Shedding New Light on The Role of ανβ3 and α5β1 Integrins in Rheumatoid Arthritis


Abstract
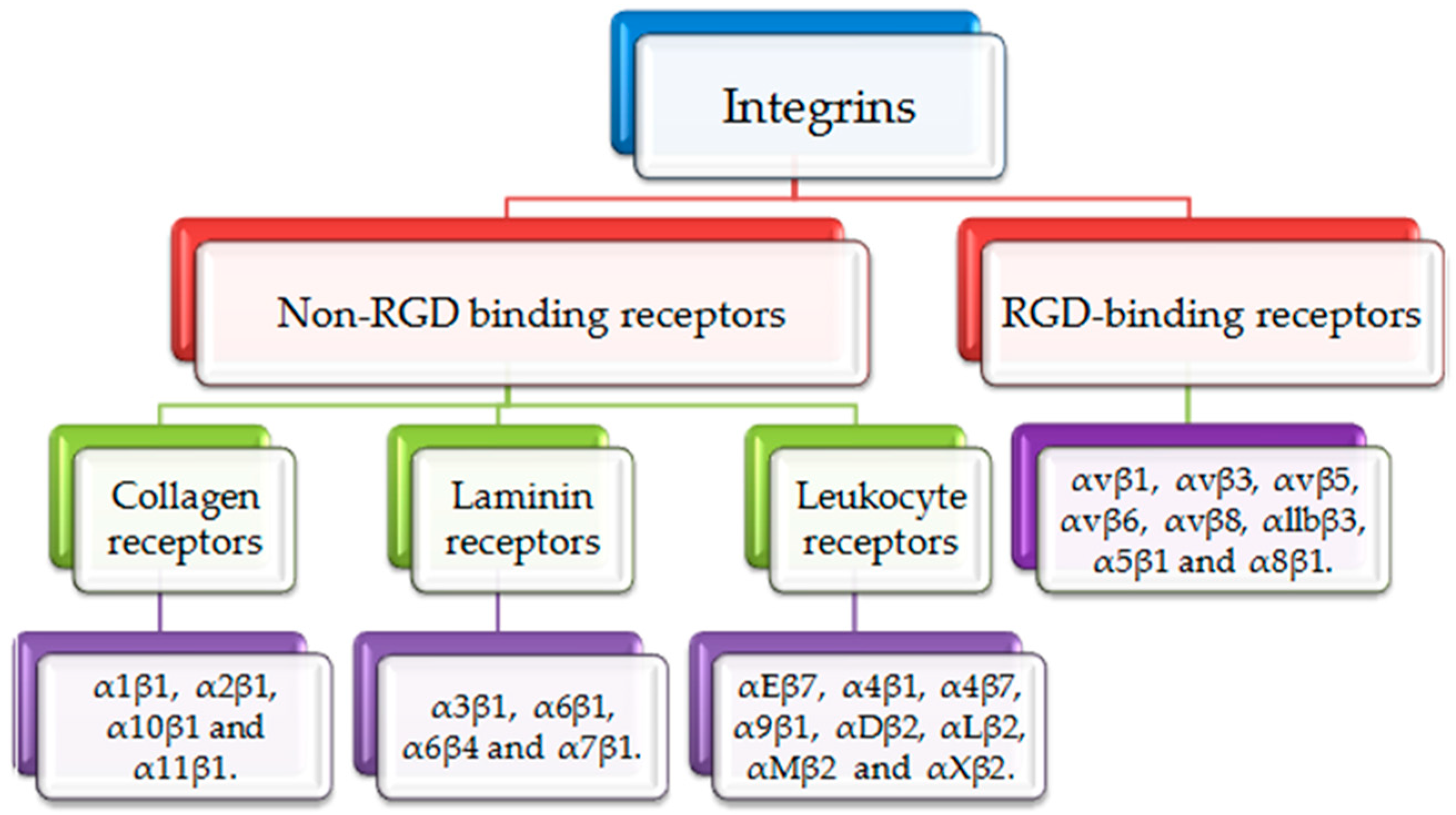
:1. Introduction
2. αvβ3 and α5β1 in RA Development
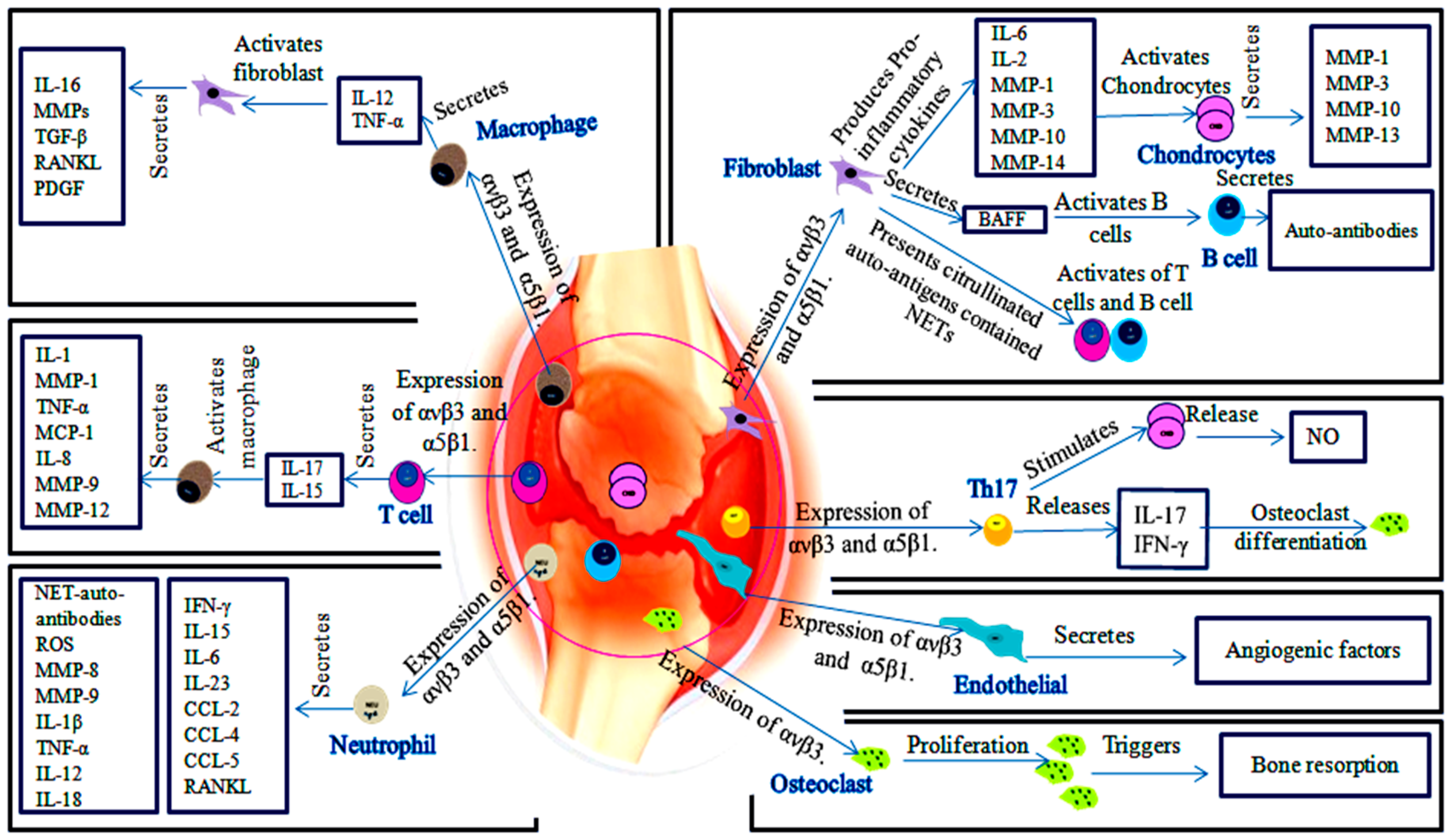
2.1. αvβ3 and α5β1 Facilitate ECM Protein-Rheumatoid Cells and Cell-Cell Communication
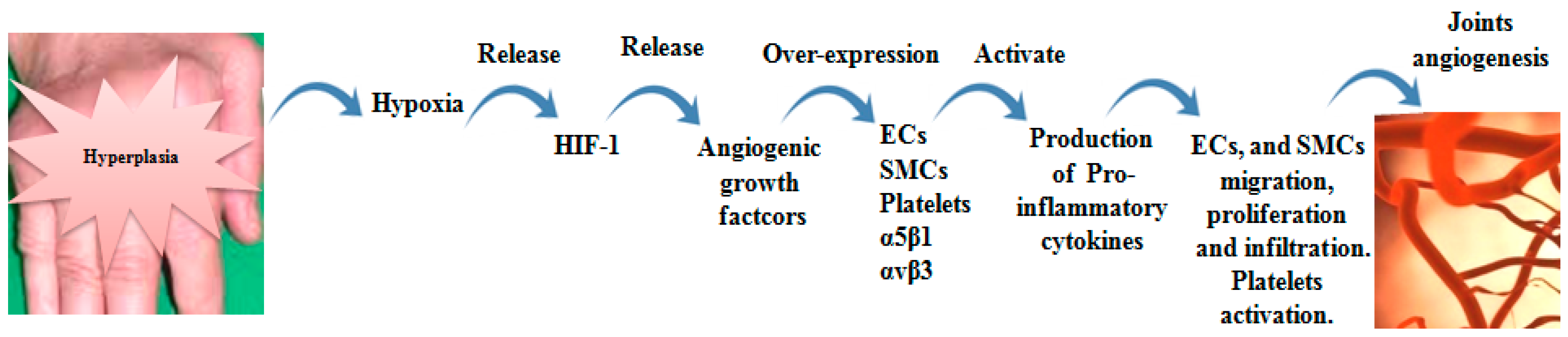
2.2. Angiogenesis
3. Targeting of αvβ3 and α5β1 Integrins as Crucial Rheumatoid Arthritis Therapies
3.1. Anti-αvβ3 Agents
3.2. Anti α5β1 Agents
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Emori, T.; Hirose, J.; Ise, K.; Yomoda, J.I.; Kasahara, M.; Shinkuma, T.; Yoshitomi, H.; Ito, H.; Hashimoto, M.; Sugahara, S. Constitutive Activation of Integrin α9 Augments Self-Directed Hyperplastic and Proinflammatory Properties of Fibroblast-like Synoviocytes of Rheumatoid Arthritis. J. Immunol. 2017, 199, 1700941. [Google Scholar] [CrossRef]
- Shim, J.-H.; Stavre, Z.; Gravallese, E.M. Bone Loss in Rheumatoid Arthritis: Basic Mechanisms and Clinical Implications. Calcif. Tissue Int. 2018, 102, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Dequattro, K.; Imboden, J.B. Neurologic Manifestations of Rheumatoid Arthritis. Rheum. Dis. Clin. North Am. 2017, 43, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, J.; Maslinski, W.; Pazdur, J.; Sliwinskastanczyk, P.; Kaminskatchorzewska, E.; Jung, L.; Lacki, J.K. Decreased expression of integrins by hematopoietic cells in patients with rheumatoid arthritis and anemia: Relationship with bone marrow cytokine levels. J. Invest. Allergol. Clin. Immunol. 2008, 18, 17. [Google Scholar]
- Put, S.; Westhovens, R.; Lahoutte, T.; Matthys, P. Molecular imaging of rheumatoid arthritis: Emerging markers, tools, and techniques. Arthritis Res. Ther. 2014, 16, 208. [Google Scholar] [CrossRef]
- Juanrivera, M.C.; Martinezferrer, M. Integrin Inhibitors in Prostate Cancer. Cancers 2018, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Bartok, B.; Firestein, G.S. Fibroblast-like synoviocytes: Key effector cells in rheumatoid arthritis. Immunol. Rev. 2010, 233, 233–255. [Google Scholar] [CrossRef] [PubMed]
- De Marco, R.; Tolomelli, A.; Juaristi, E.; Gentilucci, L. Integrin Ligands with α/β-Hybrid Peptide Structure: Design, Bioactivity, and Conformational Aspects. Med. Res. Rev. 2016, 36, 389–424. [Google Scholar] [CrossRef] [PubMed]
- Ansari, A.A.; Byrareddy, S.N. The Role of Integrin Expressing Cells in Modulating Disease Susceptibility and Progression (January 2016). Int. Trends Immun. 2016, 4, 11–27. [Google Scholar]
- Koivisto, L.; Heino, J.; Hakkinen, L.; Larjava, H. Integrins in Wound Healing. Adv. Wound Care 2014, 3, 762–783. [Google Scholar] [CrossRef] [Green Version]
- Finney, A.C.; Stokes, K.Y.; Pattillo, C.B.; Orr, A.W. Integrin signaling in atherosclerosis. Cell. Mol. Life Sci. 2017, 74, 2263–2282. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Zhang, K.; Chen, J. Role of Integrins in Regulating Proteases to Mediate Extracellular Matrix Remodeling. Cancer Microenviron. 2012, 5, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.K.; Patzak, I.; Svensson, L.; De Filippo, K.; Jones, K.; Mcdowall, A.; Hogg, N. Integrins in immunity. J. Cell Sci. 2009, 122, 215–225. [Google Scholar] [CrossRef]
