1. Introduction
Multidrug-resistant microorganisms are rapidly spreading throughout the world leading to treatment failures [
1]. Antibacterial resistance in humans is affected by an inappropriate or excessive use of antibiotics in human health but is also partially affected by the use of antibiotics in animal-rearing. Some national action plans, such as restrictive measures of antibiotic use in human and animal health, are now imposed in all parts of the world but the problem of antimicrobial resistance is far from being solved [
2,
3]. Therefore, the discovery of new antimicrobial compounds is crucial.
Several plant-derived natural products, characterized by a huge structural diversity, including phenolic compounds, are cited as antimicrobial agents and resistance-modifying agents (RMAs) [
4]. They could constitute a valuable interim solution, until new classes of antibiotics are discovered [
5].
Methicillin-resistant
Staphylococcus aureus (MRSA) strains are considered as a very urgent health problem because of their propagation in the last 15 years in the elderly and in immunocompromised patients, mainly due to their increasing implication in nosocomial infections and the lack of development of new antimicrobials. In 2016, the percentage of MRSA among all
S. aureus isolates remained above 25% in several countries in Southern Europe and greater than 15% in France, especially for invasive isolates [
6]. MRSA is also one of the principal multi-resistant bacterial pathogen responsible for complicated skin and hospital-acquired infections, associated with high mortality rates [
7,
8]. Furthermore, these strains are very often involved in infections of diabetic foot ulcers characterized by frequent complications and risks of lower-limb amputations [
9].
S. aureus has the ability, like most Gram-positive organisms, to acquire resistance to practically all useful antibiotics. Several mechanisms can explain this resistance: modification of target sites, enzymatic degradation or structural modification of antibiotics and, expression of efflux pumps [
5,
10,
11,
12]. These bacteria also produce virulence factors involved in pathogenesis like adhesion and biofilm formation, which enhance bacterial resistance [
13,
14].
On the other hand, many parasitic infections deserve a special attention due to the lack of effective treatments.
Trypanosoma brucei is a parasite conveyed by the tsetse fly and responsible for the Human African Trypanosomiasis (HAT) or sleeping sickness. This fatal disease if untreated leads to central nervous system disturbances, including sensory, motor and psychic troubles and neuroendocrine abnormalities [
15,
16]. Leishmaniasis, caused by different
Leishmania species, is responsible for chronic skin and visceral diseases transmitted by sand-flies [
17].
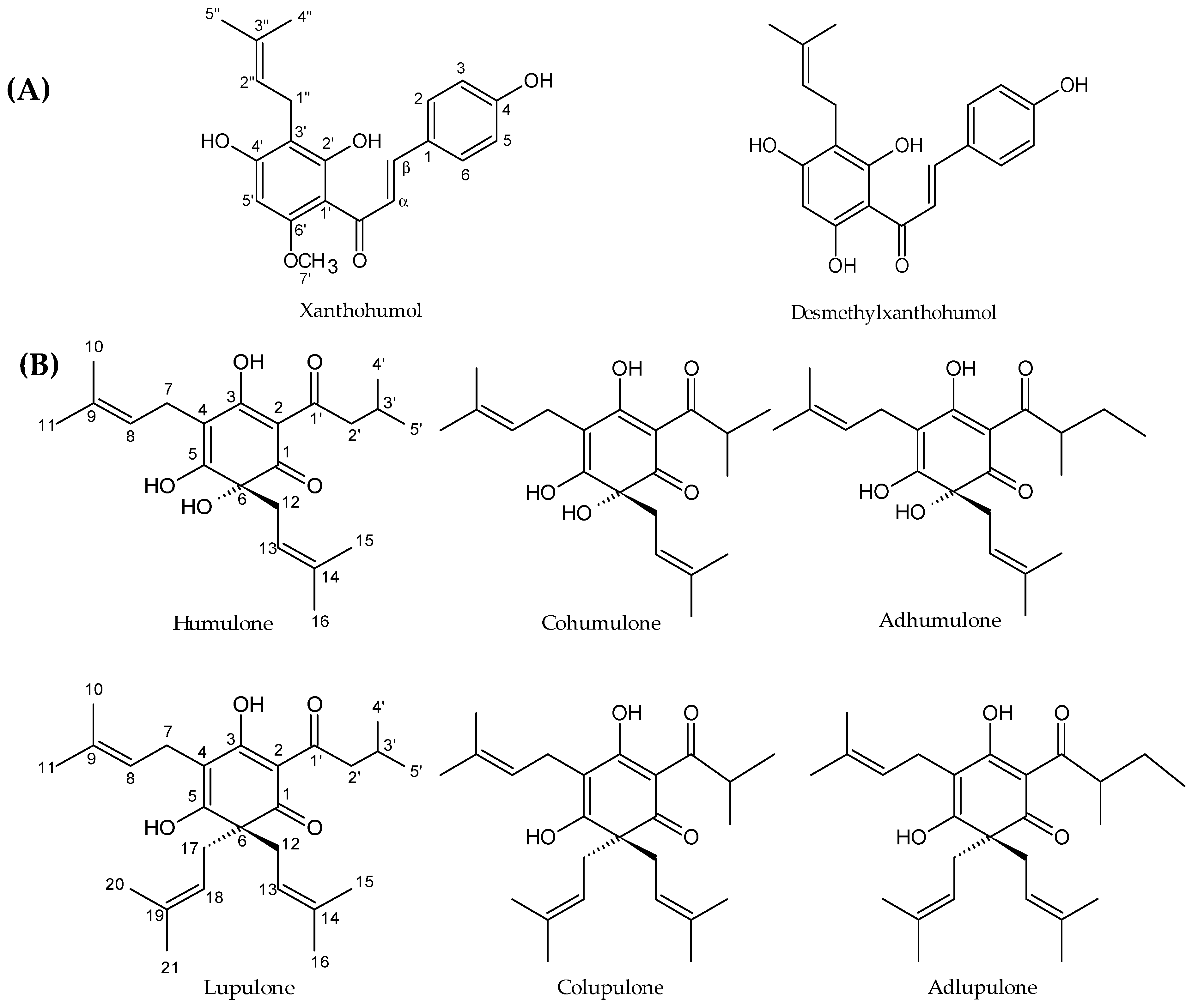
Hop (
Humulus lupulus L.) is a climbing dioecious plant belonging to the Cannabaceae family. This species is cultivated worldwide for its female inflorescences (cones), usually called “hops”, which are used in the brewing industry. The compounds sought by brewers are prenylated acylphloroglucinol derivatives, also called bitter acids and in particular α-acids (humulone derivatives) which are isomerized into iso-α-acids during the brewing process. These compounds confer some bitterness and antiseptic properties to beer [
18]. This plant is also known to have many pharmacological activities, including sedative, oestrogenic, anti-inflammatory, chemopreventive and antimicrobial activities [
19]. The antimicrobial activity is mainly attributed to prenylated acylphloroglucinols, in particular to α-acids and β-acids (lupulone derivatives) and to prenylated chalcones, including xanthohumol (
Figure 1) [
20,
21,
