Isolation, Characterisation, and Lipase Production of a Cold-Adapted Bacterial Strain Pseudomonas sp. LSK25 Isolated from Signy Island, Antarctica

 ,
, 
Abstract
:1. Introduction
2. Results
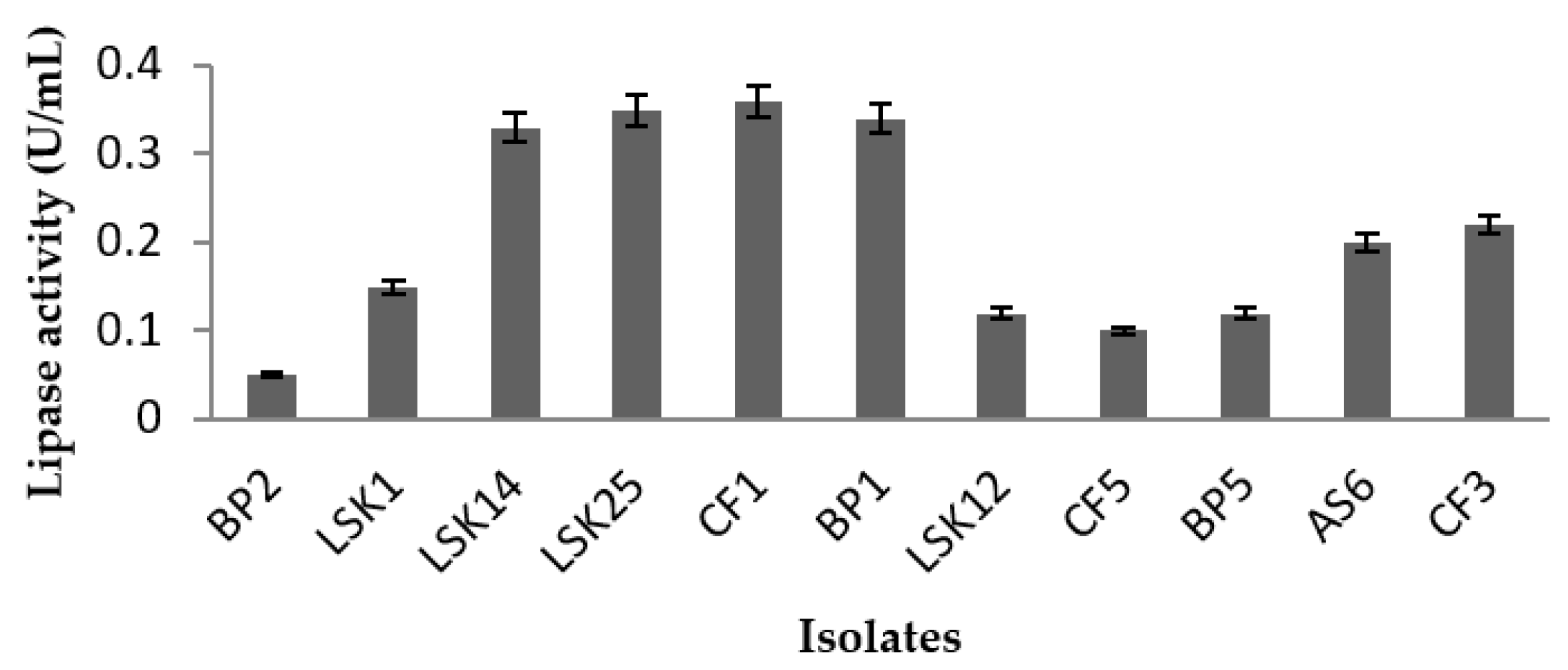
2.1. Screening for Cold Adapted Lipase
2.2. Lipase Gene Quantification via qPCR
2.3. Strain Characterisation
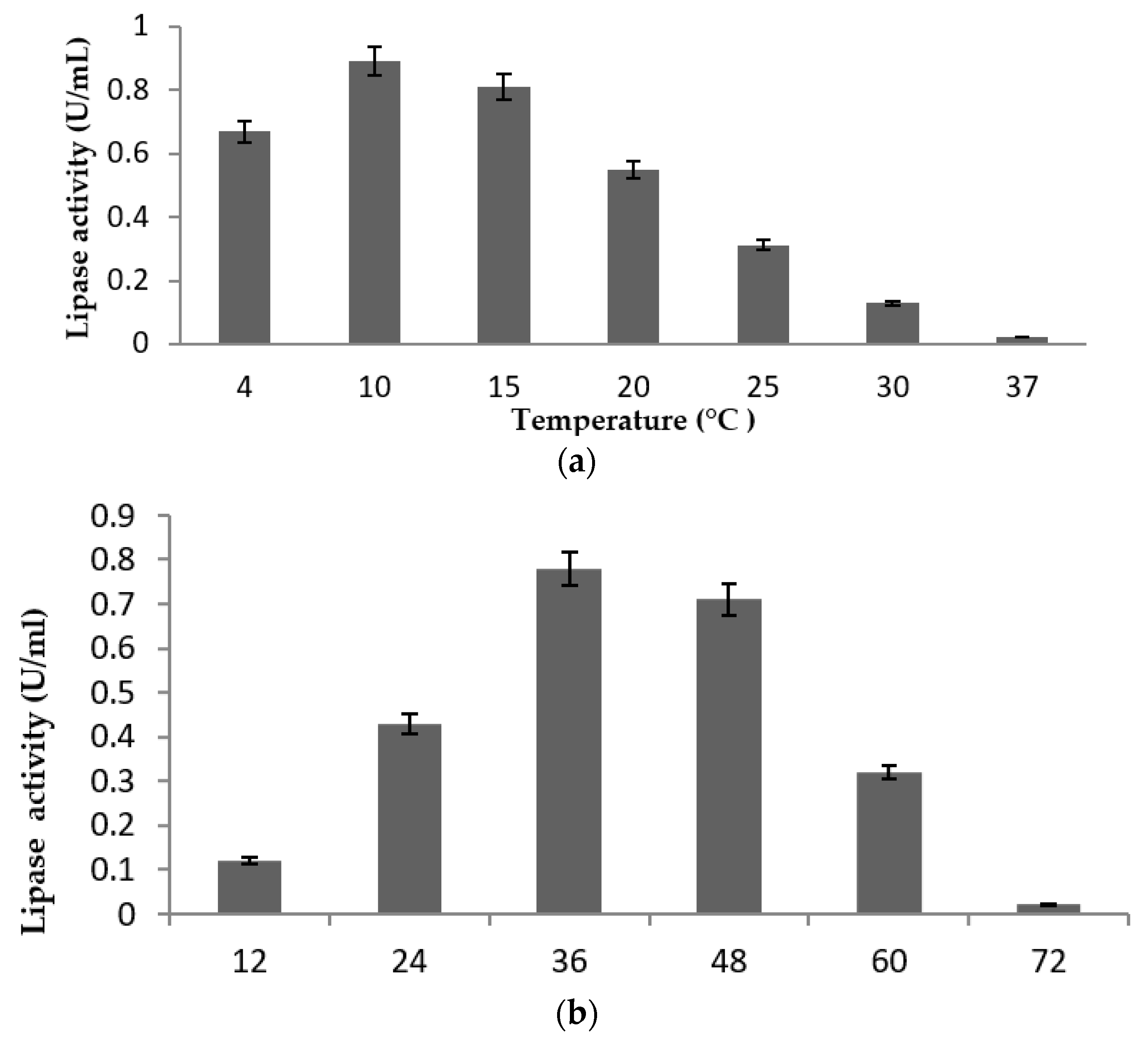
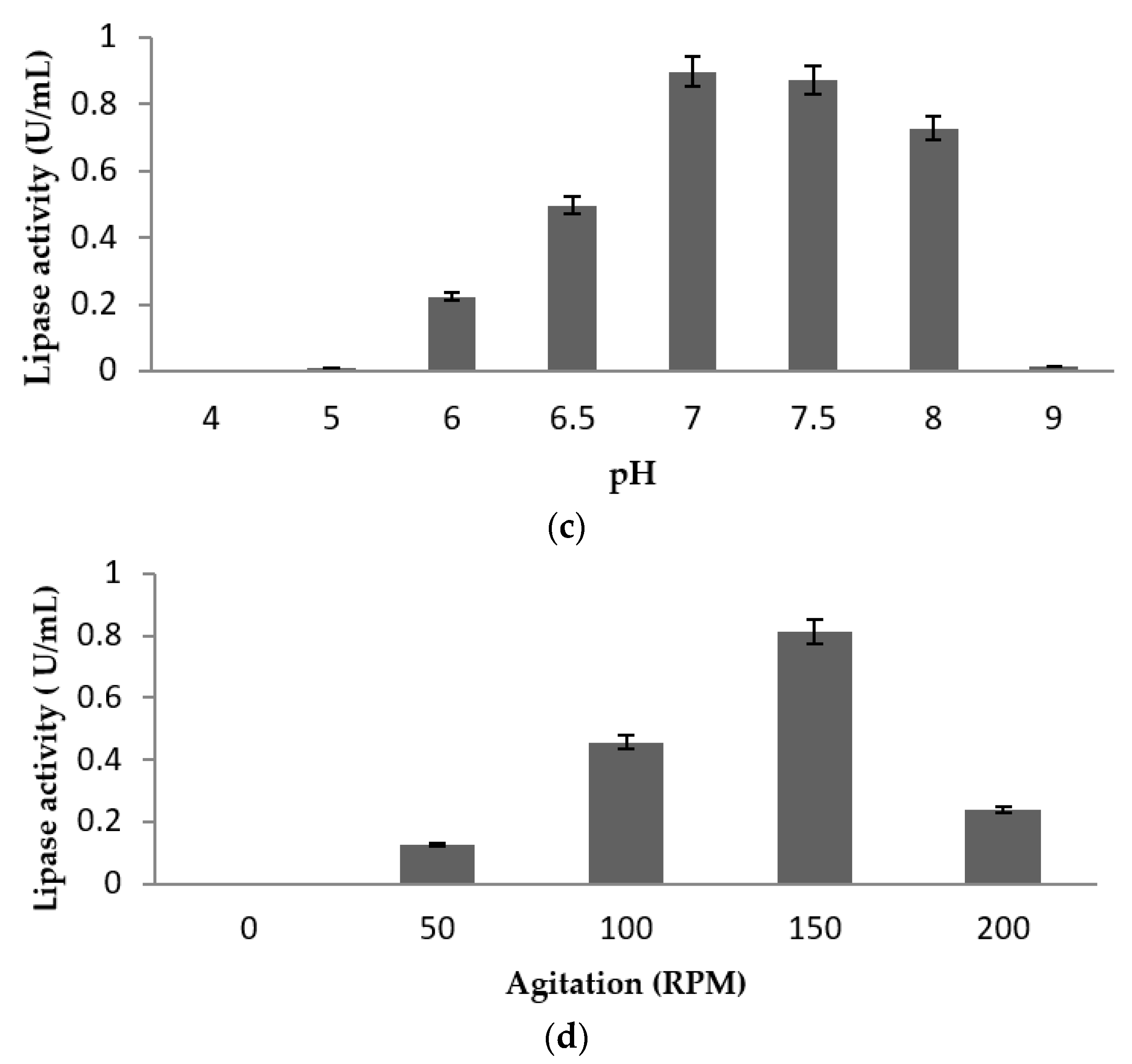
2.4. Effect of Physical Parameters on Lipase Production
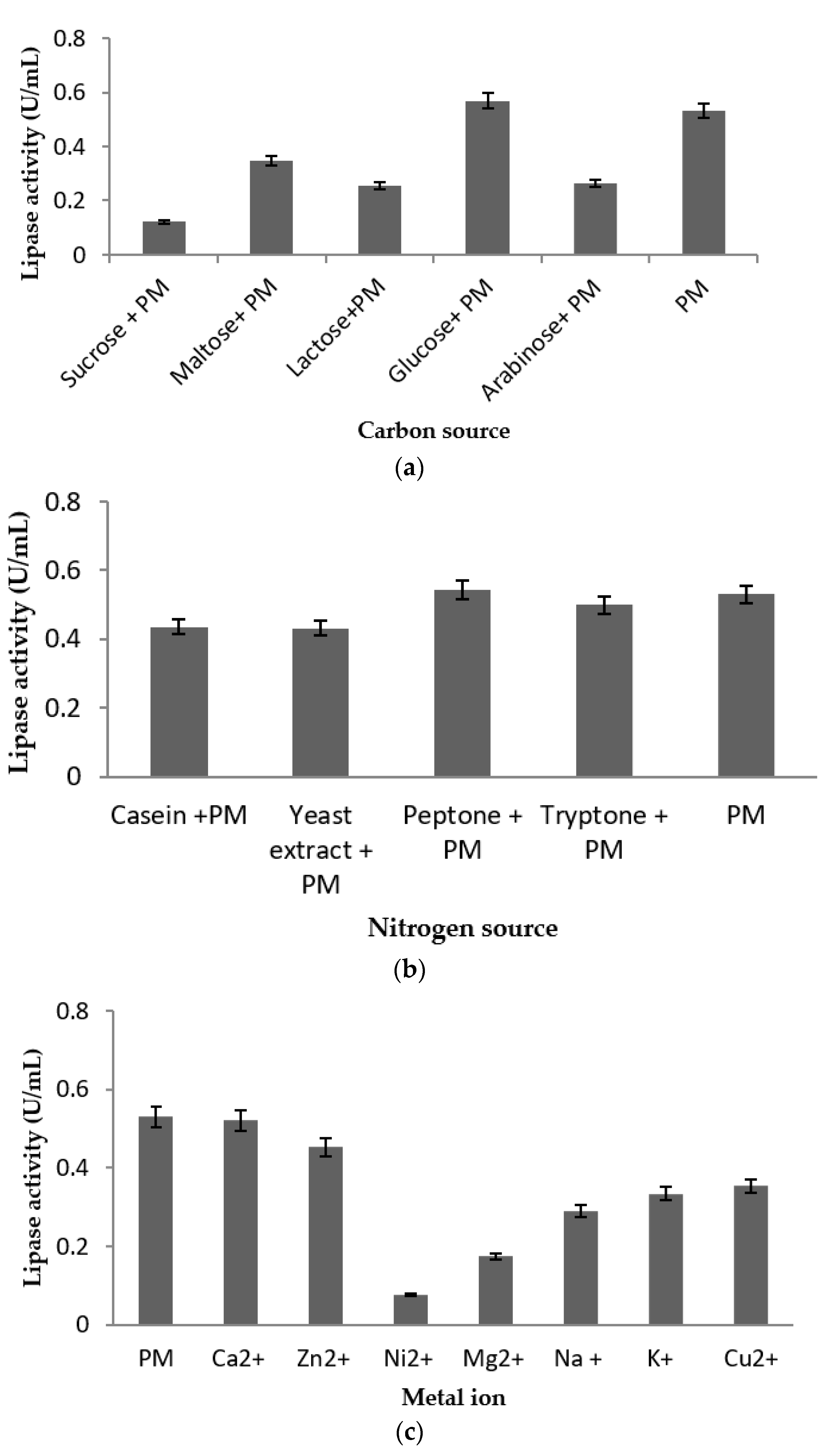
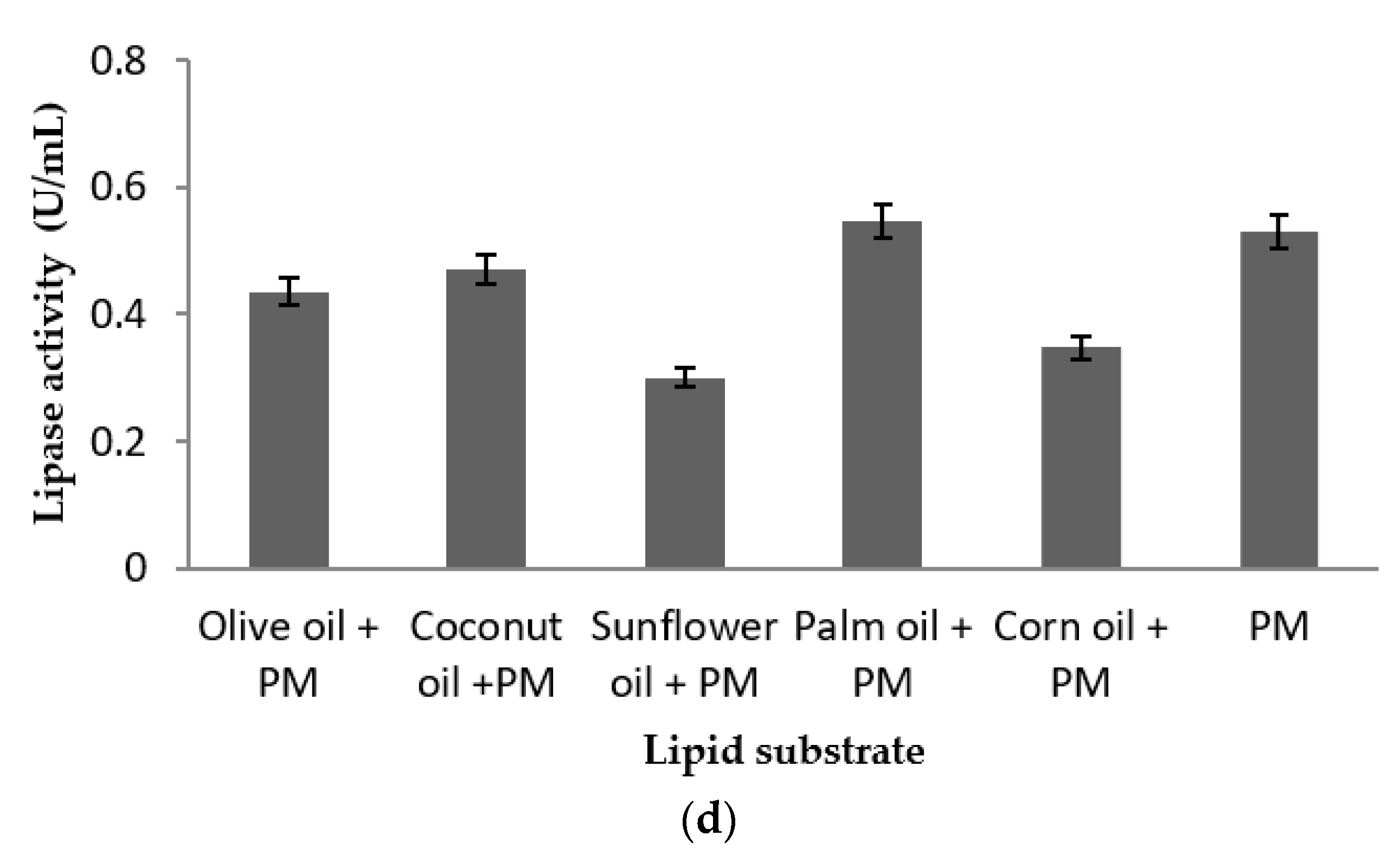
2.5. Effect of Nutritional Parameters on Lipase Production
3. Discussion
4. Materials and Methods
4.1. Study Sites
4.2. Soil Sampling
4.3. Bacterial Culture
4.4. Screening of Cold-Adapted Lipase
4.4.1. Qualitative Analyses
4.4.2. Quantitative Analyses
4.4.3. Characterisation of the Strains
4.5. Absolute Quantification (qPCR)
4.6. Effect of Physical and Nutritional Factors on Bacterial Growth and Lipase Production
4.6.1. Effect of Temperature, Incubation Time pH and Agitation
4.6.2. Effect of Nutritional Factors on Bacterial Growth and Lipase Production
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gerday, C.; Aittaleb, M.; Bentahir, M.; Chessa, J.-P.; Claverie, P.; Collins, T.; D’Amico, S.; Dumont, J.; Garsoux, G.; Georlette, D.; et al. Cold-adapted enzymes: From fundamentals to biotechnology. Trends Biotechnol. 2000, 18, 103–107. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Charlton, T.; Ertan, H.; Mohd Omar, S.; Siddiqui, K.S.; Williams, T.J. Biotechnological uses of enzymes from psychrophiles. Microb. Biotechnol. 2011, 4, 449–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, L.L.; Hou, Y.J.; Fan, H.X.; Qu, J.; Qi, C.; Liu, Y.; Li, D.F.; Liu, Z.P. Molecular structural basis for the cold adaptedness of the psychrophilic β- glucosidase BglU in Micrococcus antarcticus. Appl. Environ. Microbiol. 2016, 82, 2021–2030. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, M.; Shanthi, C. Isolation and characterization of cold active lipase producing Pseudomonas sp. 4 from marine samples of Tamilnadu Coast. Res. J. Biotech. 2013, 8, 57–62. [Google Scholar]
