Anti-Oxidative and Anti-Aging Activities of Porcine By-Product Collagen Hydrolysates Produced by Commercial Proteases: Effect of Hydrolysis and Ultrafiltration


Abstract
:1. Introduction
2. Results and Discussion
2.1. Enzymatic Hydrolysis Effect on Collagen Hydrolysates
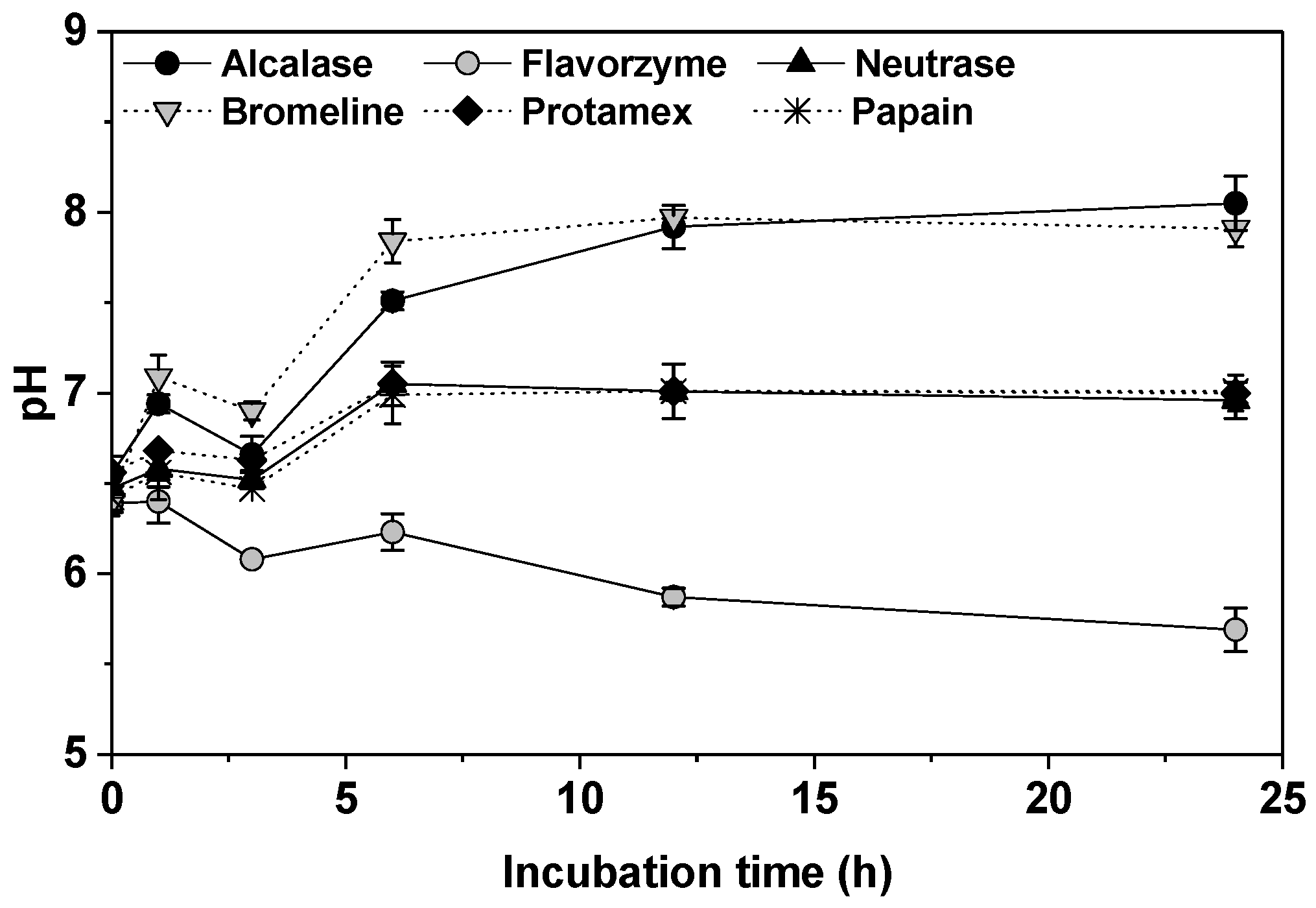
2.1.1. pH
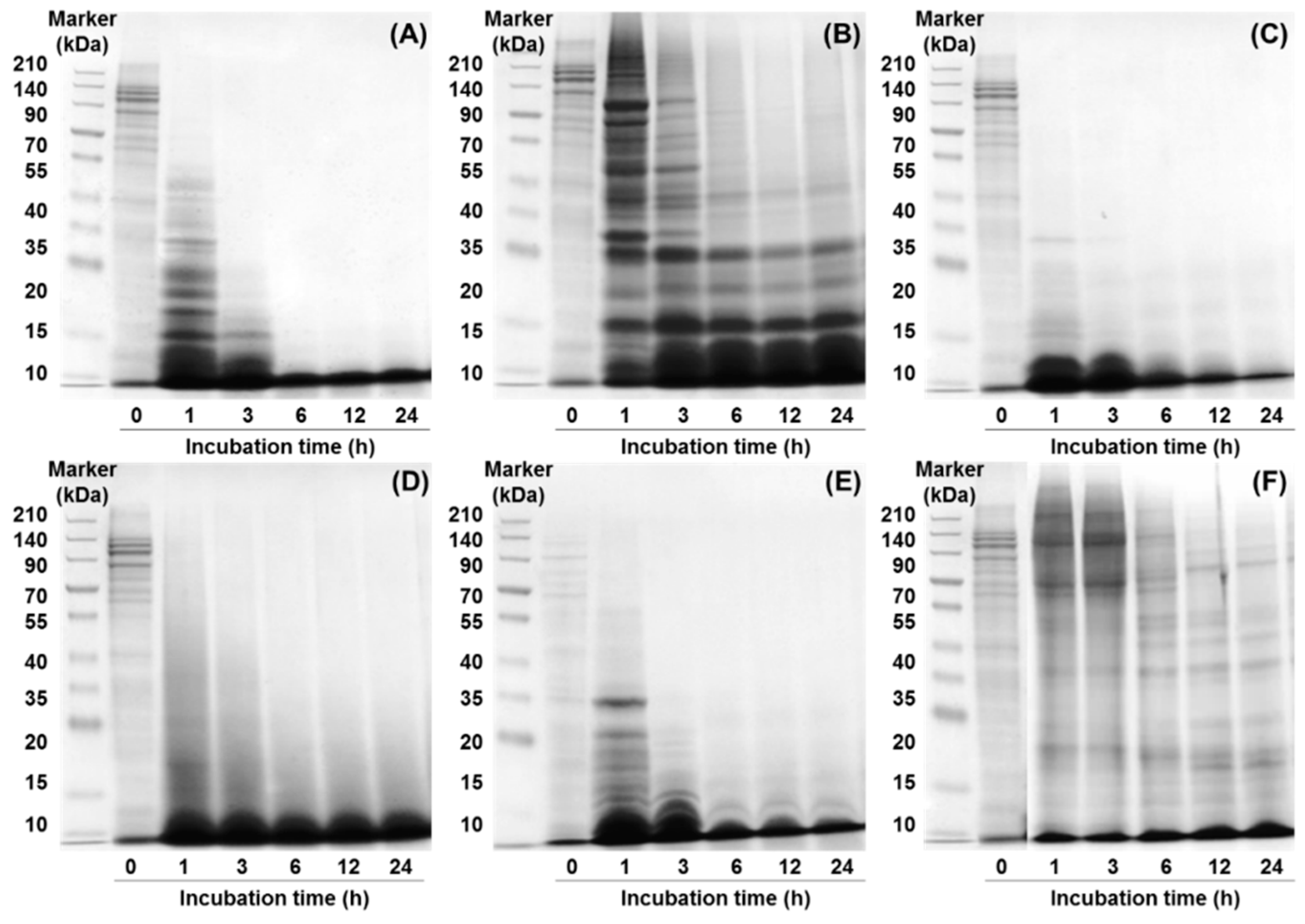
2.1.2. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
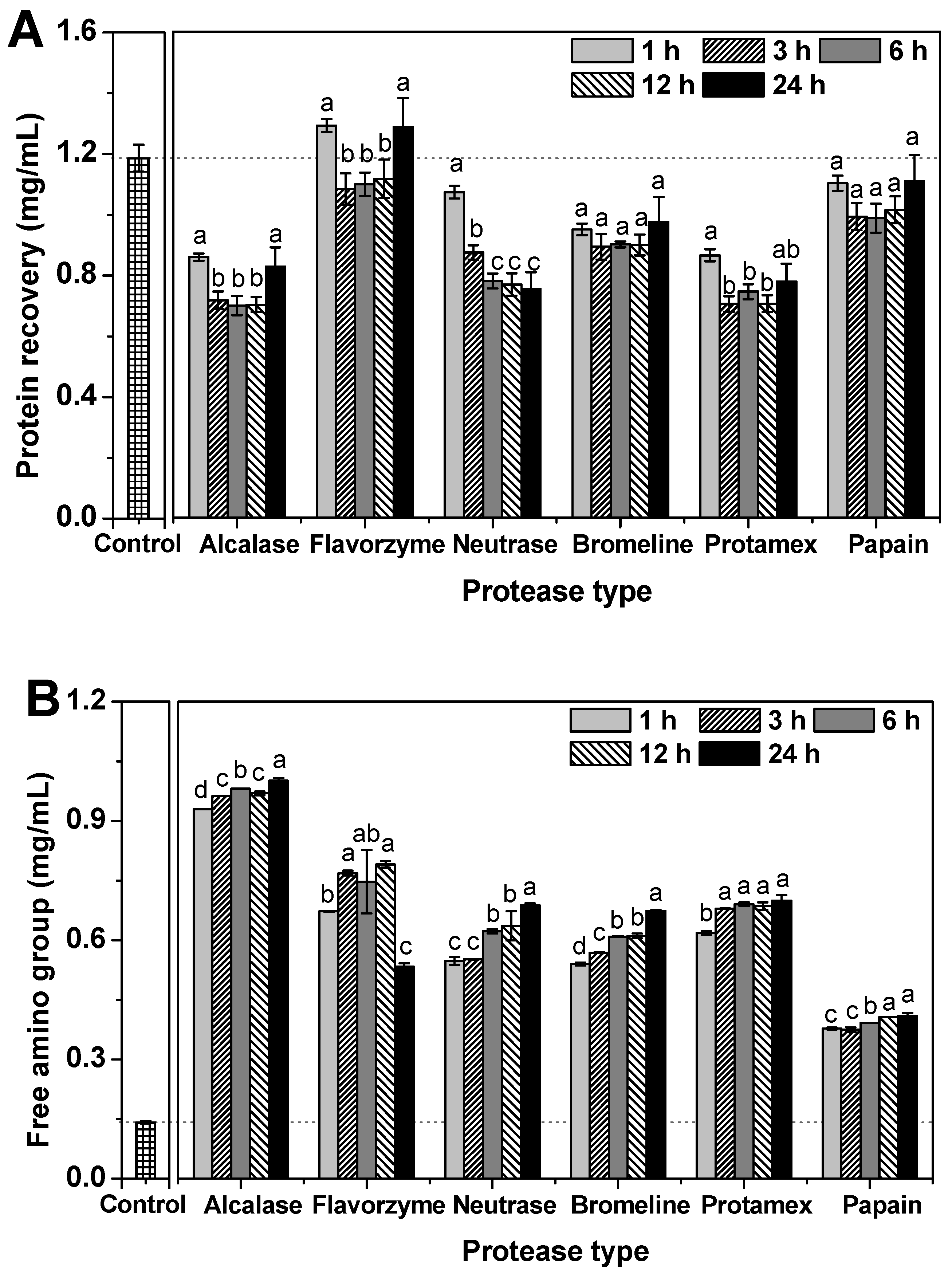
2.1.3. Protein Recovery and Free Amino Group Content
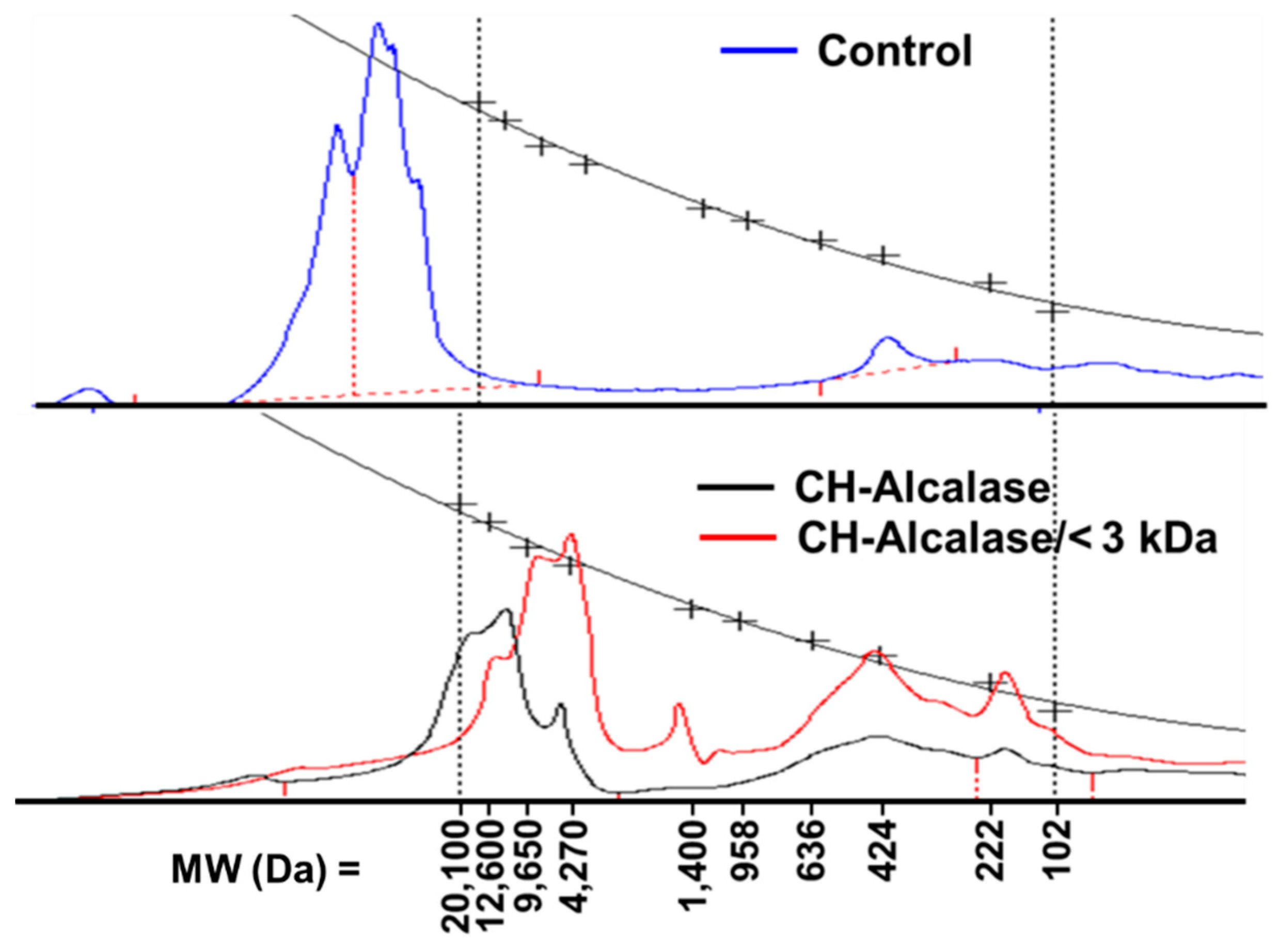
2.2. Ultrafiltration Effect on Collagen Hydrolysate Properties
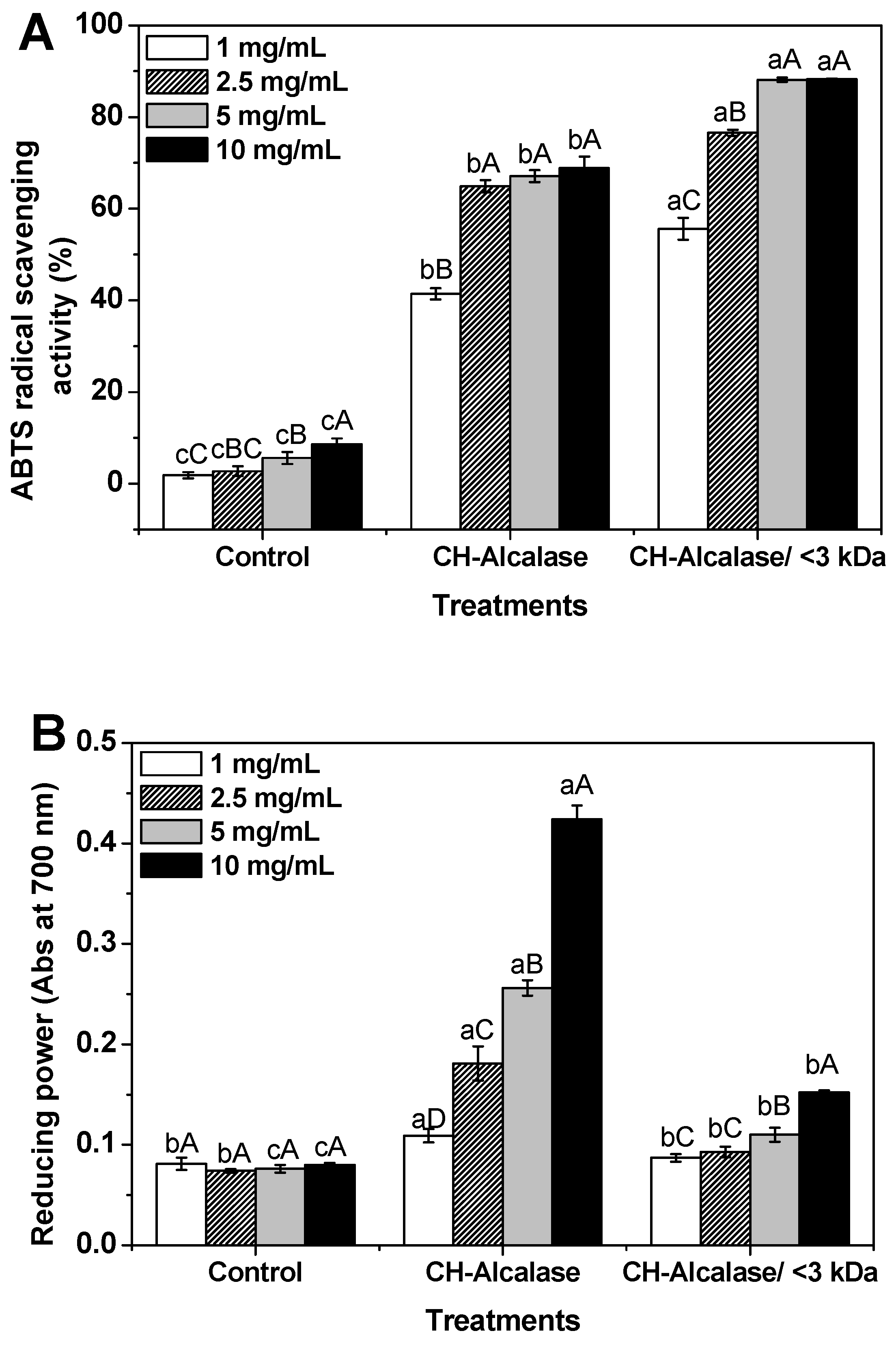
2.3. Antioxidant and Anti-Aging Activities of Collagen Hydrolysates
3. Materials and Methods
3.1. Porcine Skin Pretreatment
3.2. Commercial Proteases and Reagents
3.3. Enzyme Hydrolysis
3.4. Determination of pH, Protein Recovery, Solubility Free Amino Group Content, and Production Yield
3.5. Molecular Weight Distribution
3.5.1. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
3.5.2. Gel Permeation Chromatography (GPC)
3.6. Amino Acid Composition
3.7. Evaluation of Antioxidant Activity
3.7.1. 2,2′-Azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) Radical-Scavenging Activity
3.7.2. Reducing Power
3.8. Evaluation of the Anti-Aging Effect
3.8.1. Inhibition of Tyrosinase Activity
3.8.2. Inhibition of Collagenase Activity