22]. Few studies, however, describe the activity of this plant and especially of its specialized metabolites against multidrug resistant strains [
14,
23] and parasites [
24,
25]. In addition, the mechanisms of the inhibitory activities observed are not really expanded upon now.
In this context, we studied the antimicrobial potential of hop phenolic compounds against MRSA strains, Trypanosoma brucei brucei (Tbb) and Leishmania mexicana mexicana (Lmm). We opted for a fractionation using Centrifugal Partition Chromatography (CPC) and preparative High-Performance Liquid Chromatography (HPLC) to purify the major antimicrobial phenolic compounds from the most active sub-extract of hop. Purified compounds were then quantified in hop extracts. Their antibacterial potential against MRSA clinical isolates was assessed. The most promising compounds were then selected for further experiments in order to strengthen our knowledge of their action on the bacterial strains, through kill curves, post-antibiotic effects, anti-biofilm and synergy assays. The antiparasitic activity of some chalcones and acylphloroglucinols against Tbb and Lmm was also evaluated. Their cytotoxicity activity was determined on various human cell lines.
3. Discussion
Multi-drug resistance (MDR) bacteria, like methicillin-resistant
Staphylococcus aureus (MRSA), present a major challenge for the medical community in the treatment of infections, such as diabetic foot infections [
6,
35]. The discovery of new antibiotics is not fast enough to offset the global spread of resistant pathogens. At the same time, we can observe a worrying emergence of resistance to some of the newer antibiotic agents [
36,
37,
38]. Hence, the development of combination of current agents with other type of resistance-modifying agents, such as natural antibacterial agents, can be an alternative strategy to overcome MDR [
39].
In this context, we evaluated the antibacterial potential of hop extracts and more particularly of three pure compounds isolated from hops (xanthohumol, desmethylxanthohumol and lupulone) towards MRSA, one of the most aggressive agents, with different approaches [
33]. We began by a classical MIC determination, which is only an endpoint method, followed by kill-time curves assessing the time dependent effect and synergistic studies, in combination with antibiotics, giving more information on their resistance modifying potential. The checkerboard method is the preferred technique of choice to analyze these interactions, with however a certain variability in the methods of interpretation [
32]. The study of a post-antibiotic effect (PAE), also applicable to substances other than antibiotics, made it possible to follow the regrowth of bacteria after inactivation of the antibacterial substance after a defined contact time at an active concentration. When damaged bacteria need some time to recover in comparison to the control, the regrowth will be lowered, which means in clinical situations that administration intervals of the antimicrobial substances can be delayed. Another innovative point is the effect on biofilms, concerning about 60% of infections. In biofilms, the bacteria are surrounded by a thick layer of extracellular polysaccharides which makes them inaccessible to antibiotics but also to defenses of the immune system. Furthermore, the lack of nutrients lowers dramatically the bacterial multiplication. As most of the antibiotics act on mechanisms implicated in multiplication, this is the second reason for their lack of action on biofilms. Influence on biofilms can be studied in two manners: destruction of established biofilms or inhibition of their formation. The first manner is more relevant for clinicians as at the time of diagnosis the biofilm is already established.