- Feller, G.; Gerday, C. Psychrophilic enzymes: hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.N; Dieckmann, G.S. Antarctic sea ice–a habitat for extremophiles. Science 2002, 80, 641–644. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S; Cavicchioli, R. Cold-adapted enzymes. Annu. Rev. Biochem. 2006, 75, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S; Williams, T.J; Wilkins, D. Psychrophiles. Annu. Rev. Earth Planet Sci. 2013, 41, 87–115. [Google Scholar] [CrossRef]
- Choo, D.; Kurihara, T.; Suzuki, T.; Soda, K.A. Cold-adapted lipase of an Alaskan gene cloning and enzyme purification and characterization. Appl. Environ. Microbiol. 1998, 64, 1–7. [Google Scholar]
- Feller, G.; Narinx, E.; Arpigny, J.L.; Aittaleb, M.; Baise, E.; Genicot, S.; Gerday, C. Enzymes from psychrophilic organisms. FEMS Microbiol. Rev. 1996, 18, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Joseph, B.; Ramteke, R.W.; Thomas, G. Cold active microbial lipases: some hot issues and recent developments. Biotechnol. Adv. 2008, 26, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Afsar, A.; Cetinkaya, F. A research on increasing the effectiveness of degreasing process by using enzymes. Tekstil Ve Konfeksiyon. 2008, 18, 278–283. [Google Scholar]
- Suzuki, T.; Nakayama, T.; Kurihara, T.; Nishino, T.; Esaki, N. Cold-active lipolytic activity of psychrotrophic Acinetobacter sp. strain no. 6. J. Biosci. Bioeng. 2001, 92, 144–148. [Google Scholar] [CrossRef]
- Javed, S.; Azeem, F.; Hussain, S.; Rasul, I.; Siddique, M.H.; Riaz, M.; Afzal, M.; Kouser, A.; Nadeem, H. Bacterial lipases: A review on purification and characterization. Prog. Biophys. Mol. Biol. 2018, 132, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Kuddus, M. Cold-active enzymes in food biotechnology: An updated mini review. J. App. Biol. Biotech. 2018, 6, 58–63. [Google Scholar] [CrossRef]
- Rahman, R.; Masomian, M.; Salleh, A.; Basri, M. A new thermostable lipase by Aneurinibacillus thermoaerophilus strain HZ: Nutritional studies. Ann. Microbiol. 2009, 59, 133–139. [Google Scholar] [CrossRef]