3.8.3. Inhibition of Elastase Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Nakchum, L.; Kim, S.M. Preparation of squid skin collagen hydrolysate as an antihyaluronidase, antityrosinase, and antioxidant agent. Prep. Biochem. Biotechnol. 2016, 46, 123–130. [Google Scholar] [CrossRef]
- Je, J.-Y.; Qian, Z.-J.; Byun, H.-G.; Kim, S.-K. Purification and characterization of an antioxidant peptide obtained from tuna backbone protein by enzymatic hydrolysis. Process Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Jung, K.-H.; Choi, Y.-C.; Chun, J.-Y.; Min, S.-G.; Hong, G.-P. Effects of concentration and reaction time of trypsin, Pepsin, and chymotrypsin on the hydrolysis efficiency of porcine placenta. Korean J. Food Sci. Anim. Resour. 2014, 34, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Koh, B.-B.; Lee, E.-J.; Ramachandraiah, K.; Hong, G.-P. Characterization of bovine serum albumin hydrolysates prepared by subcritical water processing. Food Chem. 2019, 278, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Agyei, D.; Ongkudon, C.M.; Wei, C.Y.; Chan, A.S.; Danquah, M.K. Bioprocess challenges to the isolation and purification of bioactive peptides. Chem. Eng. Res. Des. 2016, 98, 244–256. [Google Scholar] [CrossRef]