The antibacterial activity of hops has been known for many years [
40,
41] but has been poorly evaluated against resistant strains. We showed here their effects on
Corynebacterium,
Enterococcus,
Mycobacterium,
Staphylococcus and
Streptococcus strains, some being resistant to antibiotics (
Table 1). Their effect on
Bacillus,
Streptomyces and
Micrococcus strains was previously underlined [
22,
41]. Hops extracts are also able to combat some human pathogenic bacteria found in food, such as
Clostridium perfringens and
Listeria monocytogenes [
42,
43]. The screening conducted on some
C. albicans strains showed that the spectrum of activity of hops extracts against these yeasts is weak, only two strains were susceptible to our extracts. The
C. albicans ATCC 10,231 reference strain was previously tested by Langezaal et al. [
44] who also found an efficiency of cones extract. Other studies have demonstrated that hops extracts are more efficient on bacteria than yeasts, showing no effect of hops constituents on
Saccharomyces strains [
41]. Contrarily to Langezaal et al. [
44] and Abram et al. [
45] who have detected a slight activity of cones extracts against
E. coli strains, our extracts were inactive towards Gram-negative bacteria. Stems, leaves and rhizomes have been rarely studied. According to our results, their crude hydro-ethanolic extracts showed a weak antimicrobial activity in comparison with cones, as already highlighted for leaves against some bacteria [
45]. Nevertheless, the rhizomes extract appeared to be more active than leaves and stems extracts (
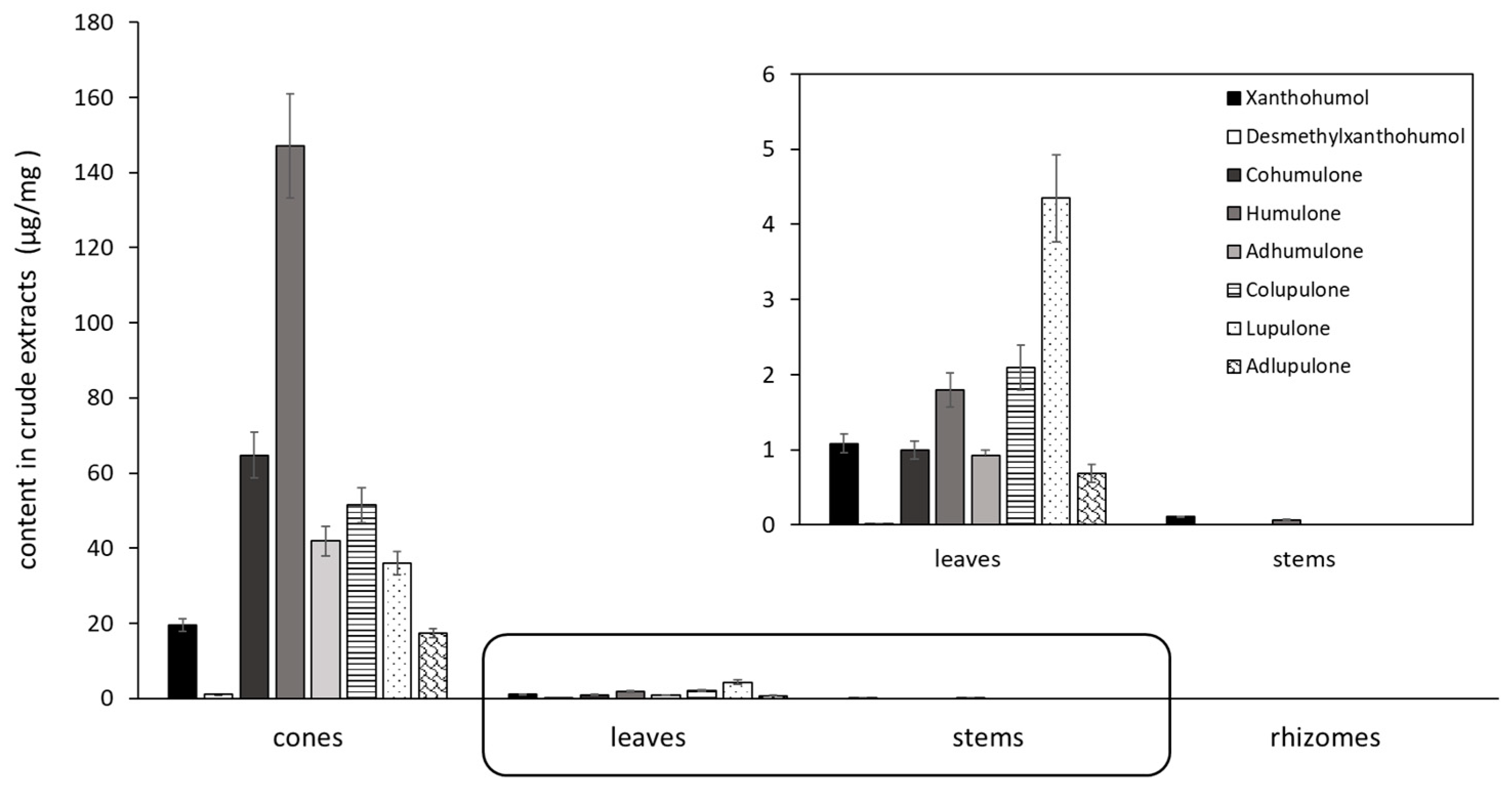
Table 1). Previous research on the quantification of the phenolic hop compounds in cones and leaves by LC-UV is reported in the literature [
46]. With our method of quantification, no prenylated phenolic compounds could be quantified in the rhizome extract. Consequently, the activity of rhizomes could be related to the presence of other metabolites. The antibacterial activity of cones was mainly attributed to apolar compounds because the DCM sub-extract of cones was more active than the crude extract of the same plant part (
Table 2). The content of phenolic compounds in hops is influenced in particular by the cultivar, the growth location, the field conditions, the climate, which may explain the differences in activities found in the literature [
47,
48].