- Masomian, M.; Rahman, R.N.Z.R.A.; Salleh, A.B.; Basri, M. A unique thermostable and organic solvent tolerant lipase from newly isolated Aneurinibacillus thermoaerophilus strain HZ: Physical factor studies. World J. Microbiol. Biotechnol. 2010, 26, 1693–1701. [Google Scholar] [CrossRef]
- Kim, Y.O.; Khosasih, V.; Nam, B.H.; Lee, S.J.; Suwanto, A.; Kim, H.K. Gene cloning and catalytic characterization of cold-adapted lipase of Photobacterium sp. MA1-3 isolated from blood clam. J. Biosci. Bioeng. 2011, 114, 589–595. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Rhee, J.S. A simple and rapid colorimetric method for determination of free fatty acids for lipase assay. J. Am. Oil Chem.’ Soc. 1986, 63, 89–92. [Google Scholar] [CrossRef]
- Ohgiya, S.; Hoshino, T.; Okuyama, H.; Tanaka, S.; Ishizaki, K. Biotechnology of enzymes from cold adapted microorganisms. In Biotechnological Applications of Cold Adapted Organisms; Margesin, R., Schinner, F., Eds.; Springer: Berlin, Germany, 1999; pp. 17–34. [Google Scholar]
- Marchi, P.; Longhi, V.; Zangrossi, S.; Gaetani, E.; Briani, F.; Dehò, G. Autogenous regulation of Escherichia coli polynucleotide phosphorylase during cold acclimation by transcription termination and antitermination. Mol. Genet. Genom. 2007, 278, 75–84. [Google Scholar] [CrossRef]
- Kavitha, M. Cold active lipases—An update. Front. Life Sci. 2016, 9, 226–238. [Google Scholar] [CrossRef]
- Baharum, S.N.; Salleh, A.B.; Razak, C.N.A.; Basri, M.; Rahman, M.B.A.; Rahman, R.N.Z.R.A. Organic solvent tolerant lipase by Pseudomonas sp. strain S5: stability of enzyme in organic solvent and physical factors affecting its production. Ann. Microb. 2003, 53, 75–83. [Google Scholar]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial lipases: An overview of production, purification and biochemical properties. Appl. Microbiol. Biotechnol. 2004, 64, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.S.N. Psychrophilic pseudomonads from Antarctica: Pseudomonas antarctica sp. nov., Pseudomonas meridiana sp. nov. and Pseudomonas proteolytica sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Rashid, N.; Shimada, Y.; Ezaki, S.; Atomi, H.; Imanaka, T. Low-temperature lipase from psychrotrophic Pseudomonas sp. strain KB700A. Appl. Environ. Microbiol. 