- Segura-Campos, M.; Chel-Guerrero, L.; Betancur-Ancona, D.; Hernandez-Escalante, V.M. Bioavailability of Bioactive Peptides. Food Rev. Int. 2011, 27, 213–226. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Liu, X.; Huang, L.; Chen, Z.; Cheng, J. Influence of hydrolysis behaviour and microfluidisation on the functionality and structural properties of collagen hydrolysates. Food Chem. 2017, 227, 211–218. [Google Scholar] [CrossRef]
- Halim, N.R.A.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Tavano, O.L. Protein hydrolysis using proteases: An important tool for food biotechnology. J. Mol. Catal. B Enzym. 2013, 90, 1–11. [Google Scholar] [CrossRef]
- Min, S.-G.; Jo, Y.-J.; Park, S.-H. Potential application of static hydrothermal processing to produce the protein hydrolysates from porcine skin by-products. LWT 2017, 83, 18–25. [Google Scholar] [CrossRef]
- Chang, O.K.; Ha, G.E.; Jeong, S.-G.; Seol, K.H.; Oh, M.H.; Kim, D.W.; Jang, A.; Kim, S.H.; Park, B.Y.; Ham, J.-S. Antioxidant activity of porcine skin gelatin hydrolyzed by pepsin and pancreatin. Korean J. Food Sci. Anim. Resour. 2013, 33, 493–500. [Google Scholar] [CrossRef]
- Ahn, C.-B.; Je, J.-Y.; Cho, Y.S. Antioxidant and anti-inflammatory peptide fraction from salmon byproduct protein hydrolysates by peptic hydrolysis. Food Res. Int. 2012, 49, 92–98. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; Barre, C.; Fitz, R.J. Enzymatic generation of whey protein hydrolysates under pH-controlled and non pH-controlled conditions: Impact on physicochemical and bioactive properties. Food Chem. 2016, 199, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 2007, 102, 1317–1327. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant and angiotensin I converting enzyme (ACE) inhibitory activities of date seed protein hydrolysates prepared using Alcalase, Flavourzyme and Thermolysin. J. Funct. Foods 2015, 18, 1125–1137. [Google Scholar] [CrossRef]
- Li, B.; Chen, F.; Wang, X.; Ji, B.; Wu, Y. Isolation and identification of antioxidative peptides from porcine collagen hydrolysate by consecutive chromatography and electrospray ionization–mass spectrometry. Food Chem. 2007, 102, 1135–1143. [Google Scholar] [CrossRef]
- Ojha, K.S.; Alvarez, C.; Kumar, P.; O’Donnell, C.P.; Tiwari, B.K. Effect of enzymatic hydrolysis on the production of free amino acids from boarfish (Capros aper) using second order polynomial regression models. LWT 2016, 68, 470–476. [Google Scholar] [CrossRef]
- Margot, A.; Flaschel, E.; Renken, A. Continuous monitoring of enzymatic whey protein hydrolysis. Correlation of base consumption with soluble nitrogen content. Process Biochem. 1994, 29, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Guillén, M.C.; Giménez, B.; López-Caballero, M.E.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.; Min, S.-G.; Jo, Y.-J. Functionality of porcine skin hydrolysates produced by hydrothermal processing for liposomal delivery system. J. Food Biochem. 2018, 42, e12464. [Google Scholar] [CrossRef]