Phenolic compounds are known to be an anti-staphylococcal class of metabolites. In our study, all purified hops prenylated chalcones and acylphloroglucinol derivatives showed an antibacterial activity towards selected
S. aureus strains (
Table 3). In the literature, the antibacterial activity of hops is mainly linked to acylphloroglucinol derivatives. This activity is enhanced by the degree of hydrophobicity of the compound [
49]. The number and the length of the side chains, because of their interaction with the bacterial cell wall, is stated to influence positively the antibacterial action, so lupulone derivatives are more efficient than humulone derivatives [
22,
42], which is in accordance with our results. Lupulone is much more active than humulone with respective MICs ranging from 0.6 to 1.2 and 78 to 156 µg/mL towards MRSA strains (
Table 3). With the exception of lupulone, we also demonstrated that xanthohumol and desmethylxanthohumol are more active than other bitter acids. Xanthohumol showed MIC ranging from 9.8 to 19.5 µg/mL against selected
S. aureus strains. In comparison with one of the most studied promising antimicrobial chalcone, licochalcone A, which showed MIC ranging from 2 to 15 µg/mL against Gram-positive bacteria including
S. aureus [
50], the activity of xanthohumol is in the same range. Several studies have focused on the antibacterial potential of xanthohumol which showed various MICs from 2 to 125 µg/mL on
S. aureus, depending on the strain’s resistance profile and the compound’s purity [
14,
22]. According to our results, desmethylxanthohumol was slightly less active than xanthohumol, with MICs ranging from 19.5 to 39 µg/mL against
S. aureus strains (
Table 3). The structural difference between the two molecules is the 6′-methoxyl group for xanthohumol replaced by a 6′-phenol group for desmethylxanthohumol (
Figure 1). The 6′-phenol substituent is considered as a crucial group in the equilibrium chalcone-flavanone but is not expected to contribute to the activity [
51]. According to Ávila et al. [
51], the methylation of the 6-phenol group could lead to less active compounds, which is not the case in our study. Desmethylxanthohumol is known for its antioxidant and apoptotic activities [
52] but to our knowledge, no antibacterial potential was highlighted. Olivella et al. [
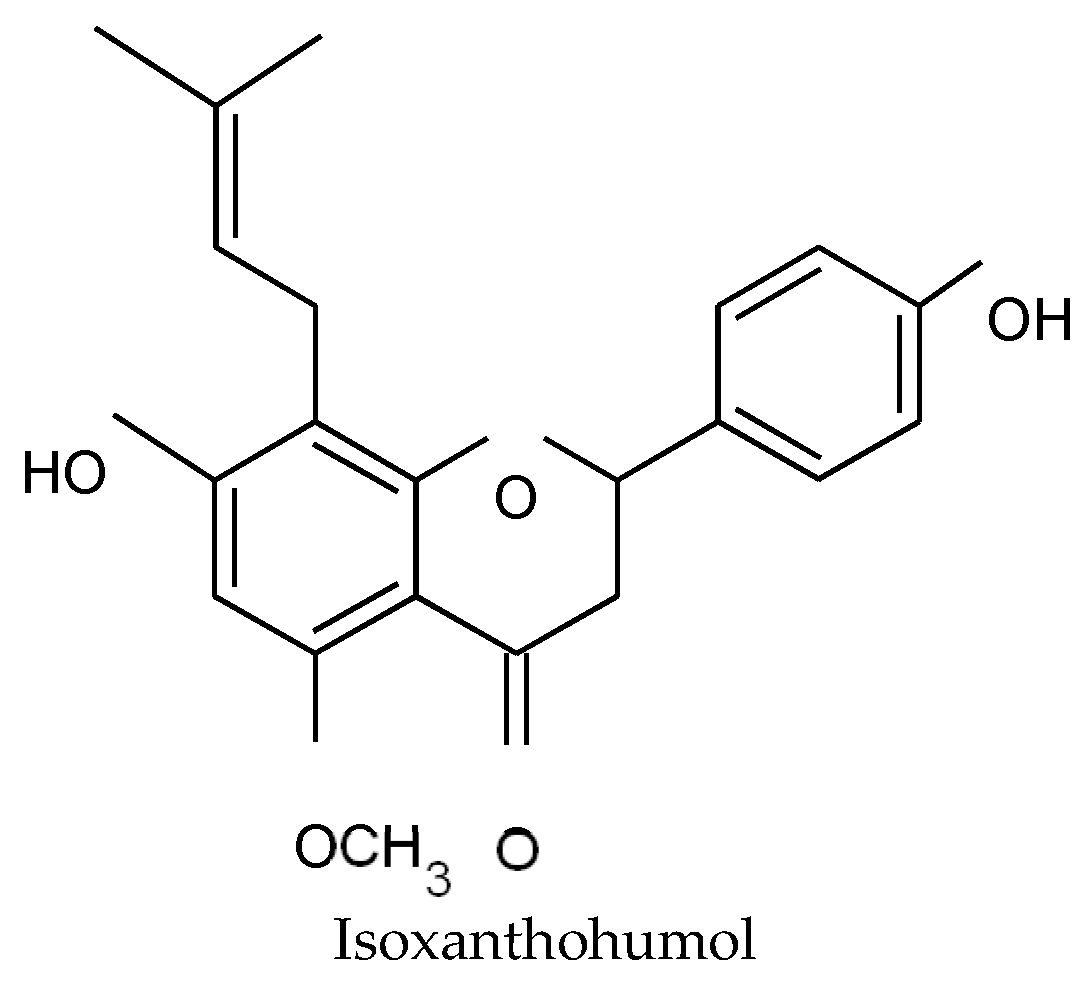
53] have demonstrated that chalcones have the most favorable structure for a bacteriostatic action, in comparison with flavanones. In rats, xanthohumol is partially absorbed by intestinal cells and transported in blood. The non-absorbed part of xanthohumol can be transformed into the corresponding flavanone, isoxanthohumol, by the intestinal microbiota [
54]. The conversion of isoxanthohumol into 8-prenyl-naringenin occurs only in the colon and not in the stomach and small intestine [
55,
56]. In this context, we also tested the activity of isoxanthohumol (
Figure 4) against the strain MRSA T28.1. This compound showed a MIC equal to 39 µg/mL. It is interesting to note that, even if isoxanthohumol is less active than xanthohumol, its MIC was found to be of similar than desmethylxanthohumol.
