Momilactones A and B Are α-Amylase and α-Glucosidase Inhibitors

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
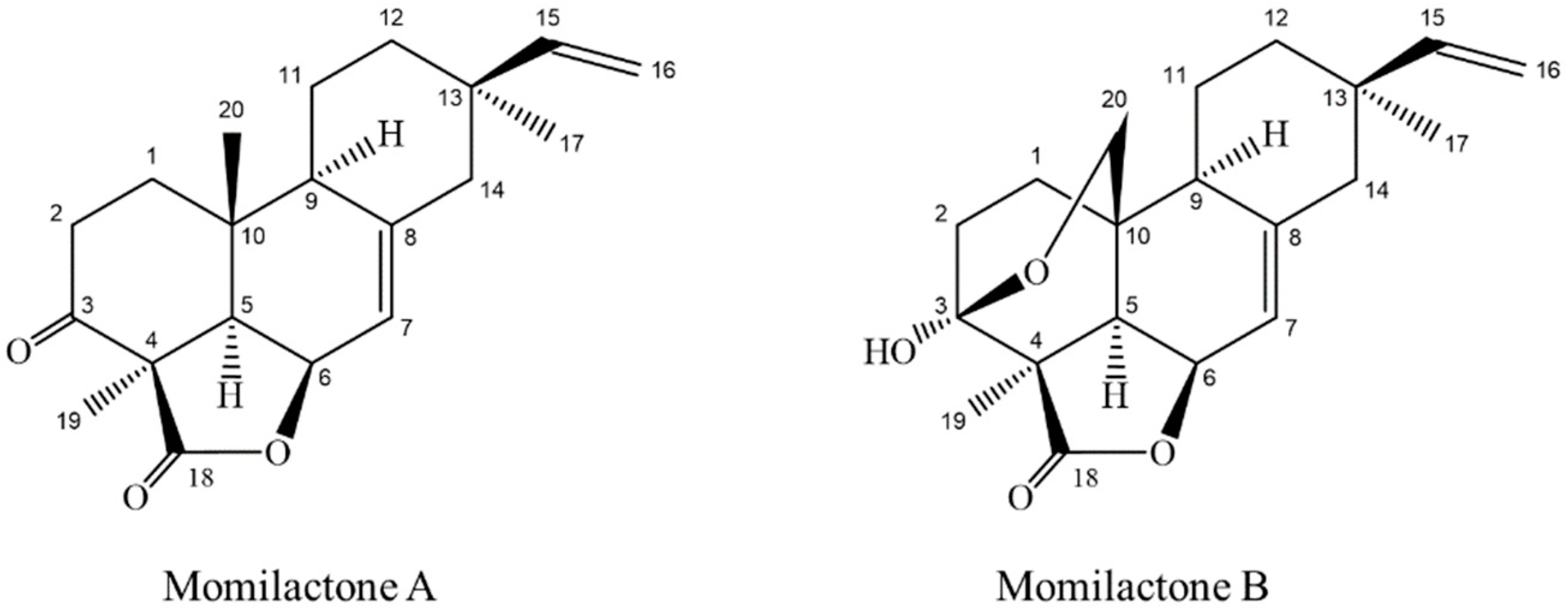
2.1. Isolation and Confirmation of Momilactones A and B
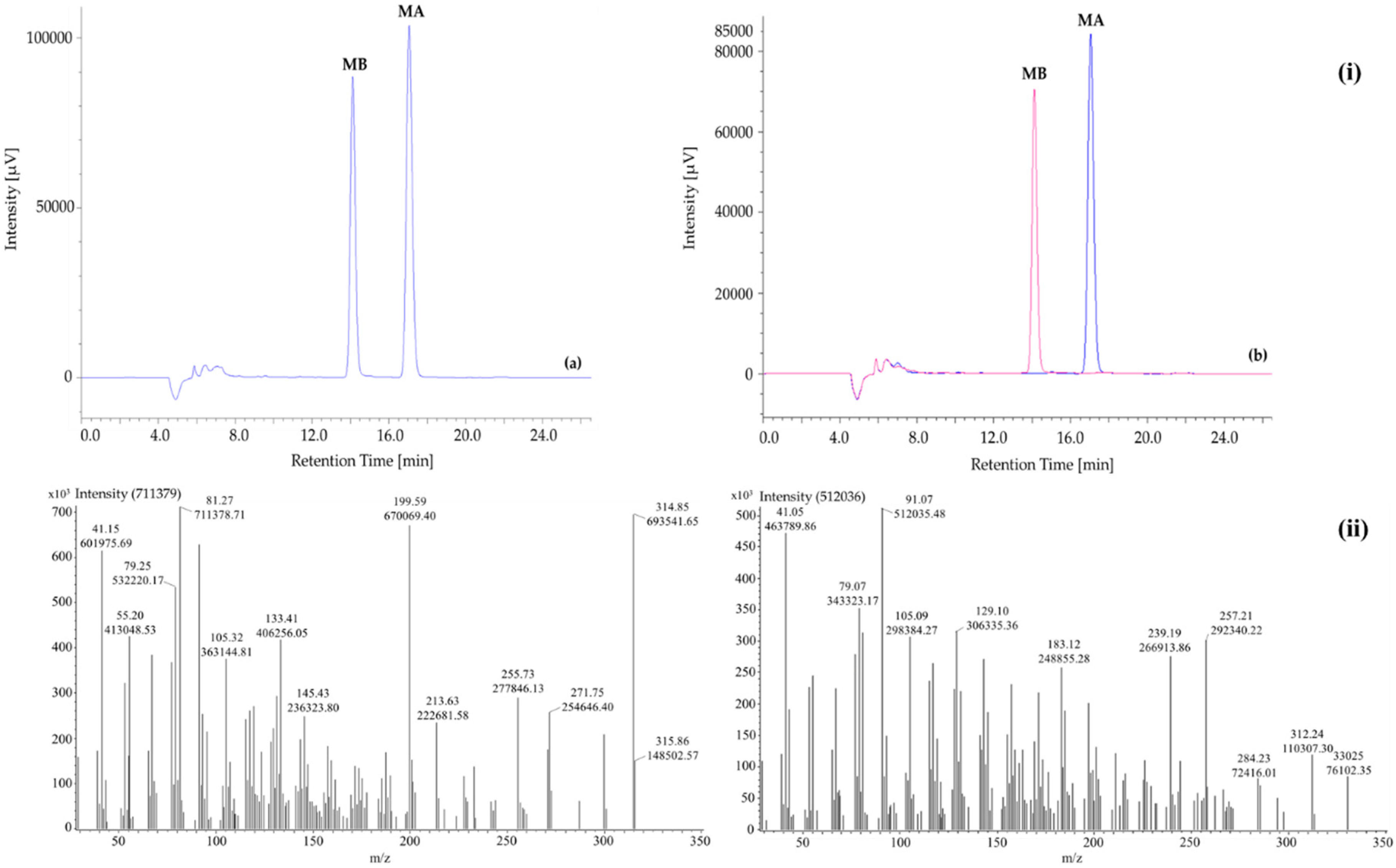
2.1.1. HPLC
