
Short-Term Neonatal Oral Administration of Oleanolic Acid Protects against Fructose-Induced Oxidative Stress in the Skeletal Muscles of Suckling Rats

 , , and
, , and
Abstract
:1. Introduction
2. Results
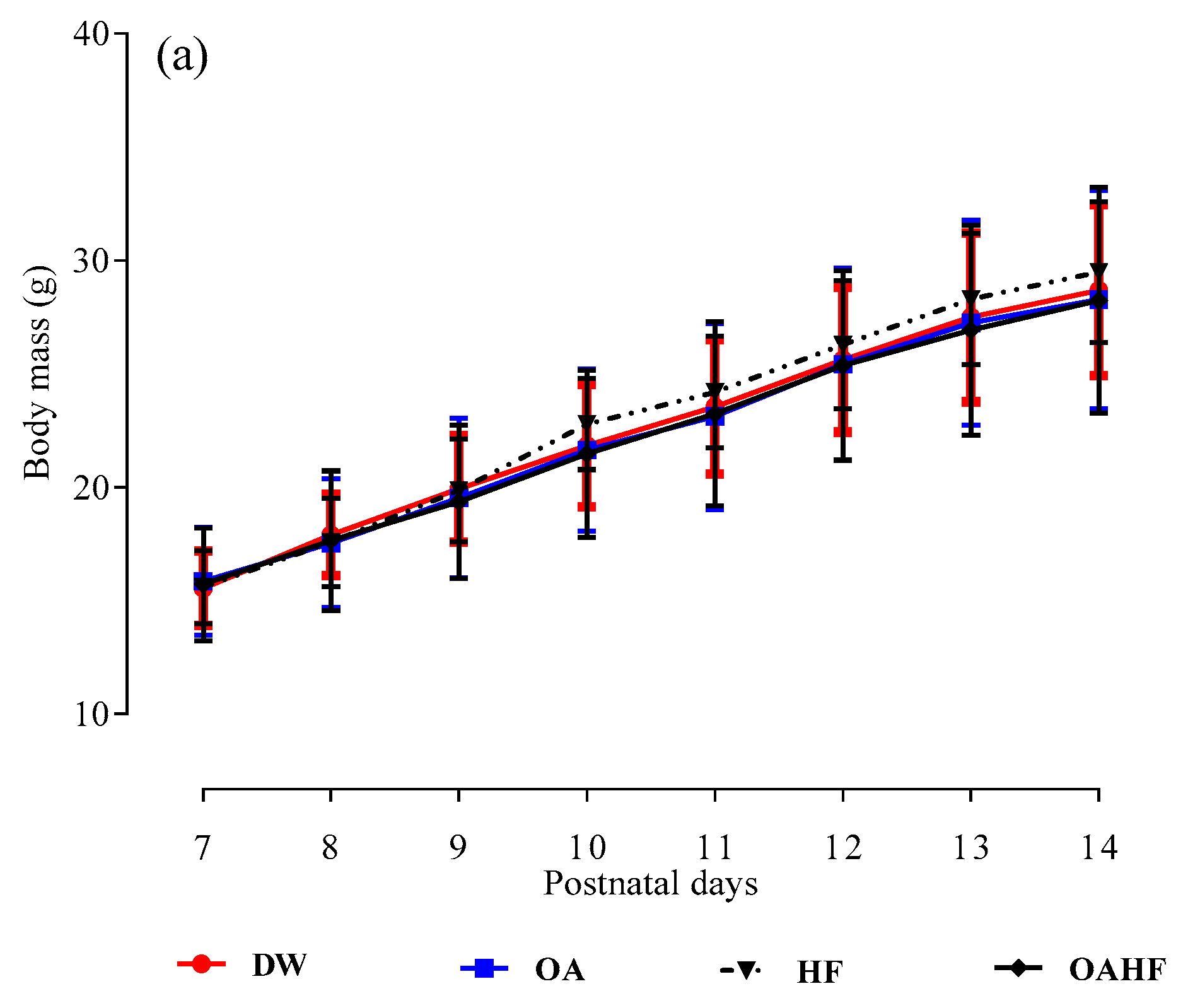
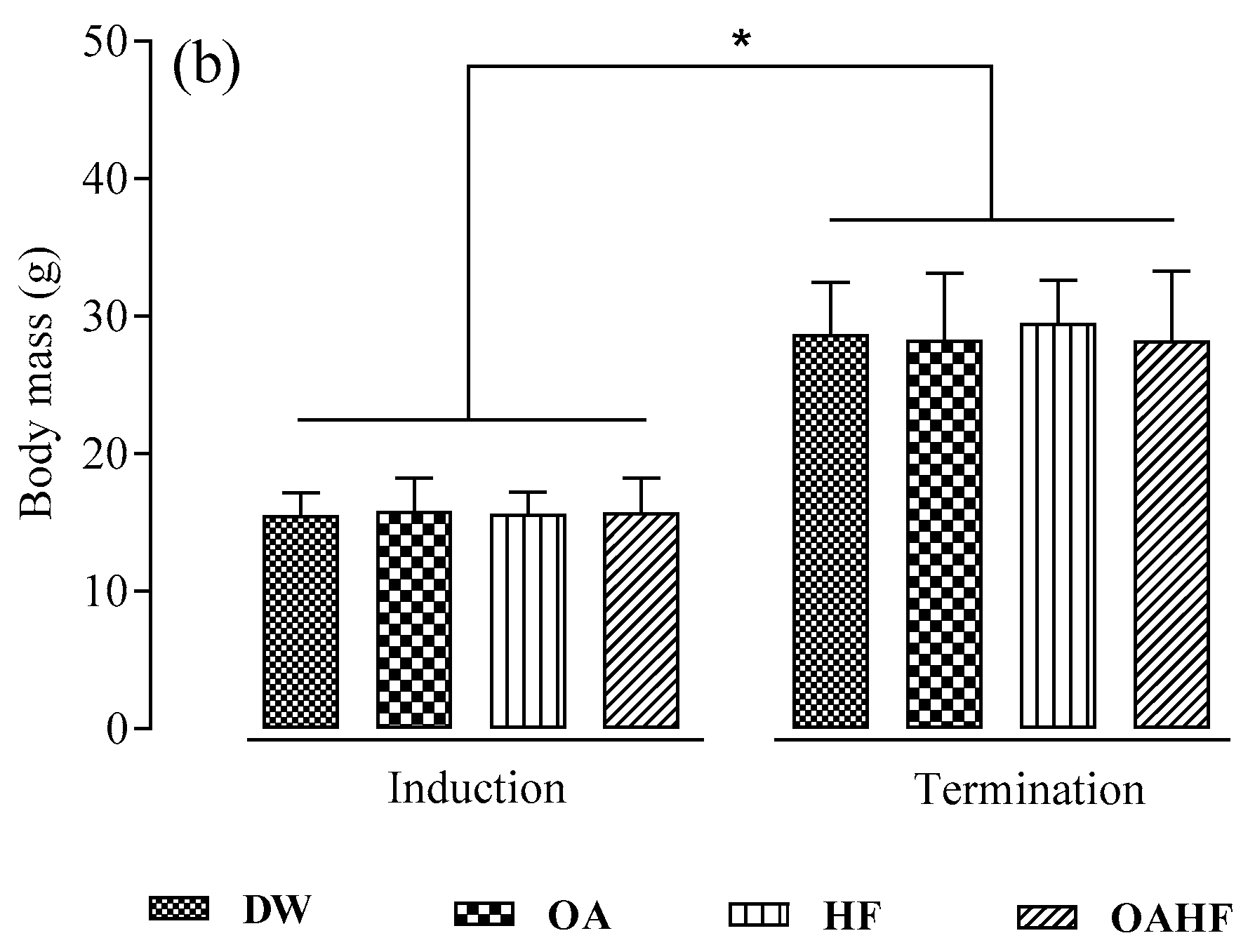
2.1. The Effect of Neonatal Oral Administration of Oleanolic Acid on Growth Performance in Suckling Male and Female Rats
2.2. Effect of Neonatal Oral Administration of Oleanolic Acid on the Gastrointestinal Tract (GIT) and Viscera Organ Morphometry in Suckling Male and Female Pups
2.3. The Effect of Neonatal Oral Administration of Oleanolic Acid on the General Clinical Health Profiles in Suckling Male and Female Pups
2.4. The Effect of Neonatal Oral Administration of Oleanolic Acid on Anti-Oxidant Enzyme Activity in the Skeletal Muscles of Suckling Male and Female Pups
2.5. The Effect of Neonatal Oral Administration of Oleanolic Acid on Antioxidant Capacity in the Skeletal Muscles of Suckling Male and Female Pups
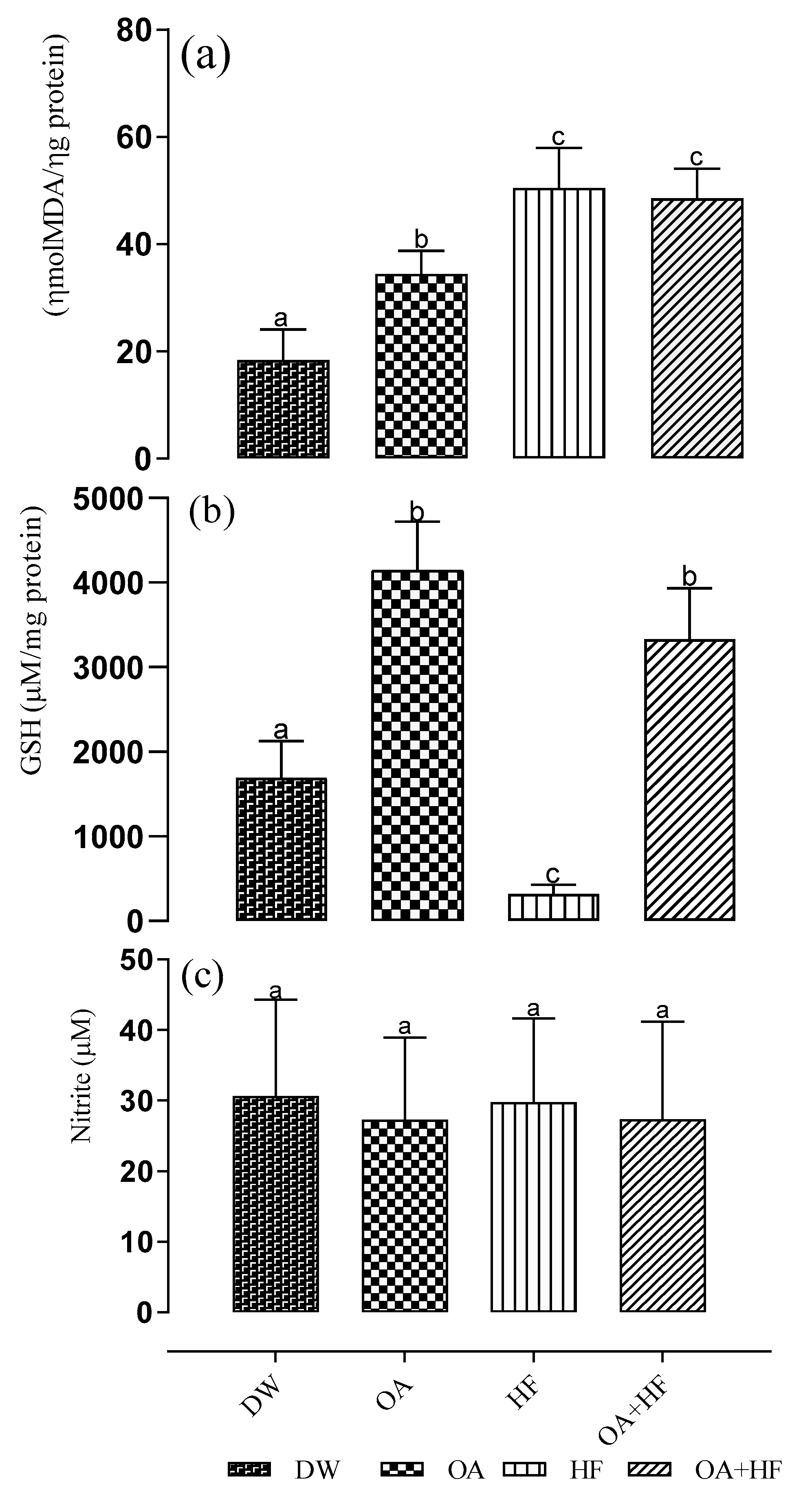
2.6. The Effect of Neonatal Oral Administration of Oleanolic Acid on Oxidative Stress Biomarkers in the Skeletal Muscles of Suckling Male and Female Pups
3. Discussion
3.1. The Effect of Neonatal Oral Administration of Oleanolic Acid on Growth Performance in Suckling Male and Female Pups
3.2. The Effect of Neonatal Oral Administration of Oleanolic Acid on the Gross Morphometry of the Abdominal Viscera in Male and Female Suckling Pups
3.3. The Effect of Neonatal Oral Administration of Oleanolic Acid on the General Clinical Health Profiles in Suckling Male and Female Pups
3.3.1. Surrogate Markers of Liver Function
3.3.2. Renal Function and General Clinical Biochemistry
3.3.3. Hepatic Lipid Storage and Biomarkers of Metabolic Function
3.4. The Effect of Neonatal Oral Administration of Oleanolic Acid on the Activities of Anti-Oxidant Enzymes and Oxidative Stress Biomarkers in the Skeletal Muscles of Suckling Male and Female Pups
4. Materials and Methods
4.1. Ethical Clearance and Study Site
4.2. Experimental Animals and Housing
4.3. Study Design and Dietary Treatments
4.4. Terminal Procedures
4.4.1. Sample Collection
4.4.2. Determination of Visceral Organ Morphometry
4.5. Measurement of Growth Performance
4.5.1. Body Mass Determination
4.5.2. Determination of Indices of Linear Bone Growth
4.6. Determination of Biochemical Health Profile Markers
4.7. Determination of Hepatic Lipid Content
4.8. Determination of the Anti-Oxidant Enzyme Activity
4.8.1. Catalase Assay
4.8.2. Superoxide Dismutase Assay
4.8.3. Glutathione Peroxidase and Glutathione Assay
4.9. Determination of Anti-Oxidant Capacity Assay
4.9.1. Ferric Reducing Anti-Oxidant Power (FRAP)
4.9.2. Trolox Equivalent Anti-Oxidant Capacity (TEAC)
4.10. Determination of Oxidative Stress Biomarkers
4.10.1. Lipid Peroxidation Assay
4.10.2. Nitrite Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khitan, Z.; Kim, D.H. Fructose: A key factor in the development of metabolic syndrome and hypertension. J. Nutr. Metab. 2013, 2013, 682673. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, S.; O’Driscoll, L. Metabolic syndrome: A closer look at the growing epidemic and its associated pathologies. Obes. Rev. 2015, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [PubMed]