- Khiari, Z.; Ndagijimana, M.; Betti, M. Low molecular weight bioactive peptides derived from the enzymatic hydrolysis of collagen after isoelectric solubilization/precipitation process of turkey by-products. Poult. Sci. 2014, 93, 2347–2362. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Yang, H.; Sun, Y.; Niu, J.; Liu, S. Purification and identification of antioxidant peptides from walnut (Juglans regia L.) protein hydrolysates. Peptides 2012, 38, 344–349. [Google Scholar] [CrossRef]
- Ao, J.; Li, B. Amino acid composition and antioxidant activities of hydrolysates and peptide fractions from porcine collagen. Food Sci. Technol. Int. 2012, 18, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Onuh, J.O.; Girgih, A.T.; Aluko, R.E.; Aliani, M. In vitro antioxidant properties of chicken skin enzymatic protein hydrolysates and membrane fractions. Food Chem. 2014, 150, 366–373. [Google Scholar] [CrossRef]
- Olagunju, A.I.; Omoba, O.S.; Enujiugha, V.N.; Alashi, A.M.; Aluko, R.E. Pigeon pea enzymatic protein hydrolysates and ultrafiltration peptide fractions as potential sources of antioxidant peptides: An in vitro study. LWT 2018, 97, 269–278. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Wu, K.-C.; Chiang, S.-H. Antioxidant properties and protein compositions of porcine haemoglobin hydrolysates. Food Chem. 2007, 100, 1537–1543. [Google Scholar] [CrossRef]