2.1.2. GC-MS
2.1.3. 1H-NMR and 13C-NMR
2.2. In Vitro Inhibition of α-Amylase and α-Glucosidase
2.3. Contents of MA and MB in Rice Plant Parts
3. Discussion
4. Materials and Methods
4.1. Collection and Extraction of Rice Husk
4.2. Isolation of Momilactones A and B from EtOAc Extract
4.3. Identification and Confirmation of Momilactones A and B by HPLC, TLC, GC-MS, and 1H-NMR and 13C-NMR Analyses
4.4. α-Amylase Inhibition Assay
4.5. α-Glucosidase Inhibition Assay
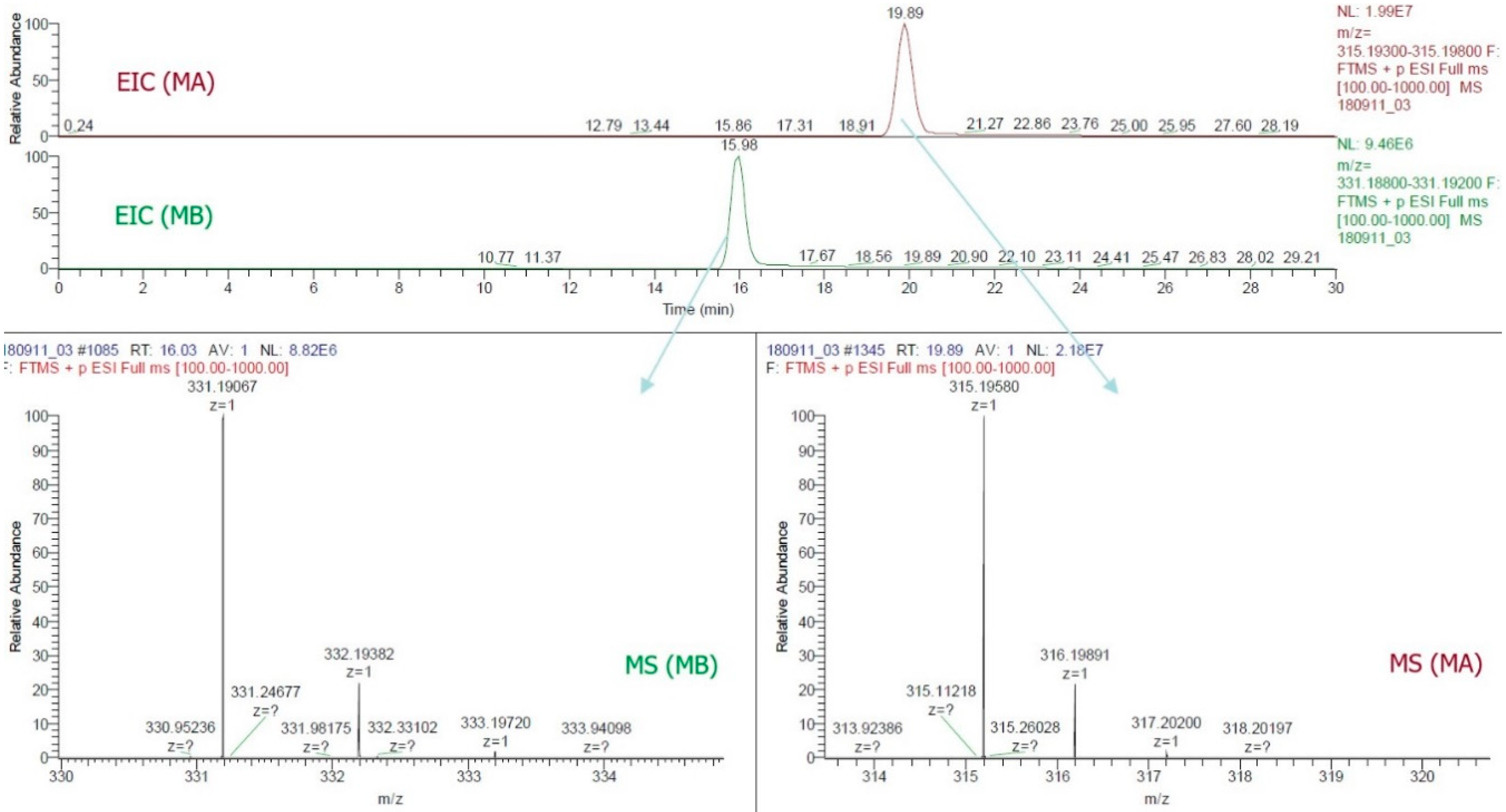
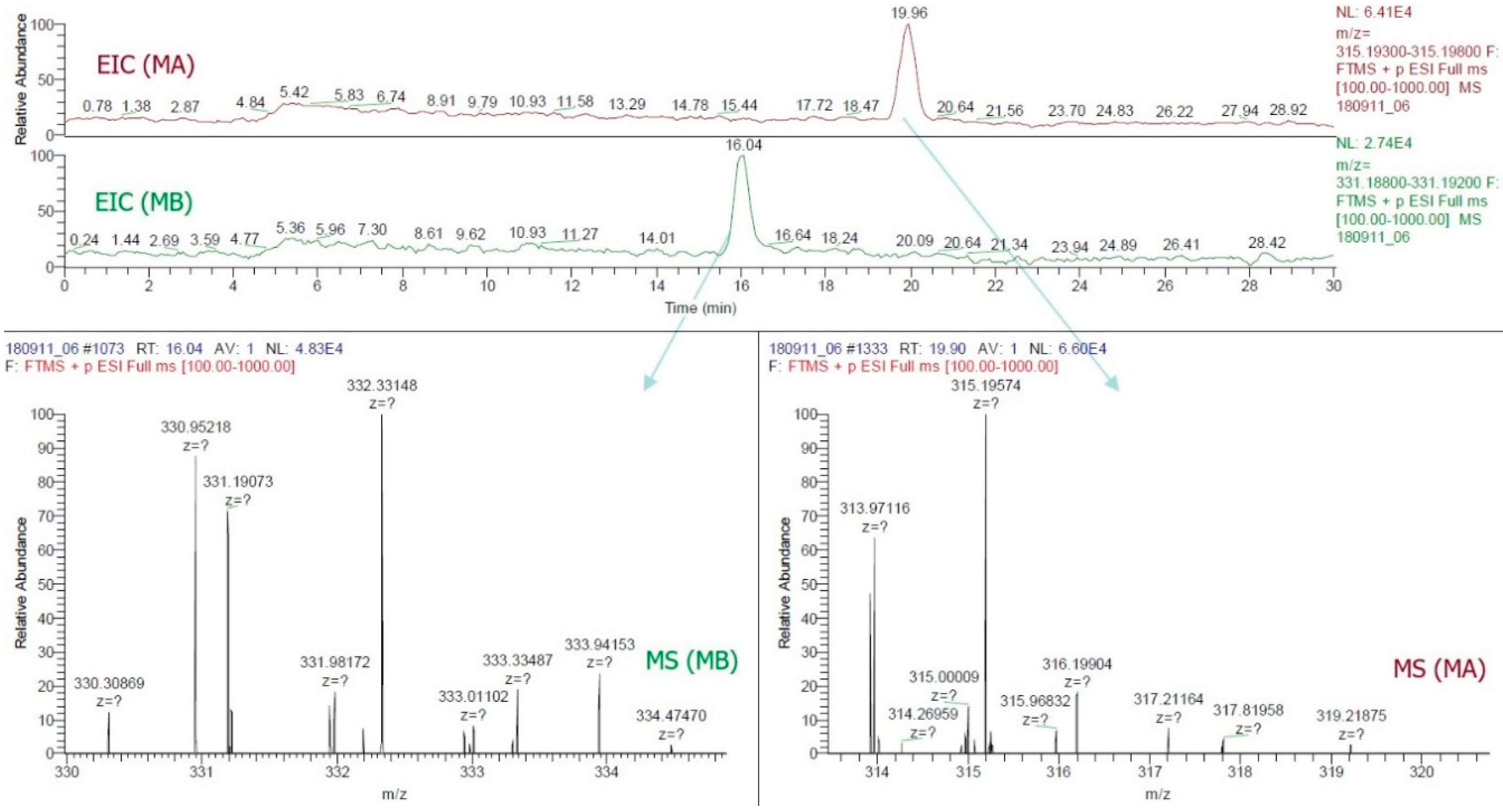
4.6. Quantification and Confirmation of MA and MB in Rice Plant Parts by HPLC and LC-ESI-MS
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IDF Diabetes Atlas—8th Edition. Available online: http://diabetesatlas.org/key-messages.html (accessed on 1 August 2018).