- Maarman, G.; Mendham, A.; Madlala, H.; Ojuka, E. Excessive consumption of fructose-containing sugars: An emerging threat for developing nations? J. Afr. Assoc. Physiol. Sci. 2016, 4, 71–80. [Google Scholar]
- Tappy, L. Health Implications of Fructose Consumption in Humans. In Sweeteners: Pharmacology, Biotechnology, and Applications; Springer: Berlin, Germany, 2017; pp. 1–26. [Google Scholar]
- Stanhope, K.L. Sugar consumption, metabolic disease and obesity: The state of the controversy. Crit. Rev. Clin. Lab. Sci. 2016, 53, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Cigliano, L.; Spagnuolo, M.S.; Crescenzo, R.; Cancelliere, R.; Iannotta, L.; Mazzoli, A.; Liverini, G.; Iossa, S. Short-Term Fructose Feeding Induces Inflammation and Oxidative Stress in the Hippocampus of Young and Adult Rats. Mol. Neurobiol. 2017, 55, 2869–2883. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2017, 114, 1752–1761. [Google Scholar] [CrossRef]
- Higdon, J.V.; Frei, B. Obesity and oxidative stress. Am. Heart Assoc. 2003, 23, 365–367. [Google Scholar] [CrossRef]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Sankhla, M.; Sharma, T.K.; Mathur, K.; Rathor, J.S.; Butolia, V.; Gadhok, A.K.; Vardey, S.K.; Sinha, M.; Kaushik, G. Relationship of oxidative stress with obesity and its role in obesity induced metabolic syndrome. Clin. Lab. 2012, 58, 385–392. [Google Scholar]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [PubMed]
- Savini, I.; Catani, M.V.; Evangelista, D.; Gasperi, V.; Avigliano, L. Obesity-associated oxidative stress: Strategies finalized to improve redox state. Int. J. Mol. Sci. 2013, 14, 10497–10538. [Google Scholar] [CrossRef] [PubMed]
- Sikora, E.; Scapagnini, G.; Barbagallo, M. Curcumin, inflammation, ageing and age-related diseases. Immun. Ageing 2010, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.-L.; Kunsch, C. Induction of cytoprotective genes through Nrf2/antioxidant response element pathway: A new therapeutic approach for the treatment of inflammatory diseases. Curr. Pharm. Des. 2004, 10, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Pall, M.L.; Levine, S. Nrf2, a master regulator of detoxification and also antioxidant, anti-inflammatory and other cytoprotective mechanisms, is raised by health promoting factors. Acta Physiol. Sin. 2015, 67, 1–18. [Google Scholar]
- Clancy, B.; Darlington, R.; Finlay, B. Translating developmental time across mammalian species. Neuroscience 2001, 105, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Quinn, R. Comparing rat’s to human’s age: How old is my rat in people years? Nutrition 2005, 21, 775–777. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar]
- Vickers, M.H. Developmental programming of the metabolic syndrome-critical windows for intervention. World J. Diabetes 2011, 2, 137–148. [Google Scholar] [CrossRef]
- Lucas, A. Programming by early nutrition: An experimental approach. J. Nutr. 1998, 128, 401S–406S. [Google Scholar] [CrossRef]