- Balcioglu, H.E.; Van Hoorn, H.; Donato, D.M.; Schmidt, T.; Danen, E.H.J. The integrin expression profile modulates orientation and dynamics of force transmission at cell–matrix adhesions. J. Cell Sci. 2015, 128, 1316–1326. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.R.; Byron, A.; Humphries, M.J.; Bass, M.D. Giving off mixed signals—Distinct functions of α5β1 and αvβ3 integrins in regulating cell behaviour. Iubmb Life 2009, 61, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Perdih, A.; Dolenc, M.S. Small molecule antagonists of integrin receptors. Curr. Med. Chem. 2010, 17, 2371–2392. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Straub, R.H. Integrins and their ligands in rheumatoid arthritis. Arthritis Res. Ther. 2011, 13, 244. [Google Scholar] [CrossRef]
- Bustamante, M.F.; Garciacarbonell, R.; Whisenant, K.D.; Guma, M. Fibroblast-like synoviocyte metabolism in the pathogenesis of rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, G.; Moreno, C.; Montserrat, E. Comment on “Soluble BAFF levels inversely correlate with peripheral B cell numbers and the expression of BAFF receptors”. J. Immunol. 2012, 188, 2930–2931. [Google Scholar] [CrossRef] [PubMed]
- Wilder, R.L. Integrin alpha V beta 3 as a target for treatment of rheumatoid arthritis and related rheumatic diseases. Ann. Rheum. Dis. 2002, 61, 96–99. [Google Scholar] [CrossRef]
- Rocha, L.A.; Learmonth, D.A.; Sousa, R.A.; Salgado, A.J. αvβ3 and α5β1 integrin-specific ligands: From tumor angiogenesis inhibitors to vascularization promoters in regenerative medicine? Biotechnol. Adv. 2017, 36, 208–227. [Google Scholar] [CrossRef] [PubMed]
- Storgard, C.M.; Stupack, D.G.; Jonczyk, A.; Goodman, S.L.; Fox, R.I.; Cheresh, D.A. Decreased angiogenesis and arthritic disease in rabbits treated with an αvβ3 antagonist. J. Clin. Invest. 1999, 103, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badger, A.M.; Blake, S.M.; Kapadia, R.; Sarkar, S.K.; Levin, J.M.; Swift, B.A.; Hoffman, S.J.; Stroup, G.B.; Miller, W.H.; Gowen, M. Disease-modifying activity of SB 273005, an orally active, nonpeptide αvβ3 (vitronectin receptor) antagonist, in rat adjuvant-induced arthritis. Arthritis Rheum. 2001, 44, 128–137. [Google Scholar] [CrossRef]
- Mousa, S.A.; Davis, P.J. Integrin Antagonists and Angiogenesis. In Angiogenesis Modulations in Health and Disease: Practical Applications of Pro- and Anti-angiogenesis Targets; Mousa, S.A., Davis, P.J., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2013; pp. 119–141. [Google Scholar]
- Huang, R.; Li, J.; Wang, Y.; Zhang, L.; Ma, X.; Wang, H.; Li, W.; Cao, X.; Xu, H.; Hu, J. The Protective Effect of a Long-Acting and Multi-Target HM-3-Fc Fusion Protein in Rheumatoid Arthritis. Int. J. Mol. Sci. 2018, 19, 2683. [Google Scholar] [CrossRef] [PubMed]
- Attur, M.; Dave, M.N.; Clancy, R.R.; Patel, I.R.; Abramson, S.B.; Amin, A.R. Functional genomic analysis in arthritis-affected cartilage: Yin-yang regulation of inflammatory mediators by alpha 5 beta 1 and alpha V beta 3 integrins. J. Immunol. 2000, 164, 2684–2691. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.; Iommelli, F.; De Rosa, V.; Carriero, M.V.; Miceli, R.; Camerlingo, R.; Minno, G.D.; Vecchio, S.D. Integrin-dependent cell adhesion to neutrophil extracellular traps through engagement of fibronectin in neutrophil-like cells. PLOS ONE 2017, 12, e0171362. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F. Integrins and chondrocyte–matrix interactions in articular cartilage. Matrix Biol. 2014, 39, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Steenvoorden, M.M.C.; Bank, R.A.; Ronday, H.K.; Toes, R.E.M.; Huizinga, T.W.J.; Degroot, J. Fibroblast-like synoviocyte-chondrocyte interaction in cartilage degradation. Clin. Exp. Rheumatol. 2007, 25, 239–245. [Google Scholar]
- Otero, M.; Goldring, M.B. Cells of the synovium in rheumatoid arthritis. Chondrocytes. Arthritis Res. Ther. 2007, 9, 220. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, Q.; Ke, Y.; Lin, J. Neutrophil Function in an Inflammatory Milieu of Rheumatoid Arthritis. Clin. Dev. Immunol. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, C.B.; Pulai, J.I.; Loeser, R.F. Fibronectin fragments and blocking antibodies to α2β1 and α5β1 integrins stimulate mitogen-activated protein kinase signaling and increase collagenase 3 (matrix metalloproteinase 13) production by human articular chondrocytes. Arthritis Rheum. 2002, 46, 2368–2376. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y. Metalloproteinases in Rheumatoid Arthritis: Potential Therapeutic Targets to Improve Current Therapies. Prog. Mol. Biol. Transl. Sci. 2017, 148, 327–338. [Google Scholar] [PubMed]
- Van Hamburg, J.P.; Tas, S.W. Molecular mechanisms underpinning T helper 17 cell heterogeneity and functions in rheumatoid arthritis. J. Autoimmun. 2017, 87, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Kinne, R.W.; Brauer, R.; Stuhlmuller, B.; Palombokinne, E.; Burmester, G.R. Macrophages in rheumatoid arthritis. Arthritis Res. Ther. 2000, 2, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Bondeson, J. The Role of Synovial Macrophages in Rheumatoid Arthritis and Osteoarthritis: Its Implications for Radiosynovectomy. In Local Treatment of Inflammatory Joint Diseases: Benefits and Risks; Kampen, W.U., Fischer, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 31–48. [Google Scholar]
- Brilha, S.; Wysoczanski, R.; Whittington, A.M.; Friedland, J.S.; Porter, J.C. Monocyte Adhesion, Migration, and Extracellular Matrix Breakdown Is Regulated by Integrin αVβ3 in Mycobacterium tuberculosis Infection. J. Immunol. 2017, 199, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Bishop, G.G.; Mcpherson, J.A.; Sanders, J.M.; Hesselbacher, S.E.; Feldman, M.J.; Mcnamara, C.A.; Gimple, L.W.; Powers, E.R.; Mousa, S.A.; Sarembock, I.J. Selective αvβ3-Receptor Blockade Reduces Macrophage Infiltration and Restenosis After Balloon Angioplasty in the Atherosclerotic Rabbit. Circulation 2001, 103, 1906. [Google Scholar] [CrossRef]
- Nakamura, I.; Duong, L.T.; Rodan, S.B.; Rodan, G.A. Involvement of αvβ3 integrins in osteoclast function. J. Bone Miner. Metab. 2007, 25, 337–344. [Google Scholar] [CrossRef]
- Tanaka, S.; Nakamura, K.; Oda, H. The osteoclast: A potential therapeutic target of bone and joint destruction in rheumatoid arthritis. Mod. Rheumatol. 2001, 11, 177–183. [Google Scholar] [CrossRef]
- Cascao, R.; Rosario, H.S.; Soutocarneiro, M.M.; Fonseca, J.E. Neutrophils in rheumatoid arthritis: More than simple final effectors. Autoimmun. Rev. 2010, 9, 531–535. [Google Scholar] [CrossRef]