- Intarasirisawat, R.; Benjakul, S.; Visessanguan, W.; Wu, J. Antioxidative and functional properties of protein hydrolysate from defatted skipjack (Katsuwonous pelamis) roe. Food Chem. 2012, 135, 3039–3048. [Google Scholar] [CrossRef] [PubMed]
- Je, J.-Y.; Lee, K.-H.; Lee, M.H.; Ahn, C.-B. Antioxidant and antihypertensive protein hydrolysates produced from tuna liver by enzymatic hydrolysis. Food Res. Int. 2009, 42, 1266–1272. [Google Scholar] [CrossRef]
- Meir, S.; Kanner, J.; Akiri, B.; Philosoph-Hadas, S. Determination and involvement of aqueous reducing compounds in oxidative defense systems of various senescing leaves. J. Agric. Food Chem. 1995, 43, 1813–1819. [Google Scholar] [CrossRef]
- Xia, Y.; Bamdad, F.; Gänzle, M.; Chen, L. Fractionation and characterization of antioxidant peptides derived from barley glutelin by enzymatic hydrolysis. Food Chem. 2012, 134, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.-H.; Jung, E.Y.; Noh, D.O.; Suh, H.J. Physiological effects of formulation containing tannase-converted green tea extract on skin care: Physical stability, collagenase, elastase, and tyrosinase activities. Integr. Med. Res. 2014, 3, 25–33. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Solubility % | Free Amino Group % | Yield % 1 |
|---|---|---|---|
| Control | 11.95 ± 0.62 b,* | 0.79 ± 0.06 c | 27.27 |
| CH-Alcalase | 19.83 ± 0.04 a | 5.69 ± 0.02 b | 21.76 |
| CH-Alcalase/<3 kDa | 21.17 ± 0.15 a | 14.17 ± 0.25 a | 12.42 |