The antibacterial potential of xanthohumol, desmethylxanthohumol and lupulone against MRSA is therefore promising. Their MICs are in the same order of magnitude as those obtained for antibacterial phenolic compounds known for their important activity against MRSA strains, with MICs varying between 1.56 and 125 µg/mL [
4]. Most of them are known to be antibacterial compounds but they are not used to combat strains resistant to antibiotics. Some hops metabolites also showed antibacterial activities against others strains such as:
Bacillus subtilis,
Clostridium difficile,
Enterococcus strains and
Streptococcus strains for the Gram-positive bacteria;
Bacteroides fragilis,
Helicobacter pylori and
Yersinia enterocolitica for the Gram-negative bacteria [
20].
To overcome bacterial resistance, one of the strategies employed is the use of a combination of drugs, such as antibiotics combined with natural products, which has already shown promising results [
57]. The checkerboard is a method to establish the best combination of products increasing the activity [
32,
58]. The antibacterial action of hops compounds can be enhanced by combining concomitantly xanthohumol with desmethylxanthohumol or with lupulone. The effect is twice as intense because the MIC is divided by two. By contrast, the combination of desmethylxanthohumol with lupulone leads to an antagonist effect (
Table 4). The combination of natural products with antibiotics may in some cases have a synergistic effect. Xanthohumol and rifampicin can be 8 times more active when they are combined. Furthermore, for both xanthohumol and lupulone, the MIC for oxacillin drops below the threshold concentration