- McArdle, H.; Andersen, H.; Jones, H.; Gambling, L. Fetal programming: Causes and consequences as revealed by studies of dietary manipulation in rats–a review. Placenta 2006, 27, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Pico, C.; Oliver, P.; Sanchez, J.; Miralles, O.; Caimari, A.; Priego, T.; Palou, A. The intake of physiological doses of leptin during lactation in rats prevents obesity in later life. Int. J. Obes. 2007, 31, 1199–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gluckman, P.D.; Buklijas, T.; Hanson, M.A. The Developmental Origins of Health and Disease (DOHaD) Concept: Past, Present, and Future. In The Epigenome and Developmental Origins of Health and Disease; Academic Press: London, UK, 2015; pp. 1–13. [Google Scholar]
- Wang, X.-M. Early life programming and metabolic syndrome. World J. Pediatr. 2013, 9, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Date, Y.; Kojima, M.; Hosoda, H.; Sawaguchi, A.; Mondal, M.S.; Suganuma, T.; Matsukura, S.; Kangawa, K.; Nakazato, M. Ghrelin, a Novel Growth Hormone-Releasing Acylated Peptide, Is Synthesized in a Distinct Endocrine Cell Type in the Gastrointestinal Tracts of Rats and Humans** This work was supported in part by grants-in-aid from the Ministry of Education, Science, Sports, and Culture, Japan, and the Ministry of Health and Welfare, Japan (to MN). Endocrinology 2000, 141, 4255–4261. [Google Scholar] [PubMed]
- Ahlman, H.; Nilsson, O. The gut as the largest endocrine organ in the body. Ann. Oncol. 2001, 12, S63–S68. [Google Scholar] [CrossRef] [PubMed]
- Beya, W.; Davidson, B.; Erlwanger, K. The effects of crude aqueous and alcohol extracts of Aloe vera on growth and abdominal viscera of suckling rats. Afr. J. Tradit. Complem. Altern. Med. 2012, 9, 553–560. [Google Scholar] [CrossRef]
- Dik, B. Treatment of metabolic syndrome. Atat. Univ. Vet. Bil. Derg. 2013, 8, 259–269. [Google Scholar]
- Lane, G. Editorial: The Role of Neutraceuticals in the Prevention & Treatment of Metabolic Syndrome. J. Nutr. Therap. 2016, 4, 113–114. [Google Scholar]
- Wang, X.; Chen, Y.; Abdelkader, D.; Hassan, W.; Sun, H.; Liu, J. Combination therapy with oleanolic acid and metformin as a synergistic treatment for diabetes. J. Diabetes Res. 2015, 2015, 973287. [Google Scholar]
- Castellano, J.; Guinda, A.; Macías, L.; Santos-Lozano, J.; Lapetra, J.; Rada, M. Free radical scavenging and α-glucosidase inhibition, two potential mechanisms involved in the anti-diabetic activity of oleanolic acid. Grasas y Aceites 2016, 67, 142. [Google Scholar] [CrossRef]
- Gutiérrez, R.M.P. Hypolipidemic and hypoglycemic activities of a oleanolic acid derivative from Malva parviflora on streptozotocin-induced diabetic mice. Arch. Pharm. Res. 2017, 40, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Khathi, A.; Mbatha, B.; Musabayane, C.T. The hypoglycaemic and antioxidant properties of oleanolic acid ameliorate blood pressure and kidney function of experimental animals. Endocr. Abs. 2015, 38, 224. [Google Scholar] [CrossRef]
- Tsao, S.-m.; Yin, M.-c. Antioxidative and antiinflammatory activities of asiatic acid, glycyrrhizic acid, and oleanolic acid in human bronchial epithelial cells. J. Agric. Food Chem. 2015, 63, 3196–3204. [Google Scholar] [CrossRef]