| Amino Acids | Control | CH-Alcalase | CH-Alcalase/<3 kDa |
|---|---|---|---|
| Aspartic acid | 34.53 | 43.09 | 63.74 |
| Glutamic acid | 62.21 | 78.38 | 120.51 |
| Asparagine | N.D. * | N.D. | N.D. |
| Serine | 19.75 | 24.42 | 35.56 |
| Glutamine | N.D. | N.D. | N.D. |
| Histidine | 5.30 | 6.03 | 8.05 |
| Glycine | 116.63 | 148.61 | 218.5 |
| Threonine | 10.50 | 13.83 | 19.32 |
| Arginine | 44.05 | 52.01 | 73.05 |
| Alanine | 46.90 | 59.02 | 91.17 |
| GABA | N.D. | N.D. | N.D. |
| Tyrosine | 5.96 | 6.49 | 9.30 |
| Valine | 16.22 | 19.73 | 29.85 |
| Methionine | 4.30 | 4.39 | 9.29 |
| Phenylalanine | 13.42 | 16.25 | 22.51 |
| Isoleucine | 9.00 | 10.69 | 14.76 |
| Leucine | 19.86 | 22.74 | 33.24 |
| Lysine | 18.09 | 21.44 | 29.90 |
| Proline | 91.14 | 95.09 | 152.85 |
| Treatments 1 | Tyrosinase Inhibition Activity (%) | Collagenase Inhibition Activity (%) | Elastase Inhibition Activity (%) |
|---|---|---|---|
| Ascorbic acid 2 | 95.50 ± 0.02 a,* | 48.09 ± 0.01 b,c | 27.00 ± 0.05 |
| Control | 28.2 ± 2.8 b | 6.45 ± 3.3 d | - |
| CH-Alcalase | 15.44 ± 0.01 c | 54.4 ± 9.0 b | - |
| CH-Alcalase/<3 kDa | 30.20 ± 0.05 b | 61.9 ± 2.6 a | - |
| Name | Type | Origin | Optimal Temperature (°C) | Optimal pH |
|---|---|---|---|---|
| Alcalase | endo (serine endoprotease) | Bacillus licheniformis | 60 | 8 |
| Flavorzyme | exo (aminopeptidase), endo mix | Aspergillus oryzae | 60 | 6.5 |