of 2 µg/mL for oxacillin resistance, which means that in the presence of these synergistic compounds the strains will no longer be classified as MRSA, so reverting their resistance. Desmethylxanthohumol has an additive effect with oxacillin but this interaction does not render the strain susceptible to this antibiotic. Desmethylxanthohumol also has a promising interaction with ciprofloxacin and gentamicin. Some authors have previously detected synergies of xanthohumol and lupulone with polymyxin, ciprofloxacin or tobramycin [
59]; and of xanthohumol with oxacillin or linezolid [
14].
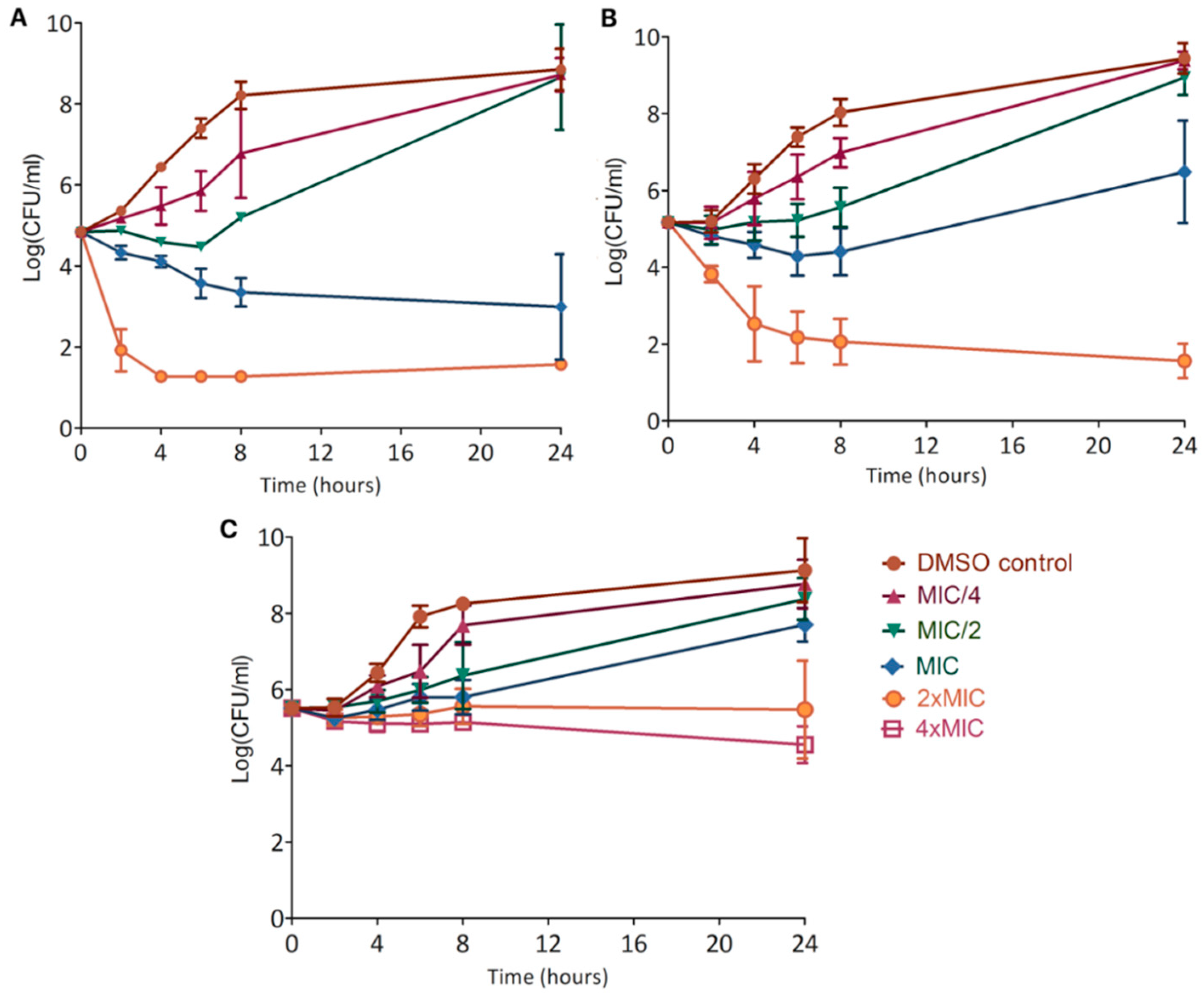
Kill curves demonstrated a great bactericidal action of xanthohumol at the MIC, of desmethylxanthohumol from 2 × MIC, whereas lupulone is slightly bactericidal after 24 h only at 4 × MIC (
Figure 5). Comparing the activity of the two chalcones, desmethylxanthohumol showed a lower bactericidal action at the MIC than xanthohumol, which is probably linked to the presence of the 6′-hydroxyl group (
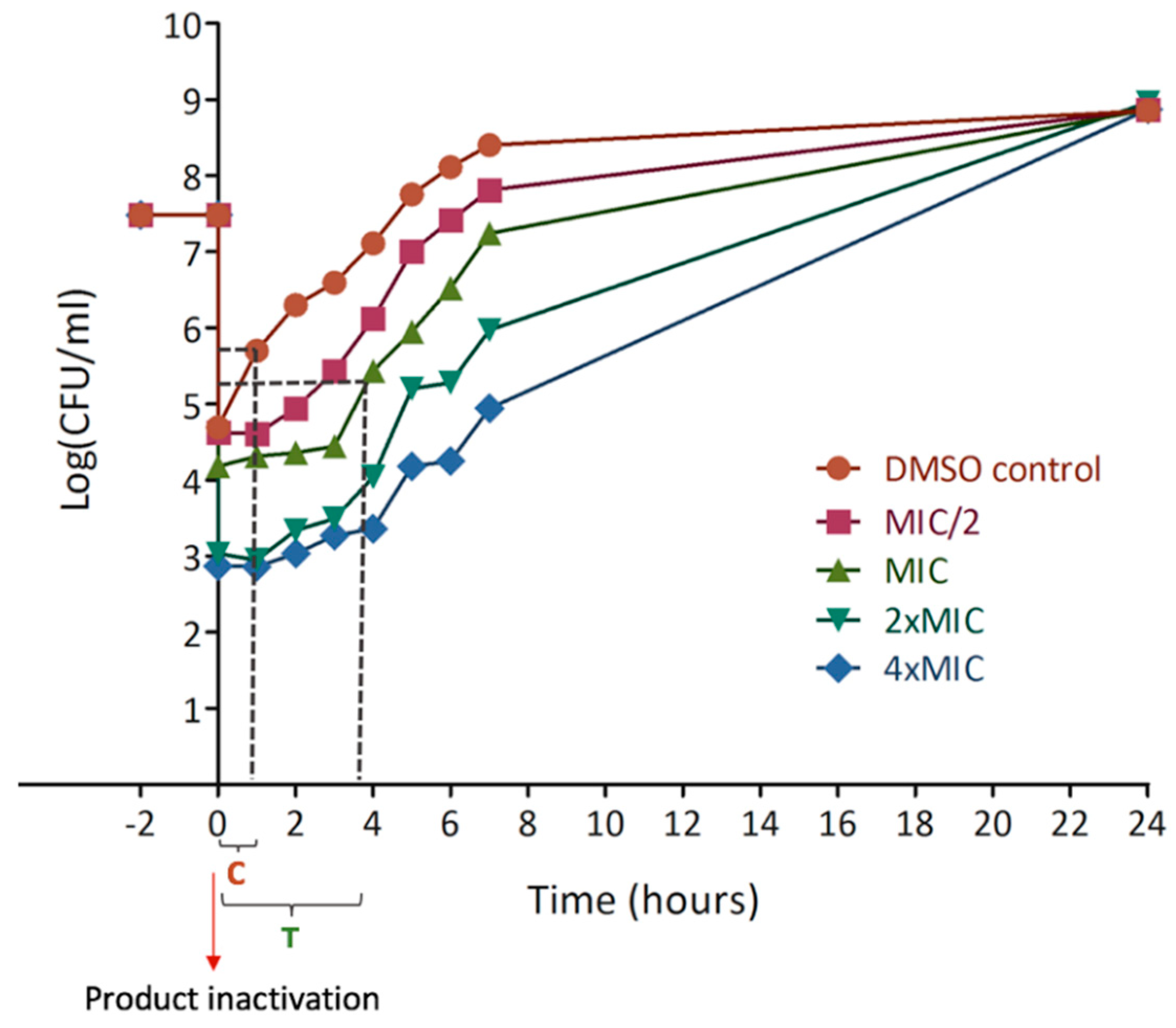
Figure 1). Post-antibiotic effect is a part of pharmacodynamic studies, showing that xanthohumol and desmethylxanthohumol cause a significant delay for regrowth (
Table 6). It means that the bacterial growth remains inhibited even after the product has been inactivated or metabolized by the body. These data provide an indication of the delay between two applications in a clinical situation. This is the first time that PAE is analyzed for hop compounds. This effect underlines an important reduction time for recovery which means that in vivo models will have to be checked for delay in drug administration.