- Jäger, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic triterpene distribution in various plants–rich sources for a new group of multi-potent plant extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef] [PubMed]
- Camer, D.; Yu, Y.; Szabo, A.; Huang, X.F. The molecular mechanisms underpinning the therapeutic properties of oleanolic acid, its isomer and derivatives for type 2 diabetes and associated complications. Mol. Nutr. Food Res. 2014, 58, 1750–1759. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Quesada, C.; López-Biedma, A.; Gaforio, J.J. Oleanolic Acid, a Compound Present in Grapes and Olives, Protects against Genotoxicity in Human Mammary Epithelial Cells. Molecules 2015, 20, 13670–13688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef] [PubMed]
- Castellano, J.M.; Guinda, A.; Delgado, T.; Rada, M.; Cayuela, J.A. Biochemical basis of the antidiabetic activity of oleanolic acid and related pentacyclic triterpenes. Diabetes 2013, 62, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Pharmacology of oleanolic acid and ursolic acid. J. Ethnopharmacol. 1995, 49, 57–68. [Google Scholar] [CrossRef]
- Wang, X.; Ye, X.-l.; Liu, R.; Chen, H.-L.; Bai, H.; Liang, X.; Zhang, X.-D.; Wang, Z.; Li, W.-l.; Hai, C.-X. Antioxidant activities of oleanolic acid in vitro: Possible role of Nrf2 and MAP kinases. Chem. Biol. Interact. 2010, 184, 328–337. [Google Scholar] [CrossRef]
- Chen, B.; Lu, Y.; Chen, Y.; Cheng, J. The role of Nrf2 in oxidative stress-induced endothelial injuries. J. Endocrinol. 2015, 225, R83–R99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benevenga, N.J.; Calvert, C.; Eckhert, C.D.; Fahey, G.C.; Greger, J.L.; Keen, C.; Knapka, J.; Magalhaes, H.; Oftedal, O. Nutrient requirements of laboratory animals. Nutr. Requirements Gerbil 1995, 140–143. [Google Scholar]
- Maltin, C.; Delday, M.; Reeds, P. The effect of a growth promoting drug, clenbuterol, on fibre frequency and area in hind limb muscles from young male rats. Biosci. Rep. 1986, 6, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Hack, M.; Flannery, D.J.; Schluchter, M.; Cartar, L.; Borawski, E.; Klein, N. Outcomes in young adulthood for very-low-birth-weight infants. N. Engl. J. Med. 2002, 346, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Cole, T.J.; Fewtrell, M.; Deanfield, J.; Lucas, A. Is slower early growth beneficial for long-term cardiovascular health? Circulation 2004, 109, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.; Godfrey, K.M.; Gluckman, P.D.; Harding, J.E.; Owens, J.A.; Robinson, J.S. Fetal nutrition and cardiovascular disease in adult life. Lancet 1993, 341, 938–941. [Google Scholar] [CrossRef]
- Curhan, G.C.; Willett, W.C.; Rimm, E.B.; Spiegelman, D.; Ascherio, A.L.; Stampfer, M.J. Birth weight and adult hypertension, diabetes mellitus, and obesity in US men. Circulation 1996, 94, 3246–3250. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Spurgeon, H.A.; Rakusan, K.; Weisfeldt, M.L.; Lakatta, E.G. Use of tibial length to quantify cardiac hypertrophy: Application in the aging rat. Am. J. Physiol. Heart Circ. Physiol. 1982, 243, H941–H947. [Google Scholar] [CrossRef]
- Sundström, K.; Cedervall, T.; Ohlsson, C.; Camacho-Hübner, C.; Sävendahl, L. Combined treatment with GH and IGF-I: Additive effect on cortical bone mass but not on linear bone growth in female rats. Endocrinology 2014, 155, 4798–4807. [Google Scholar] [CrossRef]