- Leahy, J.L. Pathogenesis of type 2 diabetes mellitus. Arch. Med. Res. 2005, 36, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Abesundara, K.J.M.; Matsui, T.; Matsumoto, K. α-Glucosidase inhibitory activity of some Sri Lanka plant extracts, one of which, Cassia auriculata, exerts a strong antihyperglycemic effect in rats comparable to the therapeutic drug acarbose. J. Agric. Food Chem. 2004, 52, 2541–2545. [Google Scholar] [CrossRef] [PubMed]
- Ercan, P.; El, S.N. Inhibitory effects of chickpea and Tribulus terrestris on lipase, α-amylase and α-glucosidase. Food Chem. 2016, 205, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Apostolidis, E.; Kwon, Y.I.; Shetty, K. Inhibitory potential of herb, fruit, and fungal-enriched cheese against key enzymes linked to type 2 diabetes and hypertension. Innov. Food Sci. Emerg. Technol. 2007, 8, 46–54. [Google Scholar] [CrossRef]
- Arulselvan, P.; Ghofar, H.A.A.; Karthivashan, G.; Halim, M.F.A.; Ghafar, M.S.A.; Fakurazi, S. Antidiabetic therapeutics from natural source: A systematic review. Biomed. Prev. Nutr. 2014, 4, 607–617. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Convergent or parallel molecular evolution of momilactone A and B: Potent allelochemicals, momilactones have been found only in rice and the moss Hypnum plumaeforme. J. Plant Physiol. 2011, 168, 1511–1516. [Google Scholar] [CrossRef]
- Minh, T.N.; Xuan, T.D.; Ahmad, A.; Elzaawely, A.A.; Teschke, R.; Van, T.M. Momilactones A and B: Optimization of yields from isolation and purification. Separations 2018, 5, 28. [Google Scholar] [CrossRef]
- Toyomasu, T.; Kagahara, T.; Okada, K.; Koga, J.; Hasegawa, M.; Mitsuhashi, W.; Sassa, T.; Yamane, H. Diterpene phytoalexins are biosynthesized in and exuded from the roots of rice seedlings. Biosci. Biotechnol. Biochem. 2008, 72, 562–567. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Hasegawa, M.; Ino, T.; Ota, K.; Kujime, H. Contribution of momilactone A and B to rice allelopathy. J. Plant Physiol. 2010, 167, 787–791. [Google Scholar] [CrossRef]
- Chung, I.M.; Jung, T.K.; Kim, S.H. Evaluation of allelopathic potential and quantification of momilactone A,B from rice hull extracts and assessment of inhibitory bioactivity on paddy field weeds. J. Agric. Food Chem. 2006, 54, 2527–2536. [Google Scholar] [CrossRef]
- Cartwright, D.W.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Isolation and characterization of two phytoalexins from rice as momilactones A and B. Phytochemistry 1981, 20, 535–537. [Google Scholar] [CrossRef]
- Obara, N.; Hasegawa, M.; Kodama, O. Induced volatiles in elicitor-treated and rice blast fungus-inoculated rice leaves. Biosci. Biotechnol. Biochem. 2002, 66, 2549–2559. [Google Scholar] [CrossRef] [PubMed]
- Fukuta, M.; Xuan, T.D.; Deba, F.; Tawata, S.; Khanh, T.D.; Chung, I.M. Comparative efficacies in vitro of antibacterial, fungicidal, antioxidant, and herbicidal activities of momilatones A and B. J. Plant Interact. 2007, 2, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Xuan, T.D.; Minh, T.N.; Anh, L.H.; Khanh, T.D. Allelopathic momilactones A and B are implied in rice drought and salinity tolerance, not weed resistance. Agron. Sustain. Dev. 2016, 36, 52. [Google Scholar] [CrossRef]
- Quan, N.T.; Xuan, T.D. Foliar application of vanillic and p-hydroxybenzoic acids enhanced drought tolerance and formation of phytoalexin momilactones in rice. Arch. Agron. Soil Sci. 2018, 1–16. [Google Scholar] [CrossRef]
- Chung, I.M.; Ali, M.; Hahn, S.J.; Siddiqui, N.A.; Lim, Y.H.; Ahmad, A. Chemical constituents from the hulls of Oryza sativa with cytotoxic activity. Chem. Nat. Compd. 2005, 41, 182–189. [Google Scholar] [CrossRef]
