStaphylococcus aureus. Front. Microbiol. 2018, 9, 245. [Google Scholar] [CrossRef]
- Liu, B.; Chen, F.; Bi, C.; Wang, L.; Zhong, X.; Cai, H.; Deng, X.; Niu, X.; Wang, D. Quercitrin, an Inhibitor of Sortase A, Interferes with the Adhesion of Staphylococcal aureus. Molecules 2015, 20, 6533–6543. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Teng, Z.; Li, M.; Niu, X.; Wang, J.; Deng, X. Epigallocatechin gallate inhibits Streptococcus pneumoniae virulence by simultaneously targeting pneumolysin and sortase A. J. Cell. Mol. Med. 2017, 21, 2586–2598. [Google Scholar] [CrossRef] [Green Version]
- Park, W.; Ahn, C.H.; Cho, H.; Kim, C.K.; Shin, J.; Oh, K.B. Inhibitory effects of flavonoids from Spatholobus suberectus on sortase A and sortase A-mediated aggregation of Streptococcus mutans. J. Microbiol. Biotechnol. 2017, 27, 1457–1460. [Google Scholar] [PubMed]
- Huang, P.; Hu, P.; Zhou, S.Y.; Li, Q.; Chen, W.M. Morin inhibits sortase A and subsequent biofilm formation in Streptococcus mutans. Curr. Microbiol. 2014, 68, 47–52. [Google Scholar] [CrossRef]
- Kuspradini, H.; Mitsunaga, T.; Ohashi, H. Antimicrobial activity against Streptococcus sobrinus and glucosyltransferase inhibitory activity of taxifolin and some flavanonol rhamnosides from kempas (Koompassia malaccensis) extracts. J. Wood Sci. 2009, 55, 308–313. [Google Scholar] [CrossRef]
- Huang, H.; Cheng, Z.; Shi, H. Isolation and Characterization of Two Flavonoids, Engeletin and Astilbin, from the Leaves of Engelhardia roxburghiana and Their Potential Anti-inflammatory Properties. J. Agric. Food Chem. 2011, 59, 4562–4569. [Google Scholar] [CrossRef]
- Zhang, Q.-F.; Zhang, Z.-R.; Cheung, H.-Y. Antioxidant activity of Rhizoma Smilacis Glabrae extracts and its key constituent-astilbin. Food Chem. 2009, 115, 297–303. [Google Scholar] [CrossRef]
- Jianming, G.; Feng, Q.; Jianxin, L.; Qiang, X.; Ting, C. Identification of a new metabolite of astilbin, 3’-O-methylastilbin, and its immunosuppressive activity against contact dermatitis. Clin. Chem. 2007, 53, 465–471. [Google Scholar]
- Matayoshi, E.D.; Wang, G.T.; Krafft, G.A.; Erickson, J. Novel fluorogenic substrates for assaying retroviral proteases by resonance energy transfer. Science 1990, 247, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Okahashi, N.; Sasakawa, C.; Yoshikawa, M.; Hamada, S.; Koga, T. Molecular characterization of a surface protein antigen gene from serotype c Streptococcus mutans, implicated in dental caries. Mol. Microbiol. 2010, 3, 673–678. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Skaar, E.P.; Gaspar, A.H.; Humayun, M.; Gornicki, P.; Jelenska, J.; Joachmiak, A.; Missiakas, D.M.; Schneewind, O. Passage of heme-iron across the envelope of Staphylococcus aureus. Science 2003, 299, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Bolken, T.C.; Franke, C.A.; Jones, K.F.; Zeller, G.O.; Jones, C.H.; Dutton, E.K.; Hruby, D.E. Inactivation of the srtA Gene in Streptococcus gordonii Inhibits Cell Wall Anchoring of Surface Proteins and Decreases In Vitro and In Vivo Adhesion. Infect. Immun. 2001, 69, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Lévesque, C.M.; Voronejskaia, E.; Huang, Y.C.; Mair, R.W.; Ellen, R.P.; Cvitkovitch, D.G. Involvement of sortase anchoring of cell wall proteins in biofilm formation by Streptococcus mutans. Infect. Immun. 2005, 73, 3773–3777. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Huang, P.; Chen, W.M. Curcumin inhibits the Sortase A activity of the Streptococcus mutans UA159. Appl. Biochem. Biotech. 2013, 171, 396–402. [Google Scholar]