2001, 67, 4064–4069. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Ahn, M.J.; Kwak, S.H.; Song, W.H.; Jeong, B.C. Purification and characterization of cold active lipase from psychrotrophic Aeromonas sp. LPB 4. J. Microbiol. 2003, 41, 22–27. [Google Scholar]
- Rahman, R.N.Z.A.; Geok, L.P.; Basri, M.; Salleh, A.B. Physical factors affecting the production of organic solvent-tolerant protease by Pseudomonas aeroginosa strain K. Bioresource Technol. 2005, 96, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Parulekar, S.J. A parametric study of protease production in batch and fed-batch culture of Bacillus firmus. Biotechnol. Bioeng. 1991, 37, 467–483. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Joseph, B. Production of extracellular cold active lipase by Curtobacterium sp. using cell immobilization. Int. J. Genet. Eng. Biotech. 2011, 2, 33–46. [Google Scholar]
- Geoffry, K.; Achur, R.N. Screening and production of lipase from fungal organisms. Biocatal. Agric. Biotechnol. 2018, 14, 241–253. [Google Scholar] [CrossRef]
- Joseph, B.; Shrivastava, N.; Ramteke, P.W. Extracellular cold-active lipase of Microbacterium luteolum isolated from Gangotri glacier, western Himalaya: Isolation, partial purification and characterization. J. Genet. Eng. Biotechnol. 2012, 10, 137–144. [Google Scholar] [CrossRef]
- Jacques, R. Culture media. In Industrial Application of Microbiology; Moss, M.O., Smith, J.E., Eds.; Surrey University Press: London, UK, 1977; pp. 27–58. [Google Scholar]
- Boonchaidung, T.; Papone, T.; Leesing, R. Effect of carbon and nitrogen sources on lipase production by isolated lipase-producing soil yeast. J. Life Sci. Technol. 2013, 1, 176–179. [Google Scholar] [CrossRef]
- Teng, Y.; Xu, Y. Culture condition improvement for whole-cell lipase production in submerged fermentation by Rhizopus chinensis using statistical method. Bioresour. Technol. 2008, 99, 3900–3907. [Google Scholar] [CrossRef]