- Kim, S.J.; Park, H.R.; Park, E.; Lee, S.C. Cytotoxic and antitumor activity of momilactone B from rice hulls. J. Agric. Food Chem. 2007, 55, 1702–1706. [Google Scholar] [CrossRef]
- Joung, Y.H.; Lim, E.J.; Kim, M.S.; Lim, S.D.; Yoon, S.Y.; Lim, Y.C.; Yoo, Y.B.; Ye, S.K.; Park, T.; Chung, I.M.; et al. Enhancement of hypoxia-induced apoptosis of human breast cancer cells via STAT5b by momilactone B. Int. J. Oncol. 2008, 33, 477–484. [Google Scholar] [CrossRef]
- Park, C.; Jeong, N.Y.; Kim, G.Y.; Han, M.H.; Chung, I.M.; Kim, W.J.; Yoo, Y.H.; Choi, Y.H. Momilactone B induces apoptosis and G1 arrest of the cell cycle in human monocytic leukemia U937 cells through downregulation of pRB phosphorylation and induction of the cyclin-dependent kinase inhibitor p21Waf1/Cip1. Oncol. Rep. 2014, 31, 1653–1660. [Google Scholar] [CrossRef] [Green Version]
- Minh, T.N.; Xuan, T.D.; Ahmad, A.; Elzaawely, A.A.; Teschke, R.; Van, T.M. Efficacy from different extractions for chemical profile and biological activities of rice husk. Sustainability 2018, 10, 1356. [Google Scholar] [CrossRef]
- Kang, D.Y.; Nipin, S.P.; Darvin, P.; Joung, Y.H.; Byun, H.J.; Do, C.H.; Park, K.D.; Park, M.N.; Cho, K.H.; Yang, Y.M. Momilactone B inhibits ketosis in vitro by regulating the ANGPTL3-LPL pathway and inhibiting HMGCS2. Anim. Biotechnol. 2017, 28, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Eliza, J.; Daisy, P.; Ignacimuthu, S.; Duraipandiyan, V. Normo-glycemic and hypolipidemic effect of costunolide isolated from Costus specious (Koen ex Retz) Sm. in streptozotocin-induced diabetic rats. Chem. Biol. Interact. 2009, 179, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.C.; Hung, C.R.; Chen, W.C.; Cheng, J.T. Antihyperglycemic effect of andrographolide in streptozotocin-induced diabetic rats. Planta Med. 2003, 69, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, R.; Asmawi, M.Z.; Sadikun, A. In vitro α-glucosidase and α-amylase enzyme inhibitory effects of Andrographis paniculata extract and andrographolide. Acta Biochim. Pol. 2008, 55, 391–398. [Google Scholar] [PubMed]
- Brahmachari, G. Andrographolide: A Molecule of Antidiabetic Promise. In Discovery and Development of Antidiabetic Agents from Natural Products; Brahmachari, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–27. ISBN 978-0-12-809450-1. [Google Scholar]
- Xu, H.W.; Liu, G.Z.; Dai, G.F.; Wu, C.L.; Liu, H.M. Modification of 15-akylidene andrographolide derivatives as alpha-glucosidase inhibitor. Drug Discov. Ther. 2007, 1, 73–77. [Google Scholar] [PubMed]
- Reddy, P.P.; Tiwari, A.K.; Ranga Rao, R.; Madhusudhana, K.; Rama Subba Rao, V.; Ali, A.Z.; Suresh Babu, K.; Madhusudana Rao, J. New Labdane diterpenes as intestinal α-glucosidase inhibitor from antihyperglycemic extract of Hedychium spicatum (Ham. Ex Smith) rhizomes. Bioorg. Med. Chem. Lett. 2009, 19, 2562–2565. [Google Scholar] [CrossRef] [PubMed]
- Ayinampudi, S.R.; Domala, R.; Merugu, R.; Bathula, S.; Janaswamy, M.R. New icetexane diterpenes with intestinal α-glucosidase inhibitory and free-radical scavenging activity isolated from Premna tomentosa roots. Fitoterapia 2012, 83, 88–92. [Google Scholar] [CrossRef]
- Li, G.; Ding, W.; Wan, F.; Li, Y. Two new clerodane diterpenes from Tinospora sagittata. Molecules 2016, 21, 1250. [Google Scholar] [CrossRef]
- Ghosh, S.; Rangan, L. Molecular docking and inhibition studies of α-amylase activity by labdane diterpenes from Alpinia nigra seeds. Med. Chem. Res. 2014, 23, 4836–4852. [Google Scholar] [CrossRef]
- Jelenković, L.; Jovanović, V.S.; Palić, I.; Mitić, V.; Radulović, M. In vitro screening of α-amylase inhibition by selected terpenes from essential oils. Trop. J. Pharm. Res. 2014, 13, 1421–1428. [Google Scholar] [CrossRef]