- Yakar, S.; Rosen, C.J.; Beamer, W.G.; Ackert-Bicknell, C.L.; Wu, Y.; Liu, J.-L.; Ooi, G.T.; Setser, J.; Frystyk, J.; Boisclair, Y.R. Circulating levels of IGF-1 directly regulate bone growth and density. J. Clin. Investig. 2002, 110, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Guilloteau, P.; Zabielski, R.; Hammon, H.M.; Metges, C.C. Nutritional programming of gastrointestinal tract development. Is the pig a good model for man? Nutr. Res. Rev. 2010, 23, 4–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, S.; Bouley, C.; Boutron, M.-C.; Cummings, J.; Franck, A.; Gibson, G.; Isolauri, E.; Moreau, M.-C.; Roberfroid, M.; Rowland, I. Functional food science and gastrointestinal physiology and function. Br. J. Nutr. 1998, 80, S147–S171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henning, S.J. Functional development of the gastrointestinal tract. Physiol. Gastrointest. Tract 1987, 1, 285–300. [Google Scholar]
- Downes, N.; Burns, L. Juvenile toxicity: A study of histological organ development in juvenile rats and correlation with known stage of development in man. Reprod. Toxicol. 2008, 26, 60. [Google Scholar] [CrossRef]
- Jean, M. Physiology of the gastro-intestinal tract: Regulation of function and metabolism. J. Dairy Sci. 1993, 76, 2080–2093. [Google Scholar]
- Botermans, J.; Pierzynowski, S. Relations between body weight, feed intake, daily weight gain, and exocrine pancreatic secretion in chronically catheterized growing pigs. J. Anim. Sci. 1999, 77, 450–456. [Google Scholar] [CrossRef] [PubMed]
- McCance, K.L.; Huether, S.E. Pathophysiology: The Biologic Basis for Disease in Adults and Children. Dimen. Crit. Care Nurs. 1994, 13, 6. [Google Scholar]
- Henning, S.J. Postnatal development: Coordination of feeding, digestion, and metabolism. Am. J. Physiol. Gastrointest. Liver Physiol. 1981, 241, G199–G214. [Google Scholar] [CrossRef]
- Khan, I.Y.; Dekou, V.; Douglas, G.; Jensen, R.; Hanson, M.A.; Poston, L.; Taylor, P.D. A high-fat diet during rat pregnancy or suckling induces cardiovascular dysfunction in adult offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R127–R133. [Google Scholar] [CrossRef] [Green Version]
- Armitage, J.A.; Taylor, P.D.; Poston, L. Experimental models of developmental programming: Consequences of exposure to an energy rich diet during development. J. Physiol. 2005, 565, 3–8. [Google Scholar] [CrossRef]
- Huynh, M.; Luiken, J.J.; Coumans, W.; Bell, R.C. Dietary fructose during the suckling period increases body weight and fatty acid uptake into skeletal muscle in adult rats. Obesity 2008, 16, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- LIU, L.; Liu, Y.; Mao, Q.; KLAASSEN, C. The effects of 10 triterpenoid compounds on experimental liver injury in mice. Toxicol. Sci. 1994, 22, 34–40. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Klaassen, C.D. Protective effect of oleanolic acid against chemical-induced acute necrotic liver injury in mice. Acta Pharmacol. Sin. 1995, 16, 97–102. [Google Scholar]
- Rouiller, C. The Liver: Morphology, Biochemistry, Physiology; Academic Press: London, UK, 2013. [Google Scholar]