- Peberdy, J.F. Protein secretion in filamentous fungi—trying to understand a highly productive black box. Trends Biotechnol. 1994, 12, 50–57. [Google Scholar] [CrossRef]
- Borkar, P.S.; Bodade, R.G.; Rao, S.R.; Khobragade, C.N. Purification and characterization of extracellular lipase from a new strain: Pseudomonas aeruginosa SRT 9. Braz. J. Microbiol. 2009, 40, 358–366. [Google Scholar] [CrossRef]
- Singh, P.; Singh, S.M.; Tsuji, M.; Prasad, G.S.; Hoshino, T. Rhodotorula svalbardensis sp. nov., a novel yeast species isolated from cryoconite holes of Ny-Ålesund, Arctic. Cryobiology 2014, 68, 122–128. [Google Scholar] [CrossRef]
- Zouaoui, B.; Bouziane, A. Isolation, optimisation and purification of lipase production by Pseudomonas aeruginosa. J. Biotechnol. Biomater. 2011, 1, 10–13. [Google Scholar] [CrossRef]
- Lee, P.G. Production, purification, characterization and expression of the organic solvent tolerant protease gene. Ph.D. Thesis, Universiti Putra Malaysia, Selangor, Malaysia, 2002. [Google Scholar]
- Russi, A.E.; Brown, M.A. HHS Public Access. 2016, 165, 255–269. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Siddiqui, K.S. Cold adapted enzymes. In Enzyme Technology; Pandey, A., Webb, C., Soccol, C.R., Larroche, C., Eds.; Asiatech Publishers: New Delhi, India, 2004; pp. 615–638. [Google Scholar]
- Alford, J.A.; Pierce, D.A. Lipolytic activity of microorganisms at low and intermediate temperatures. Activity of microbial lipases at temperatures below 0 °C. J. Food Sci. 1961, 26, 518–524. [Google Scholar] [CrossRef]
- Joseph, B.; Ramteke, P.W.; Kumar, P.A. Studies on the enhanced production of extracellular lipase by Staphylococcus epidermidis. J. Gen. Appl. Microbiol. 2006, 52, 315–320. [Google Scholar] [CrossRef]
- Haki, G.D.; Rakshit, S.K. Developments in industrially important thermostable enzymes: A review. Bioresour. Technol. 2003, 89, 17–34. [Google Scholar] [CrossRef]