- Malemud, C.J. Matrix Metalloproteinases and Synovial Joint Pathology. Prog. Mol. Biol. Transl. Sci. 2017, 148, 305–325. [Google Scholar]
- Gizinski, A.M.; Fox, D.A. T cell subsets and their role in the pathogenesis of rheumatic disease. Curr. Opin. Rheumatol. 2014, 26, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Giancotti, F.G. Integrin signalling during tumour progression. Nature Reviews Molecular Cell Biology 2004, 5, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Teoh, C.M.; Tam, J.; Tran, T. Integrin and GPCR Crosstalk in the Regulation of ASM Contraction Signaling in Asthma. J. Allergy 2012, 2012, 341282. [Google Scholar] [CrossRef] [PubMed]
- Short, S.M.; Boyer, J.L.; Juliano, R.L. Integrins regulate the linkage between upstream and downstream events in G protein-coupled receptor signaling to mitogen-activated protein kinase. J. Biol. Chem. 2000, 275, 12970–12977. [Google Scholar] [CrossRef] [PubMed]
- Kramarenko, I.I.; Bunni, M.A.; Raymond, J.R.; Garnovskaya, M.N. Bradykinin B2 Receptor Interacts with Integrin α5β1 to Transactivate Epidermal Growth Factor Receptor in Kidney Cells. Mol. Pharmacol. 2010, 78, 126–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarbock, A.; Ley, K. Neutrophil Adhesion and Activation under Flow. Microcirculation 2009, 16, 31–42. [Google Scholar] [CrossRef]
- Antonio, J.D.S.; Zoeller, J.J.; Habursky, K.; Turner, K.; Pimtong, W.; Burrows, M.; Choi, S.; Basra, S.; Bennett, J.S.; Degrado, W.F. A Key Role for the Integrin α2β1 in Experimental and Developmental Angiogenesis. Am. J. Pathol. 2009, 175, 1338–1347. [Google Scholar] [CrossRef] [Green Version]
- Szekanecz, Z.; Besenyei, T.; Paragh, G.; Koch, A.E. Angiogenesis in rheumatoid arthritis. Autoimmunity 2009, 42, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Millard, M.; Odde, S.; Neamati, N. Integrin targeted therapeutics. Theranostics 2011, 1, 154–188. [Google Scholar] [CrossRef]
- Paleolog, E.M. Angiogenesis in rheumatoid arthritis. Arthritis Res. Ther. 2002, 4, 339–365. [Google Scholar]
- Margadant, C.; Sonnenberg, A. Integrin–TGF-β crosstalk in fibrosis, cancer and wound healing. EMBO Rep. 2010, 11, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Avraamides, C.J.; Garmysusini, B.; Varner, J.A. Integrins in angiogenesis and lymphangiogenesis. Nat. Rev. Cancer 2008, 8, 604–617. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Mythreye, K.; Golzio, C.; Katsanis, N.; Blobe, G.C. Endoglin mediates fibronectin/α5β1 integrin and TGF-β pathway crosstalk in endothelial cells. EMBO J. 2012, 31, 3885–3900. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Saba, T.M.; Tsan, M. Role of alpha(v)beta(3)-integrin in TNF-alpha-induced endothelial cell migration. Am. J. Physiol.-Cell Physiol. 2002, 283, C1196–C1205. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Kim, H. Osteoclasts: Crucial in Rheumatoid Arthritis. J. Reprod. Dev. 2016, 23, 141–147. [Google Scholar] [CrossRef]
- Simic, D.; Bogdan, N.; Teng, F.; Otieno, M.A. Blocking α5β1 Integrin Attenuates sCD40L-Mediated Platelet Activation. Clin. Appl. Throm.-Hemost. 2017, 23, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Harifi, G.; Sibilia, J. Pathogenic role of platelets in rheumatoid arthritis and systemic autoimmune diseases. Perspectives and therapeutic aspects. Saudi Med. J. 2016, 37, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.; Vijayan, K.V.; Rumbaut, R.E. Platelets and Their Interactions with Other Immune Cells. Compr. Physiol. 2015, 5, 1265–1280. [Google Scholar]