- Chiou, S.Y.; Lai, J.Y.; Liao, J.A.; Sung, J.M.; Lin, S.D. In vitro inhibition of lipase, α-amylase, α-glucosidase, and angiotensin-converting enzyme by defatted rice bran extracts of red-pericarp rice mutant. Cereal Chem. 2017, 95, 167–176. [Google Scholar] [CrossRef]
- Uraipong, C.; Zhao, J. Rice bran protein hydrolysates exhibit strong in vitro α-amylase, β-glucosidase and ACE-inhibition activities. J. Sci. Food Agric. 2016, 96, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Boue, S.M.; Daigle, K.W.; Chen, M.H.; Cao, H.; Heiman, M.L. Antidiabetic potential of purple and red rice (Oryza sativa L.) bran extracts. J. Agric. Food Chem. 2016, 64, 5345–5353. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.H.; Huang, H.Y.; Chen, Y.Y.; Huang, C.L.; Chang, C.J.; Chen, H.L.; Lai, M.H. Ameliorative effects of stabilized rice bran on type 2 diabetes patients. Ann. Nutr. Metab. 2010, 56, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Xuan, T.D.; Minh, T.N.; Ali Siddiqui, N.; Quan, N.V. Comparative extraction and simple isolation improvement techniques of active constituents’ momilactone A and B from rice husks of Oryza sativa by HPLC analysis and column chromatography. Saudi Pharm. J. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Yoneyama, K.; Takeuchi, Y.; Konnai, M.; Tamogami, S.; Kodama, O. Momilactones A and B in rice straw harvested at different growth stages. Biosci. Biotechnol. Biochem. 1999, 63, 1318–1320. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, D.W.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Chemical activation of host defence mechanisms as a basis for crop protection. Nature 1977, 267, 511–513. [Google Scholar] [CrossRef]
- Yin, Z.; Zhang, W.; Feng, F.; Zhang, Y.; Kang, W. α-Glucosidase inhibitors isolated from medicinal plants. Food Sci. Hum. Wellness 2014, 3, 136–174. [Google Scholar] [CrossRef] [Green Version]
- Teschke, R.; Xuan, T.D. Viewpoint: A contributory role of shell ginger (Alpinia zerumbet) for human longevity in Okinawa, Japan? Nutrients 2018, 10, 166. [Google Scholar] [CrossRef]
- Xuan, T.D.; Khanh, T.D.; Khang, D.T.; Quan, N.T.; Ezaawey, A.A. Changes in chemical composition, total phenolics and antioxidant activity of Alpinia (Alpinia zerumbet) leaves exposed to UV. Intl. Let. Nat. Sci. 2016, 55, 25–34. [Google Scholar] [CrossRef]
- Xuan, T.D.; Teschke, R. Dihydro-5, 6-dehydrokavain (DDK) from Alpinia zerumbet: Its isolation, synthesis, and characterization. Molecules 2015, 20, 16306–16319. [Google Scholar] [CrossRef] [PubMed]
- Van, T.M.; Xuan, T.D.; Minh, T.N.; Quan, N.V. Isolation and purification of potent growth inhibitors from Piper methysticum root. Molecules 2018, 23, 1907. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Kang, M.Y.; Nam, S.H.; Friedman, M. Antidiabetic effects of rice hull smoke extract in alloxan-induced diabetic mice. J. Agric. Food Chem. 2012, 60, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R.S.; Saleh, A.M. Antifungal activity of rice straw extract on some phytopathogenic fungi. Afr. J. Biotechnol. 2012, 11, 13586–13590. [Google Scholar] [CrossRef]
- Hu, E.A.; Pan, A.; Malik, V.; Sun, Q. White rice consumption and risk of type 2 diabetes: Meta-analysis and systematic review. BMJ 2012, 344, e1454. [Google Scholar] [CrossRef]
- Sun, Q.; Spiegelman, D.; Van Dam, R.M.; Holmes, M.D.; Malik, V.; Willett, W.C.; Hu, F.B. White rice, brown rice, and risk of type 2 diabetes in US men and women. Arch. Intern. Med. 2011, 140, 961–969. [Google Scholar] [CrossRef]
- Zhang, G.; Malik, V.S.; Pan, A.; Kumar, S.; Holmes, M.D.; Spiegelman, D.; Lin, X.; Hu, F.B. Substituting brown rice for white rice to lower diabetes risk: A focus-group study in Chinese adults. J. Am. Diet. Assoc. 2010, 110, 1216–1221. [Google Scholar] [CrossRef]