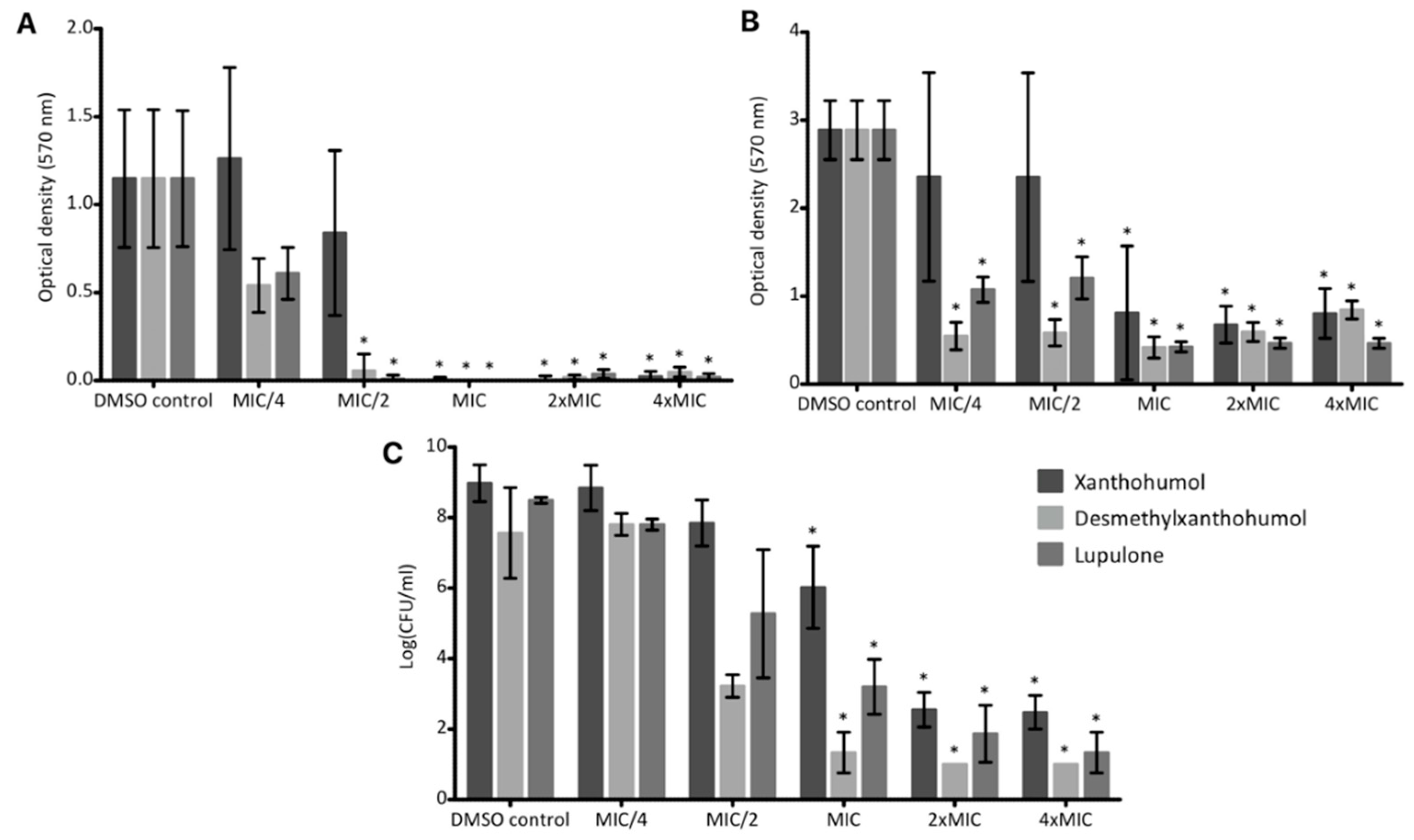
Xanthohumol, desmethylxanthohumol and lupulone showed an inhibition of the biofilm formation of the
S. aureus model strain on abiotic surface, with a sub-inhibitory action for desmethylxanthohumol and lupulone (
Figure 7). Rozalski et al. studied the anti-adherent potential of a hops extract enriched in xanthohumol, pure xanthohumol and a spent hops extract rich in various common flavonols and flavanols [
14]. They demonstrated a potent effect of xanthohumol on the biofilm formation at the MIC with 86.5% of inhibition. In comparison, our results showed an inhibition close to 100% at the MIC for the selected MRSA clinical isolate. In addition, we have also demonstrated that a previous formation of the biofilm does not prevent hops compounds to act on bacteria. In both cases, desmethylxanthohumol and lupulone seem to be more effective than xanthohumol, with an inhibition of the biofilm formation and a biofilm destruction at sub-inhibitory concentrations (
Figure 7). Bogdanova et al. [
23] also showed an anti-biofilm potential of some hops compounds but lower than that of our study. This result could be related to a lower purity of their products (from 82 to 87% in Reference [
23]). This potential has been confirmed on a synthetic bone substitute which is the natural colonized substrate of
S. aureus T28.1 (
Figure 7). Even if the inhibition of the biofilm formation is found somewhat less intense in bone substitute than on inert surface, this potential remains very interesting, in particular for desmethylxanthohumol and lupulone. Moreover, according to our results, the anti-biofilm effect for these two products seems to be independent of the bactericidal effect pointed out with kill curves (
Figure 5). To our knowledge, the anti-biofilm effect has never been assessed for hops compounds on bone substitutes. These data confirmed the promising potential of hops compounds to tackle MRSA not only on planktonic cells (MIC, kill curves) but also on biofilms approaching clinical situations.
Diabetic foot infections (DFI) affect one ulcerated foot out of two and in many cases lead to serious complications [
60]. About 50% of patients hospitalized for a DFI suffer from an osteomyelitis and the prevalence of MRSA is often associated [
61]. Low diffusion in necrotic tissues emphasizes topical antibiotic therapy for the management of mildly to moderately infected diabetic foot ulcers which has shown satisfactory results in some cases, allowing high concentrations of antibiotics at the site of infection without potentially toxic systemic levels [
62]. Some medical devices such as beads loaded with antibiotics can bring high concentrations of local antibiotics for a long time in the case of deep wounds [
63,
64]. In addition, some topical antimicrobial agents, such as impregnated wound dressings with antimicrobials, could be of interest in the prevention or possibly the treatment of mild infections [