- Habets, K.L.L.; Trouw, L.A.; Levarht, E.W.N.; Korporaal, S.J.A.; Habets, P.A.M.; De Groot, P.; Huizinga, T.W.J.; Toes, R.E.M. Anti-citrullinated protein antibodies contribute to platelet activation in rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 209. [Google Scholar] [CrossRef]
- Veeverslowe, J.; Ball, S.G.; Shuttleworth, A.; Kielty, C.M. Mesenchymal stem cell migration is regulated by fibronectin through α5β1-integrin-mediated activation of PDGFR-β and potentiation of growth factor signals. J. Cell Sci. 2011, 124, 1288–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakshmikanthan, S.; Sobczak, M.; Chun, C.; Henschel, A.; Dargatz, J.; Ramchandran, R.; Chrzanowskawodnicka, M. Rap1 promotes VEGFR2 activation and angiogenesis by a mechanism involving integrin αvβ3. Blood 2011, 118, 2015–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchings, H.; Ortega, N.; Plouet, J. Extracellular matrix-bound vascular endothelial growth factor promotes endothelial cell adhesion, migration, and survival through integrin ligation. FASEB J. 2003, 17, 1520–1522. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Li, M.B.; Wu, X.; Wu, S.; Zhu, W.; Chen, D.; Luo, M.; Eitenmuller, I.; Kampmann, A.; Schaper, J. Activation of the integrins α5β1 and αvβ3 and focal adhesion kinase (FAK) during arteriogenesis. Mol. Cell. Biochem. 2009, 322, 161–169. [Google Scholar] [CrossRef]
- Marrelli, A.; Cipriani, P.; Liakouli, V.; Carubbi, F.; Perricone, C.; Perricone, R.; Giacomelli, R. Angiogenesis in rheumatoid arthritis: A disease specific process or a common response to chronic inflammation? Autoimmun. Rev. 2011, 10, 595–598. [Google Scholar] [CrossRef]
- Ray, A.; Schaffner, F.; Janouskova, H.; Noulet, F.; Rognan, D.; Lelongrebel, I.; Choulier, L.; Blandin, A.; Lehmann, M.; Martin, S. Single cell tracking assay reveals an opposite effect of selective small non-peptidic α5β1 or αvβ3/β5 integrin antagonists in U87MG glioma cells. Biochim. Biophys. Acta 2014, 1840, 2978–2987. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, F.; Chen, X. Integrin αvβ3-targeted cancer therapy. Drug Dev. Res. 2008, 69, 329–339. [Google Scholar] [CrossRef]
- Schaffner, F.; Ray, A.M.; Dontenwill, M. Integrin α5β1, the Fibronectin Receptor, as a Pertinent Therapeutic Target in Solid Tumors. Cancers 2013, 5, 27–47. [Google Scholar] [CrossRef]
- Goodman, S.L.; Picard, M. Integrins as therapeutic targets. Trends Pharmacol. Sci. 2012, 33, 405–412. [Google Scholar] [CrossRef]
- Hatley, R.J.; Macdonald, S.J.F.; Slack, R.J.; Le, J.; Ludbrook, S.B.; Lukey, P.T. An αv-RGD Integrin Inhibitor Toolbox: Drug Discovery Insight, Challenges and Opportunities. Angew. Chem. 2018, 57, 3298–3321. [Google Scholar] [CrossRef]
- Ferrari, M.; Onuoha, S.C.; Pitzalis, C. Going with the flow: Harnessing the power of the vasculature for targeted therapy in rheumatoid arthritis. Drug Discovery Today 2016, 21, 172–179. [Google Scholar] [CrossRef]