- Pimentel, M.C.; Krieger, N.; Coelho, L.C.; Fontana, J.O.; Melo, E.H.; Ledingham, W.M.; Lima Filho, J.L. Lipase from a Brazilian strain of Penicillium citrinum. Appl. Biochem. Biotechnol. 1994, 49, 59–74. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, A.; Kumar, A.; Singh, D. Lipase from Bacillus pumilus RK31: Production, purification and some properties. World Appl. Sci. J. 2012, 16, 940–948. [Google Scholar]
- Zhang, A.; Gao, R.; Diao, N.; Xie, G.; Gao, G.; Cao, S. Cloning, expression and character-ization ofan organic solvent tolerant lipase from Pseudomonas fluorescens JCM5963. J. Mol. Catal. B Enzym. 2009, 56, 78–84. [Google Scholar] [CrossRef]
- Soleymani, S.; Alizadeh, H.; Mohammadian, H.; Rabbani, E.; Moazen, F.; Sadeghi, H.M.; Shariat, Z.S.; Etemadifar, Z.; Rabbani, M. Efficient media for high lipase production: One variable at a time approach. Avicenna J. Med. Biotechnol. 2017, 9, 82–86. [Google Scholar] [CrossRef]
- Sabat, S.; Murthy, V.K.; Pavithra, M.; Mayur, P.; Chandavar, A. Production and characterisation of extracellular lipase from Bacillus stearothermophilus MTCC 37 under different fermentation conditions. Int. J. Eng. Res. Appl. 2012, 2, 1775–1781. [Google Scholar]
- Rathi, P.; Saxena, R.K.; Gupta, R. A novel alkaline lipase from Burkholderia cepacia for detergent formulation. Process Biochem. 2001, 37, 187–192. [Google Scholar] [CrossRef]
- Kumar, S.; Kikon, K.; Upadhyay, A.; Kanwar, S.S.; Gupta, R. Production, purification, and characterization of lipase from thermophilic and alkaliphilic Bacillus coagulans BTS-3. Protein Expr. Purif. 2005, 41, 38–44. [Google Scholar] [CrossRef]
- Eltaweel, M.A.; Rahman, R.N.Z.R.A.; Salleh, A.B.; Basri, M. An organic solvent-stable lipase from Bacillus sp. strain 42. Ann. Microbiol. 2005, 55, 187–192. [Google Scholar]
- Bokhorst, S.; Huiskes, A.; Convey, P.; Van Bodegom, P.M.; Aerts, R. Climate change effects on soil arthropod communities from the Falkland Islands and the Maritime Antarctic. Soil Biol. Biochem. 2008, 40, 1547–1556. [Google Scholar] [CrossRef]
- Convey, P.; Bowman, V.C.; Chown, S.L.; Francis, J.E.; Fraser, C.; Smellie, J.L.; Storey, B.; Terauds, A. Ice-bound Antarctica: Biotic consequences of the shift from a temperate to a polar climate. In Mountains, Climate, and Biodiversity; Hoorn, C., Perrigo, A., Antonelli, A., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2018; pp. 345–355. [Google Scholar]