- Oosterveer, M.H.; Schoonjans, K. Hepatic glucose sensing and integrative pathways in the liver. Cell. Mol. Life Sci. 2014, 71, 1453–1467. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Feng, X.; Zhang, L.; Chen, S.; Cheng, Y.; He, X.; Yang, Y.; He, Y.; Wang, H.; Wang, R. Hepatic expression of detoxification enzymes is decreased in human obstructive cholestasis due to gallstone biliary obstruction. PLoS ONE 2015, 10, e0120055. [Google Scholar] [CrossRef] [PubMed]
- Leise, M.D.; Poterucha, J.J.; Talwalkar, J.A. Drug-Induced Liver Injury; Elsevier: Amsterdam, The Netherlands, 2014; pp. 95–106. [Google Scholar]
- Shu, R.; David, E.; Ferraris, R. Dietary fructose enhances intestinal fructose transport and GLUT5 expression in weaning rats. Am. J. Physiol. Gastrointest. Liver Physiol. 1997, 272, G446–G453. [Google Scholar] [CrossRef] [PubMed]
- Vella, S.J.; Beattie, P.; Cademartiri, R.; Laromaine, A.; Martinez, A.W.; Phillips, S.T.; Mirica, K.A.; Whitesides, G.M. Measuring markers of liver function using a micropatterned paper device designed for blood from a fingerstick. Anal. Chem. 2012, 84, 2883–2891. [Google Scholar] [CrossRef]
- Thapa, B.; Walia, A. Liver function tests and their interpretation. Indian J. Pediatr. 2007, 74, 663–671. [Google Scholar] [CrossRef]
- Moseley, R.H. Evaluation of abnormal liver function tests. Med. Clin. N. Am. 1996, 80, 887–906. [Google Scholar] [CrossRef]
- Scheig, R. Evaluation of tests used to screen patients with liver disorders. Prim. Care 1996, 23, 551–560. [Google Scholar] [CrossRef]
- Limdi, J.; Hyde, G. Evaluation of abnormal liver function tests. Postgrad. Med. J. 2003, 79, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heemskerk, S.; Masereeuw, R.; Moesker, O.; Bouw, M.P.; van der Hoeven, J.G.; Peters, W.H.; Russel, F.G.; Pickkers, P.; Group, A.S. Alkaline phosphatase treatment improves renal function in severe sepsis or septic shock patients. Crit. Care Med. 2009, 37, 417-e1. [Google Scholar] [CrossRef] [PubMed]
- Carvounis, C.P.; Nisar, S.; Guro-Razuman, S. Significance of the fractional excretion of urea in the differential diagnosis of acute renal failure. Kidney Int. 2002, 62, 2223–2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisco, M.A.; Coca, S.G.; Chen, J.; Owens, A.T.; McCauley, B.D.; Kimmel, S.E.; Testani, J.M. The Blood Urea Nitrogen to Creatinine Ratio Identifies a High Risk but Potentially Reversible Form of Renal Dysfunction in Patients with Decompensated Heart Failure. Circ. Heart Fail. 2013, 6, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Corey, K.E.; Kaplan, L.M. Obesity and liver disease: The epidemic of the twenty-first century. Clin. Liver Dis. 2014, 18, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, L.P.; Hannivoort, R.A.; Gerken, G.; Hotamisligil, G.S.; Trauner, M.; Canbay, A. The interaction of hepatic lipid and glucose metabolism in liver diseases. J. Hepatol. 2012, 56, 952–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar]
- Basaranoglu, M.; Basaranoglu, G.; Bugianesi, E. Carbohydrate intake and nonalcoholic fatty liver disease: Fructose as a weapon of mass destruction. Hepatobil. Surg. Nutr. 2014, 4, 109–116. [Google Scholar]