- Gutheil, J.; Campbell, T.N.; Pierce, P.R.; Watkins, J.D.; Huse, W.D.; Bodkin, D.J.; Cheresh, D.A. Targeted Antiangiogenic Therapy for Cancer Using Vitaxin: A Humanized Monoclonal Antibody to the Integrin ανβ3. Clin. Cancer Res. 2000, 6, 3056–3061. [Google Scholar]
- Szekanecz, Z.; Koch, A.E. Angiogenesis and its targeting in rheumatoid arthritis. Vasc. Pharmacol. 2009, 51, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lainercarr, D.; Brahn, E. Angiogenesis inhibition as a therapeutic approach for inflammatory synovitis. Nat. Rev. Rheumatol. 2007, 3, 434–442. [Google Scholar] [CrossRef]
- Kobayashi, M.; Sawada, K.; Kimura, T. Potential of Integrin Inhibitors for Treating Ovarian Cancer: A Literature Review. Cancers 2017, 9, 83. [Google Scholar] [CrossRef]
- Pandolfi, F.; Franza, L.; Altamura, S.; Mandolini, C.; Cianci, R.; Ansari, A.A.; Kurnick, J.T. Integrins: Integrating the Biology and Therapy of Cell–cell Interactions. Clin. Ther. 2017, 39, 2420–2436. [Google Scholar] [CrossRef]
- Heidenreich, A.; de Boer, C.J.; Schrijvers, D.; Rawal, S.K.; Szkarlat, K.; Bogdanova, N.; Dirix, L.; Stenzl, A.; Welslau, M.; Wang, G.; et al. A randomized, double-blind, multicenter, phase 2 study of a human monoclonal antibody to human αν integrins (intetumumab) in combination with docetaxel and prednisone for the first-line treatment of patients with metastatic castration-resistant prostate cancer. Ann. Oncol. 2012, 24, 329–336. [Google Scholar] [Green Version]
- Investigators, E. Platelet glycoprotein IIb/IIIa receptor blockade and low-dose heparin during percutaneous coronary revascularization. New Engl. J. Med. 1997, 336, 1689–1696. [Google Scholar]
- Chilla, A.; Bianconi, D.; Geetha, N.; Dorda, A.; Poettler, M.; Unseld, M.; Sykoutri, D.; Redlich, K.; Zielinski, C.C.; Prager, G.W. Effects of cilengitide in osteoclast maturation and behavior. Exp. Cell Res. 2015, 337, 68–75. [Google Scholar] [CrossRef]
- Chinot, O. Cilengitide in glioblastoma: When did it fail? Lancet Oncol. 2014, 15, 1044–1045. [Google Scholar] [CrossRef]
- Murphy, M.G.; Cerchio, K.; Stoch, S.A.; Gottesdiener, K.M.; Wu, M.; Recker, R.R. Effect of L-000845704, an αVβ3 Integrin Antagonist, on Markers of Bone Turnover and Bone Mineral Density in Postmenopausal Osteoporotic Women. J. Clin. Endocrinol. Metab. 2005, 90, 2022–2028. [Google Scholar] [CrossRef]
- Kumar, C.C.; Malkowski, M.; Yin, Z.; Tanghetti, E.; Yaremko, B.; Nechuta, T.; Varner, J.; Liu, M.; Smith, E.M.; Neustadt, B. Inhibition of Angiogenesis and Tumor Growth by SCH221153, a Dual αvβ3 and αvβ5 Integrin Receptor Antagonist. Cancer Res. 2001, 61, 2232–2238. [Google Scholar]
- Cirkel, G.A.; Kerklaan, B.M.; Vanhoutte, F.; Der Aa, A.V.; Lorenzon, G.; Namour, F.; Pujuguet, P.; Darquenne, S.; De Vos, F.Y.F.L.; Snijders, T.J. A dose escalating phase I study of GLPG0187, a broad spectrum integrin receptor antagonist, in adult patients with progressive high-grade glioma and other advanced solid malignancies. Invest. New Drugs 2016, 34, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, K.J.; Hurrell, J.E.; Cecchini, M.G.; Der Pluijm, G.V.; Down, J.; Eaton, C.L.; Hamdy, F.C.; Clementlacroix, P.; Brown, N.J. Prostate cancer cells home to bone using a novel in vivo model: Modulation by the integrin antagonist GLPG0187. Int. J. Cancer 2015, 136, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Lorenzon, G.; Gheyle, L.; Vets, E.; Namour, F.; Pujuguet, P.; Clementlacroix, P.; Wigerinck, P.; Vanhoutte, F. Abstract 1568: GLPG0187, a small molecule integrin antagonist, shows good safety and decrease in CTX levels in single ascending dose study. Cancer Res. 2010, 70, 1568. [Google Scholar] [CrossRef]