- Wallock-Richards, D.J.; Jon, M.W.; Clarke, D.J.; Amarnath, M.; Michael, D.; Bryan, H.; Campopiano, D.J. Molecular basis of Streptococcus mutans sortase A inhibition by the flavonoid natural product trans-chalcone. Chem. Commun. 2015, 51, 10483–10485. [Google Scholar] [Green Version]
- Yang, W.Y.; Won, T.H.; Ahn, C.H.; Lee, S.H.; Yang, H.C.; Shin, J.; Oh, K.B. Streptococcus mutans sortase A inhibitory metabolites from the flowers of Sophora japonica. Bioorg. Med. Chem. Lett. 2015, 25, 1394–1397. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, E.J.; Citron, D.M.; Warren, Y.A.; Tyrrell, K.L.; Merriam, C.V.; Fernandez, H.T. In vitro activities of dalbavancin and 12 other agents against 329 aerobic and anaerobic gram-positive isolates recovered from diabetic foot infections. Antimicrob. Agents Chemother. 2006, 50, 2875. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Sebaugh, J.L. Guidelines for accurate EC50/IC50 estimation. Pharm. Stat. 2011, 10, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Islam, B.; Khan, S.N.; Naeem, A.; Sharma, V.; Khan, A.U. Novel effect of plant lectins on the inhibition of Streptococcus mutans biofilm formation on saliva-coated surface. J. App. Microbiol. 2010, 106, 1682–1689. [Google Scholar] [CrossRef]
- Oleg, T.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar]
- Götz, A.W.; Williamson, M.J.; Xu, D.; Poole, D.; Le Grand, S.; Walker, R.C. Routine microsecond molecular dynamics simulations with AMBER on GPUs. 1. Generalized born. J. Chem. Theory Comput. 2012, 8, 1542–1555. [Google Scholar] [CrossRef]
- Pierce, L.C.; Salomon-ferrer, R.; Augusto, F.d.O.C; Mccammon, J.A.; Walker, R.C. Routine Access to Millisecond Time Scale Events with Accelerated Molecular Dynamics. J. Chem. Theory Comput. 2012, 8, 2997–3002. [Google Scholar] [CrossRef] [PubMed]
- Salomon-ferrer, R.; Götz, A.W.; Poole, D.; Le, G.S.; Walker, R.C. Routine Microsecond Molecular Dynamics Simulations with AMBER on GPUs. 2. Explicit Solvent Particle Mesh Ewald. J. Chem. Theory Comput. 2013, 9, 3878–3888. [Google Scholar] [CrossRef] [PubMed]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An overview of the amber biomolecular simulation package. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2013, 3, 198–210. [Google Scholar] [CrossRef]
Sample Availability: Not available. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequences (5′–3′) |
|---|---|
| SmsrtA-F | CGCGGATCCATGAAAAAAGAACGTCAATCTAGGA |
| SmsrtA-R | CCGCTCGAGTTAAAATGATATTTGATTATAGGACTGCC |
| R213A-SrtA-F | TTGTTCATGGCACATATAAGGGGGAA |
| R213A-SrtA-R | TTGTTGCAGCAGTCGCCC |
| L111A-SrtA-F | AAGGAGCAGATAATGTTGGCTTAAC |
| L111A-SrtA-R | TGAAGATTGGTAAATTGATTTTTAAGTCTGG |
| L116A-SrtA-F | TTGGCGCAACATATGGTG |
| L116A-SrtA-R | ATTTATCTAATCCTTTGAAGATTGGTAAATTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Shi, Y.; Jing, S.; Dong, H.; Wang, D.; Wang, T. Astilbin Inhibits the Activity of Sortase A from Streptococcus mutans. Molecules 2019, 24, 465. https://doi.org/10.3390/molecules24030465
Wang J, Shi Y, Jing S, Dong H, Wang D, Wang T. Astilbin Inhibits the Activity of Sortase A from Streptococcus mutans. Molecules. 2019; 24(3):465. https://doi.org/10.3390/molecules24030465
Chicago/Turabian StyleWang, Junxian, Yan Shi, Shisong Jing, Haisi Dong, Dacheng Wang, and Tiedong Wang. 2019. "Astilbin Inhibits the Activity of Sortase A from Streptococcus mutans" Molecules 24, no. 3: 465. https://doi.org/10.3390/molecules24030465
APA StyleWang, J., Shi, Y., Jing, S., Dong, H., Wang, D., & Wang, T. (2019). Astilbin Inhibits the Activity of Sortase A from Streptococcus mutans. Molecules, 24(3), 465. https://doi.org/10.3390/molecules24030465