- Abdel-Kawi, S.H.; Hassanin, K.M.A.; Hashem, K.S. The effect of high dietary fructose on the kidney of adult albino rats and the role of curcumin supplementation: A biochemical and histological study. Beni-Suef Univ. J. Basic Appl. Sci. 2016, 5, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef]
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of Oxidative Stress in Inflammatory Bowel Disease and Potential Antioxidant Therapies. Oxid. Med. Cell. Longev. 2017, 2017, 4535194. [Google Scholar] [CrossRef] [PubMed]
- La Russa, D.; Brunelli, E.; Pellegrino, D. Oxidative imbalance and kidney damage in spontaneously hypertensive rats: Activation of extrinsic apoptotic pathways. Clin. Sci. 2017, 131, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Kozakowska, M.; Pietraszek-Gremplewicz, K.; Jozkowicz, A.; Dulak, J. The role of oxidative stress in skeletal muscle injury and regeneration: Focus on antioxidant enzymes. J. Muscle Res. Cell Motil. 2015, 36, 377–393. [Google Scholar] [CrossRef]
- Jarukamjorn, K.; Jearapong, N.; Pimson, C.; Chatuphonprasert, W. A high-fat, high-fructose diet induces antioxidant imbalance and increases the risk and progression of nonalcoholic fatty liver disease in mice. Scientifica 2016, 2016, 5029414. [Google Scholar] [CrossRef] [PubMed]
- Seedor, J.G.; Quartuccio, H.A.; Thompson, D.D. The Biphosphonate Aledronate (MK-217) Inhibits Bone Loss due to Ovariectomy in Rats. J. Bone Miner. Res. 2005, 20, 354–362. [Google Scholar] [CrossRef]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. [30] Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar]
- Yang, H.J.; Kim, K.Y.; Kang, P.; Lee, H.S.; Seol, G.H. Effects of Salvia sclarea on chronic immobilization stress induced endothelial dysfunction in rats. BMC Complement. Altern. Med. 2014, 14, 396. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of oleanolic acid were purchased from Sigma-Aldrich South Africa and are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Parameter | DW | OA | HF | OAHF |
|---|---|---|---|---|
| Tibia | ||||
| Mass (mg) | 38.50 ± 1.4 | 35.9 ± 2.0 | 37.6 ± 1.1 | 34 ± 2.8 |
| Length (mm) | 15.2 ± 0.7 | 15 ± 0.8 | 17.1 ± 0.9 | 14.8 ± 0.9 |
| ¥ Seedor index (mg/mm) | 2.5 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.1 | 2.30 ± 0.1 |
| Femur | ||||
| Mass (mg) | 37.8 ± 5.8 | 33.5 ± 4.5 | 37.6 ± 8.5 | 34.1 ± 5.5 |
| Length (mm) | 11.4 ± 0.9 | 11.6 ± 0.7 | 11.9 ± 1.7 | 11.1 ± 1.5 |
| Seedor index (mg/mm) | 3.3 ± 0.5 | 2.9 ± 0.3 | 3.2 ± 0.4 | 3.1 ± 0.4 |
| Parameter | DW | OA | HF | OAHF |
|---|---|---|---|---|
| SI (cm) | 54.44 ± 9.23 | 46.75 ± 16.48 | 52.24 ± 5.06 | 53.34 ± 8.59 |
| SI (g) | 0.74 ± 0.19 | 0.74 ± 0.22 | 0.84 ± 0.27 | 0.93 ± 0.32 |
| SI ¥ rTL | 0.49 ± 0.14 | 0.45 ± 0.15 | 0.49 ± 0.14 | 0.62 ± 0.19 |
| LI (cm) | 7.09 ± 0.79 | 6.63 ± 2.72 | 7.64 ± 0.56 | 7.47 ± 0.55 |
| LI (g) | 0.12 ± 0.01 | 0.13 ± 0.02 | 0.13 ± 0.02 | 0.13 ± 0.03 |
| LI rTL | 0.08 ± 0.01 | 0.08 ± 0.02 | 0.08 ± 0.00 | 0.09 ± 0.01 |
| Liver (g) | 0.90 ± 0.17 | 0.94 ± 0.17 | 0.94 ± 0.16 | 1.01 ± 0.21 |
| Liver rTL | 0.60 ± 0.13 | 0.63 ± 0.13 | 0.55 ± 0.10 | 0.68 ± 0.13 |
| Caecum (g) | 0.08 ± 0.02 | 0.06 ± 0.03 | 0.07 ± 0.01 | 0.07 ± 0.02 |