65]. DFI generates many problems in clinical practice in terms of both diagnosis and therapeutic care mainly due to formation of persistent biofilms. The anti-biofilm action of hops metabolites both on artificial surface and on a synthetic bone substitute could bring out a new perspective to treat infected diabetic foot ulcers, an emerging public health problem. Thus, hop phenolic compounds with their dual action, antibacterial and anti-biofilm, are potential agents in the treatment of infections due to MRSA. Their additive or synergistic action with antibiotics could render treatments more effective and thus could prevent potential systemic toxicity if used in topical application. In this context, we evaluated the antiproliferative activity of the three phenolic compounds on different human cell lines and in particular against the human osteoblasts MG-63 cell line. After 48 h exposure, we showed a toxicity of xanthohumol on the targeted cell line. In the literature, data on its cytotoxicity depend on the cell type used and is very variable. For example, Ho et al. and Yong et al. have determined respectively an IC
50 of about 75 and 100 µg/mL against a human hepatocellular carcinoma [
66] and a lung cell line [
67]. These concentrations are higher than the active doses reported in our work. In vivo studies have also confirmed the good safety at approximately 1000 mg of xanthohumol/kg of body weight of mice [
68] and up to 180 mg of compound in humans for a short intake [
69]. According to our results, desmethylxanthohumol is the less toxic compound on the MG-63 osteoblastic cells. However, considering its moderate activity, bactericidal and anti-biofilm concentrations would be toxic. To our knowledge, there is no comparison data available in the literature. Special attention has to be paid on lupulone. Its very pronounced antibacterial activity makes it non-toxic at the active doses. Moreover, anti-biofilm concentrations (MIC/4 and MIC/2) lead to a cell viability close to 100%. Comparing with the literature, some authors have determined IC
50 ranging from 3.7 to 4.4 µg/mL on prostate cancer cells, which is close to other results [
70] and IC
50 from 8.3 to 16.6 µg/mL on breast cancer cells [
71]. IC
50 values are always higher than the active doses we have identified. All these data make lupulone a very good candidate for a topical bone application. Further research could be done by combining xanthohumol or desmethylxanthohumol with antibiotics as it would reduce the dose and avoid toxicity.
In addition, we evaluated the antiparasitic activity of the main chalcones and the main acylphloroglucinols of hop against two parasites:
Trypanosoma brucei brucei (Tbb) and
Leishmania mexicana mexicana (Lmm). Human African trypanosomiasis and leishmaniasis are indeed two protozoan infections considered as neglected tropical diseases with a strong impact on human health because in particular fatal if untreated [
16,
17]. Lupulone was the most active compound and humulone the less active. The four compounds tested were more active against Tbb than against Lmm (
Table 6). Data on the antiparasitic activities of hops compounds are quite limited and especially concern xanthohumol. This chalcone was active against
Plasmodium falciparum [
24] and against
Leishmania amazonensis [
72] with IC
50 in the µM range.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}