- Samad, M.Y.A.; Razak, C.N.A.; Salleh, A.B.; Yunus, W.Z.W.; Ampon, K.; Basri, M. A plate assay for primary screening of lipase activity. J. Microbiol. Meth. 1989, 9, 51–56. [Google Scholar] [CrossRef]
- Scholze, H.; Stutz, H.; Paltauf, F.; Hermetter, A. Fluorescent inhibitors for the qualitative and quantitative analysis of lipolytic enzymes. Anal. Biochem. 1999, 276, 72–80. [Google Scholar] [CrossRef]
- Ganasen, M.; Yaacob, N.; Rahman, R.N.Z.R.A.; Leow, A.T.C.; Basri, M.; Salleh, A.B.; Ali, M.S.M. Cold-adapted organic solvent tolerant alkalophilic family I.3 lipase from an Antarctic Pseudomonas. Int. J. Biol. Macromol. 2016, 92, 1266–1276. [Google Scholar] [CrossRef]
- Shukor, M.Y.; Hassan, N.A.A.; Jusoh, A.Z.; Perumal, N.; Shamaan, N.A.; Maccormack, W.P.; Syed, M.A. Isolation and characterization of a Pseudomonas diesel-degrading strain from Antarctica. J. Environ. Biol. 2009, 30, 1–6. [Google Scholar]
- Thomas, A.D. Evaluation of the API 20E and microbact 24E systems for the identification of Pseudomonas pseudomallei. Vet. Microbiol. 1983, 8, 611–615. [Google Scholar] [CrossRef]
- Ali, M.S.M.; Ganasen, M.; Rahman, R.N.Z.R.A.; Chor, A.L.T.; Salleh, A.B.; Basri, M. Cold-adapted RTX lipase from antarctic Pseudomonas sp. strain AMS8: isolation, molecular modeling and heterologous expression. Protein J. 2013, 32, 317–325. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |
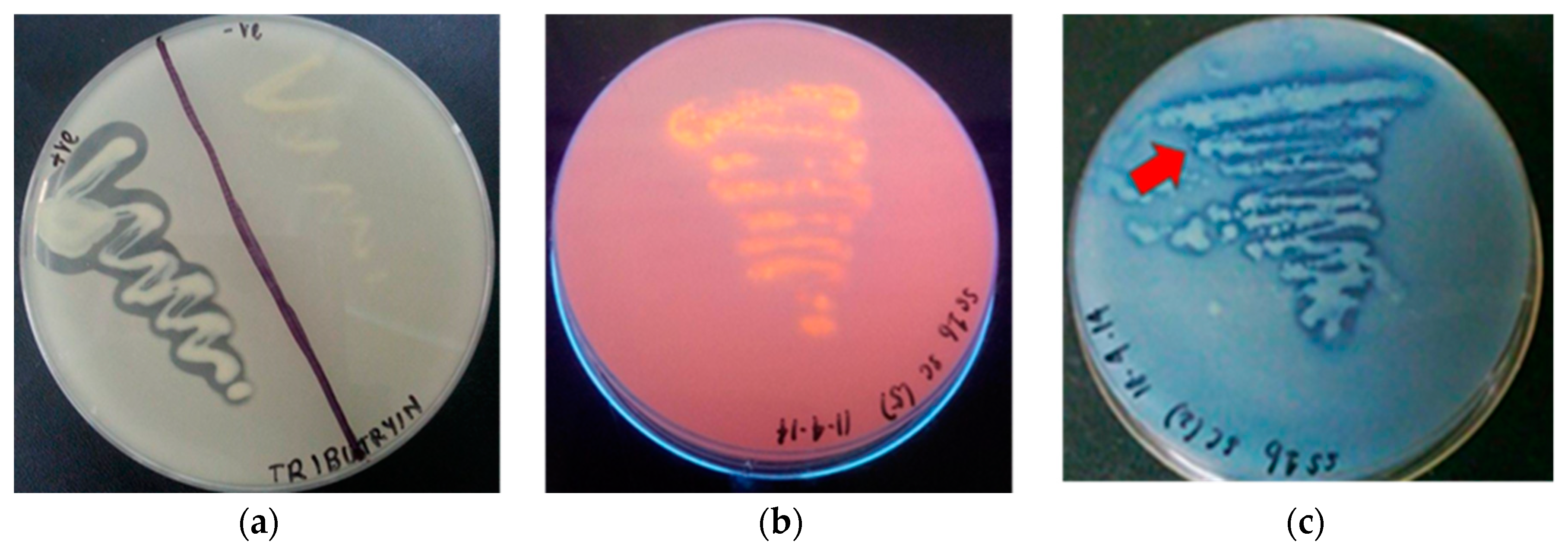
 ) colour of dye indicators on triolein agar.
) colour of dye indicators on triolein agar.
) colour of dye indicators on triolein agar.
) colour of dye indicators on triolein agar.

 ) with other Pseudomonas sp. strains obtained from the polar regions.
) with other Pseudomonas sp. strains obtained from the polar regions.
) with other Pseudomonas sp. strains obtained from the polar regions.
) with other Pseudomonas sp. strains obtained from the polar regions.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Lipase Gene Copy Numbers (n = 3) | s.d. (±) |