| Neutrase | endo (metallo-endoprotease) | Bacillus amyloliquefaciens | 45 | 8 |
| Bromeline | - | Pineapple | 60 | 7 |
| Protamex | endo (serine endoprotease) | Bacillus licheniformis Bacillus amyloliquefaciens | 60 | 7 |
| Papain | - | Carica papaya | 60 | 7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, G.-P.; Min, S.-G.; Jo, Y.-J. Anti-Oxidative and Anti-Aging Activities of Porcine By-Product Collagen Hydrolysates Produced by Commercial Proteases: Effect of Hydrolysis and Ultrafiltration. Molecules 2019, 24, 1104. https://doi.org/10.3390/molecules24061104
Hong G-P, Min S-G, Jo Y-J. Anti-Oxidative and Anti-Aging Activities of Porcine By-Product Collagen Hydrolysates Produced by Commercial Proteases: Effect of Hydrolysis and Ultrafiltration. Molecules. 2019; 24(6):1104. https://doi.org/10.3390/molecules24061104
Chicago/Turabian StyleHong, Geun-Pyo, Sang-Gi Min, and Yeon-Ji Jo. 2019. "Anti-Oxidative and Anti-Aging Activities of Porcine By-Product Collagen Hydrolysates Produced by Commercial Proteases: Effect of Hydrolysis and Ultrafiltration" Molecules 24, no. 6: 1104. https://doi.org/10.3390/molecules24061104
APA StyleHong, G. -P., Min, S. -G., & Jo, Y. -J. (2019). Anti-Oxidative and Anti-Aging Activities of Porcine By-Product Collagen Hydrolysates Produced by Commercial Proteases: Effect of Hydrolysis and Ultrafiltration. Molecules, 24(6), 1104. https://doi.org/10.3390/molecules24061104