- Kozuka, C.; Yabiku, K.; Takayama, C.; Matsushita, M.; Shimabukuro, M.; Masuzaki, H. Natural food science based novel approach toward prevention and treatment of obesity and type 2 diabetes: Recent studies on brown rice and γ-oryzanol. Obes. Res. Clin. Pract. 2013, 7, e165–e172. [Google Scholar] [CrossRef]
- Shao, Y.; Bao, J. Polyphenols in whole rice grain: Genetic diversity and health benefits. Food Chem. 2015, 180, 86–97. [Google Scholar] [CrossRef]
- Yawadio, R.; Tanimori, S.; Morita, N. Identification of phenolic compounds isolated from pigmented rices and their aldose reductase inhibitory activities. Food Chem. 2007, 101, 1616–1625. [Google Scholar] [CrossRef]
- Yao, Y.; Sang, W.; Zhou, M.; Ren, G. Antioxidant and α-glucosidase inhibitory activity of colored grains in China. J. Agric. Food Chem. 2010, 58, 770–774. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kwon, C.S.; Son, H.S. Inhibition of alpha-glucosidase and amylase by luteolin, a flavonoid. Biosci. Biotechnol. Biochem. 2000, 64, 2458–2461. [Google Scholar] [CrossRef] [PubMed]
- Hanh, H.; Tran, T.; Nguyen, M.C.; Le, H.T.; Nguyen, T.L.; Pham, T.B.; Chau, V.M.; Nguyen, H.N.; Nguyen, T.D. Inhibitors of α-glucosidase and α-amylase from Cyperus rotundus. Pharm. Biol. 2014, 52, 74–77. [Google Scholar] [CrossRef]
- Jhong, C.; Chia, Y. Screening alpha-glucosidase and alpha-amylase inhibitors from natural compounds by molecular docking in silico. Biofactors 2015, 41, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Hahn, S.J.; Ahmad, A. Confirmation of potential herbicidal agents in hulls of rice, Oryza sativa. J. Chem. Ecol. 2005, 31, 1339–1352. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H.; Lucius, A.; Meyer, T.; Gonzalez De Mejia, E. Cultivar evaluation and effect of fermentation on antioxidant capacity and in vitro inhibition of α-amylase and α-glucosidase by highbush blueberry (Vaccinium corombosum). J. Agric. Food Chem. 2011, 59, 8923–8930. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the pure momilactones A and B are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α-Amylase Inhibitory Assay (µg/mL) (IC50) | α-Glucosidase Inhibitory Assay (µg/mL) (IC50) | |
|---|---|---|
| MA | 266.68 ± 1.58 c | 991.95 ± 0.96 c |
| MB | 146.85 ± 1.12 b | 612.03 ± 0.39 b |
| Acarbose | 117.08 ± 0.85 a | 2549.00 ± 5.15 d |
| Quercetin | - | 105.68 ± 0.09 a |
| Rice Organs | Momilactone A | Momilactone B |
|---|---|---|
| Grain | 2.07 ± 0.01 d | 1.06 ± 0.01 d |
| Husk | 16.44 ± 0.09 a | 9.24 ± 0.04 b |
| Leaf | 4.28 ± 0.03 c | 12.73 ± 0.36 a |
| Root | 8.06 ± 0.13 b | 5.69 ± 0.19 c |
| Two-Way ANOVA | ||
| Momilactones | * | * |
| Rice organs | * | * |
| Interaction | * | * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, N.V.; Tran, H.-D.; Xuan, T.D.; Ahmad, A.; Dat, T.D.; Khanh, T.D.; Teschke, R. Momilactones A and B Are α-Amylase and α-Glucosidase Inhibitors. Molecules 2019, 24, 482. https://doi.org/10.3390/molecules24030482
Quan NV, Tran H-D, Xuan TD, Ahmad A, Dat TD, Khanh TD, Teschke R. Momilactones A and B Are α-Amylase and α-Glucosidase Inhibitors. Molecules. 2019; 24(3):482. https://doi.org/10.3390/molecules24030482
Chicago/Turabian StyleQuan, Nguyen Van, Hoang-Dung Tran, Tran Dang Xuan, Ateeque Ahmad, Tran Dang Dat, Tran Dang Khanh, and Rolf Teschke. 2019. "Momilactones A and B Are α-Amylase and α-Glucosidase Inhibitors" Molecules 24, no. 3: 482. https://doi.org/10.3390/molecules24030482
APA StyleQuan, N. V., Tran, H. -D., Xuan, T. D., Ahmad, A., Dat, T. D., Khanh, T. D., & Teschke, R. (2019). Momilactones A and B Are α-Amylase and α-Glucosidase Inhibitors. Molecules, 24(3), 482. https://doi.org/10.3390/molecules24030482