|---|---|---|
| LSK14 | 1.02 × 1010 | 0.16 |
| LSK25 | 2.45 × 1014 | 0.11 |
| CF1 | 6.35 × 108 | 0.12 |
| BP1 | 3.04 × 1010 | 0.11 |
| Sampling Sites | Description | Location |
|---|---|---|
| Area surrounding Signy Station (LSK) | Around the station living quarters, with high human activity, old whaling station | 60°42′56”S 45°36′22”E |
| Bernsten Point (BP) | Site of previous Signy station buildings, historical oil spillages | 60°40′16”S 45°34′42”E |
| Cemetery flats (CF) | Site of historical whaling industry activity, including oil spills and other pollution | 60°41′16”S 45°38′42”E |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salwoom, L.; Raja Abd Rahman, R.N.Z.; Salleh, A.B.; Mohd. Shariff, F.; Convey, P.; Pearce, D.; Mohamad Ali, M.S. Isolation, Characterisation, and Lipase Production of a Cold-Adapted Bacterial Strain Pseudomonas sp. LSK25 Isolated from Signy Island, Antarctica. Molecules 2019, 24, 715. https://doi.org/10.3390/molecules24040715
Salwoom L, Raja Abd Rahman RNZ, Salleh AB, Mohd. Shariff F, Convey P, Pearce D, Mohamad Ali MS. Isolation, Characterisation, and Lipase Production of a Cold-Adapted Bacterial Strain Pseudomonas sp. LSK25 Isolated from Signy Island, Antarctica. Molecules. 2019; 24(4):715. https://doi.org/10.3390/molecules24040715
Chicago/Turabian StyleSalwoom, Leelatulasi, Raja Noor Zaliha Raja Abd Rahman, Abu Bakar Salleh, Fairolniza Mohd. Shariff, Peter Convey, David Pearce, and Mohd Shukuri Mohamad Ali. 2019. "Isolation, Characterisation, and Lipase Production of a Cold-Adapted Bacterial Strain Pseudomonas sp. LSK25 Isolated from Signy Island, Antarctica" Molecules 24, no. 4: 715. https://doi.org/10.3390/molecules24040715
APA StyleSalwoom, L., Raja Abd Rahman, R. N. Z., Salleh, A. B., Mohd. Shariff, F., Convey, P., Pearce, D., & Mohamad Ali, M. S. (2019). Isolation, Characterisation, and Lipase Production of a Cold-Adapted Bacterial Strain Pseudomonas sp. LSK25 Isolated from Signy Island, Antarctica. Molecules, 24(4), 715. https://doi.org/10.3390/molecules24040715