| Caecum rTL | 0.04 ± 0.02 | 0.04 ± 0.02 | 0.04 ± 0.00 | 0.05 ± 0.01 |
| Stomach (g) | 0.21 ± 0.02 | 0.21 ± 0.05 | 0.20 ± 0.03 | 0.20 ± 0.01 |
| Stomach rTL | 0.14 ± 0.02 | 0.14 ± 0.03 | 0.12 ± 0.02 | 0.13 ± 0.01 |
| Kidneys (g) | 0.38 ± 0.04 | 0.27 ± 0.04 | 0.38 ± 0.03 | 0.32 ± 0.14 |
| Kidneys rTL | 0.25 ± 0.03 | 0.27 ± 0.04 | 0.23 ± 0.03 | 0.22 ± 0.10 |
| Parameter | DW | OA | HF | OAHF |
|---|---|---|---|---|
| BUN (mmol/L) | 4.6 ± 0.7 | 4.2 ± 0.9 | 5.2 ± 0.6 | 4.3 ± 0.5 |
| CREA (µmol/L) | 16.9 ± 3.2 | 18 ± 0 | 10.3 ± 3.4 | 14.4 ± 7.1 |
| TBIL (µmol/L) | 4 ± 2.7 | 5.5 ± 3.3 | 8 ± 3.8 | 4.12 ± 2.1 |
| PHOS (mmol/L) | 2.8 ± 0.2 | 3.17 ± 0.5 | 2.9 ± 0.3 | 3.2 ± 0.3 |
| CAL (mmol/L) | 2.7 ± 0.4 | 2.0 ± 0.9 | 2.3 ± 0.6 | 2.5 ± 0.7 |
| TPROT (g/L) | 38.1 ± 4.1 | 41.3 ± 3.7 | 41.4 ± 5.2 | 40 ± 3.0 |
| ALB (g/L) | 21.1 ± 2.9 | 19.5 ± 2.4 | 20.17 ± 2.7 | 19.3 ± 1.6 |
| GLOB (g/L) | 17 ± 4.5 | 21.7 ± 2.0 | 21.3 ± 1.2 | 20.4 ± 2.9 |
| ALT (U/L) | 35 ± 5.9 | 45 ± 14.1 | 51.8 ± 15.5 | 43.9 ± 7.2 |
| ALP (U/L) | 299.1 ± 63.8 | 309.3 ± 58.5 | 394.3 ± 70.3 | 269.4 ± 62.7 |
| * Hepatic lipid content (%) | 2.8 ± 0.0 | 2.7 ± 0.0 | 3.2 ± 0.0 | 3.2 ± 0.0 |
| Glucose (mmol/L) | 7.8 ± 1.3 | 7.6 ± 1.0 | 8.1 ± 0.3 | 8.5 ± 1.7 |
| Cholesterol (mmol/L) | 4.3 ± 0.4 | 4.3 ± 0.5 | 4.3 ± 0.5 | 4.4 ± 0.8 |
| Parameter | DW | OA | HF | OAHF |
|---|---|---|---|---|
| GPx (µM/mg protein) | 308.5 ± 19.4 a | 627.5 ± 45.7 b | 555.3 ± 29.3 b | 533.2 ± 31.1 b |
| SOD activity (% tissue inhibition rate) | 78.2 ± 3.6 a | 93.4 ± 0.9 b | 96.6 ± 0.4 b | 94.1 ± 1.2 b |
| CAT activity (kU/L) in tissue | 1.1 ± 0.0 a | 1.4 ± 0.1 b | 0.5 ± 0.0 c | 1.0 ± 0.0 ad |
| DW | OA | HF | OAHF | |
|---|---|---|---|---|
| TEAC µM/mL) | 356.4 ± 13.4 a | 392.8 ± 5.79 a | 469.3 ± 11.9 b | 393.9 ± 12.4 a |
| FRAP (µM/mL) | 36.98 ± 5.24 a | 30.16 ± 2.54 a | 30.22 ± 1.68 a | 36.95 ± 4.56 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nyakudya, T.T.; Isaiah, S.; Ayeleso, A.; Ndhlala, A.R.; Mukwevho, E.; Erlwanger, K.H. Short-Term Neonatal Oral Administration of Oleanolic Acid Protects against Fructose-Induced Oxidative Stress in the Skeletal Muscles of Suckling Rats. Molecules 2019, 24, 661. https://doi.org/10.3390/molecules24040661
Nyakudya TT, Isaiah S, Ayeleso A, Ndhlala AR, Mukwevho E, Erlwanger KH. Short-Term Neonatal Oral Administration of Oleanolic Acid Protects against Fructose-Induced Oxidative Stress in the Skeletal Muscles of Suckling Rats. Molecules. 2019; 24(4):661. https://doi.org/10.3390/molecules24040661
Chicago/Turabian StyleNyakudya, Trevor Tapiwa, Simon Isaiah, Ademola Ayeleso, Ashwell Rungano Ndhlala, Emmanuel Mukwevho, and Kennedy Honey Erlwanger. 2019. "Short-Term Neonatal Oral Administration of Oleanolic Acid Protects against Fructose-Induced Oxidative Stress in the Skeletal Muscles of Suckling Rats" Molecules 24, no. 4: 661. https://doi.org/10.3390/molecules24040661
APA StyleNyakudya, T. T., Isaiah, S., Ayeleso, A., Ndhlala, A. R., Mukwevho, E., & Erlwanger, K. H. (2019). Short-Term Neonatal Oral Administration of Oleanolic Acid Protects against Fructose-Induced Oxidative Stress in the Skeletal Muscles of Suckling Rats. Molecules, 24(4), 661. https://doi.org/10.3390/molecules24040661