- Goswami, R.K.; Liu, Y.; Liu, C.; Lerner, R.A.; Sinha, S.C. Synthesis and evaluation of the aldolase antibody-derived chemical-antibodies targeting α5β1 integrin. Mol. Pharmaceutics 2013, 10, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Bellmcguinn, K.M.; Matthews, C.M.; Ho, S.; Barve, M.; Gilbert, L.; Penson, R.T.; Lengyel, E.; Palaparthy, R.; Gilder, K.; Vassos, A. A phase II, single-arm study of the anti-α5β1 integrin antibody volociximab as monotherapy in patients with platinum-resistant advanced epithelial ovarian or primary peritoneal cancer. Gynecol. Oncol. 2011, 121, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Mateo, J.; Berlin, J.; De Bono, J.S.; Cohen, R.B.; Keedy, V.L.; Mugundu, G.; Zhang, L.; Abbattista, A.; Davis, C.; Stampino, C.G. A first-in-human study of the anti-α5β1 integrin monoclonal antibody PF-04605412 administered intravenously to patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2014, 74, 1039–1046. [Google Scholar] [CrossRef] [Green Version]
- Zahn, G.; Vossmeyer, D.; Stragies, R.; Wills, M.; Wong, C.G.; Löffler, K.U.; Adamis, A.P.; Knolle, J. Preclinical Evaluation of the Novel Small-Molecule Integrin α5β1 Inhibitor JSM6427 in Monkey and Rabbit Models of Choroidal NeovascularizationJSM6427 for Choroidal Neovascularization. Arch. Ophthalmol. 2009, 127, 1329–1335. [Google Scholar] [CrossRef]
- Khalili, P.; Arakelian, A.; Chen, G.; Plunkett, M.L.; Beck, I.; Parry, G.; Donate, F.; Shaw, D.E.; Mazar, A.P.; Rabbani, S.A. A non–RGD-based integrin binding peptide (ATN-161) blocks breast cancer growth and metastasis in vivo. Mol. Cancer Ther. 2006, 5, 2271–2280. [Google Scholar] [CrossRef]
- Wang, W.; Wang, F.; Lu, F.; Xu, S.; Hu, W.; Huang, J.; Gu, Q.; Sun, X. The Antiangiogenic Effects of Integrin α5β1 Inhibitor (ATN-161) In Vitro and In Vivo. Invest. Ophthalmol. Visual Sci. 2011, 52, 7213–7220. [Google Scholar] [CrossRef] [Green Version]
- Sökeland, G.; Schumacher, U. The functional role of integrins during intra- and extravasation within the metastatic cascade. Mol. Cancer 2019, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Dai, J.; Yao, Z.; Shelley, G.; Keller, E.T. Abituzumab Targeting of αV-Class Integrins Inhibits Prostate Cancer Progression. Mol. Cancer Res. 2017, 15, 875–883. [Google Scholar] [CrossRef]
- Li, G.; Zhang, L.; Chen, E.; Wang, J.; Jiang, X.; Chen, J.H.; Wickman, G.R.; Amundson, K.K.; Bergqvist, S.; Zobel, J. Dual Functional Monoclonal Antibody PF-04605412 Targets Integrin α5β1 and Elicits Potent Antibody-Dependent Cellular Cytotoxicity. Cancer Res. 2010, 70, 10243–10254. [Google Scholar] [CrossRef] [PubMed]
- Takagi, J.; Strokovich, K.; Springer, T.A.; Walz, T. Structure of integrin α5β1 in complex with fibronectin. EMBO J. 2003, 22, 4607–4615. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Target | Antagonist Name | Antagonist Type | Effect on Cells Response (Functions) | Clinical Trials Phase | Ref. |
|---|---|---|---|---|---|
| αvβ3 | Etaracizumab | Engineered mAb | Inhibited cellular migration, adhesion and proliferation. Induced blood vessels apoptosis. Anti-angiogenic activity via blocking FGF-2 and TNF-α. | Phase II for RA, solid tumors, lymphoma and psoriasis. | [71,73,77] |
| Intetumumab (CNTO95) | mAb | Inhibited HUVECs migration and adhesion of melanoma cells. | Phase II for solid tumors. | [51,71,78] | |
| DI17E6 | mAb | Inhibited HUVECs migration and adhesion of melanoma cells. Suppressed development of prostate cancer. | Phase I for solid tumors. | [70,71,93,94] | |
| Abciximab (c7E3) | Chimeric mAb | Inhibited platelet aggregation by binding to αvβ3 and αllbβ3. Anti-tumor activity. | Approved for cancer therapy. | [51,71,79] | |
| Cilengitide (EMD121974) | RGD-peptide | Attenuated endothelial cells and tumor cells proliferation and migration by inhibiting the FAK/Src/AKT and Erk pathway. Induced apoptosis in endothelial cells. | Failure in phase III for cancer. | [71,80,81] | |
| L000845704 (MK-0429) | Small molecule | Inhibited bone resorption. | Phase I for osteoporosis and prostate cancer. | [71,82] | |
| SB273005 | Small molecule | Inhibited endothelial cells migration and bone loss. | Failure in phase I for osteoporosis. | [23,71] | |
| SCH221153 | RGD-peptide mimetic | Inhibited endothelial cells disorders and FGF-2 inhibitor. | - | [71,83] | |
| GLPG-0187 | Small molecule | Anti-angiogenic. Anti-tumor. Anti-bone resorption. | Phase I for solid tumors. | [70,71,85,86] | |
| HM-3 | RGD-peptide | Inhibited inflammatory factors, VEGF and PDGF-A in endothelial cells. | Phase I for cancer. | [25] | |
| α5β1 | Volociximab | Chimeric mAb | Induced in vivo and in vitro endothelial apoptosis. Prevented blood vessels formation. | Phase II for cancer. | [51,88] |
| PF-04605412 | mAb | Exhibited anti-angiogenesis and anti-tumor properties. | Phase I for cancer. | [93,95] | |
| JSM6427 | Small molecule | Induced anti-proliferative of endothelial cells activity. Showed an inhibition of choroidal neovascularization. | Phase I for age-related macular degeneration (AMD). | [90] | |
| ATN-161 | Non-RGD peptide | Blunted macrophage activation. Inhibited CAM expression. Exhibited anti-angiogenic properties. | Phase II for renal cancer. | [70,92] | |
| HM-3 | RGD-peptide | Inhibited inflammatory factors, VEGF and PDGF-A in endothelial cells. | Phase I for cancer. | [25] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morshed, A.; Abbas, A.B.; Hu, J.; Xu, H. Shedding New Light on The Role of ανβ3 and α5β1 Integrins in Rheumatoid Arthritis. Molecules 2019, 24, 1537. https://doi.org/10.3390/molecules24081537
Morshed A, Abbas AB, Hu J, Xu H. Shedding New Light on The Role of ανβ3 and α5β1 Integrins in Rheumatoid Arthritis. Molecules. 2019; 24(8):1537. https://doi.org/10.3390/molecules24081537
Chicago/Turabian StyleMorshed, Arwa, Abdul Baset Abbas, Jialiang Hu, and Hanmei Xu. 2019. "Shedding New Light on The Role of ανβ3 and α5β1 Integrins in Rheumatoid Arthritis" Molecules 24, no. 8: 1537. https://doi.org/10.3390/molecules24081537
APA StyleMorshed, A., Abbas, A. B., Hu, J., & Xu, H. (2019). Shedding New Light on The Role of ανβ3 and α5β1 Integrins in Rheumatoid Arthritis. Molecules, 24(8), 1537. https://doi.org/10.3390/molecules24081537