Review on the Biological Detoxification of Mycotoxins Using Lactic Acid Bacteria to Enhance the Sustainability of Foods Supply


Abstract
:1. Introduction
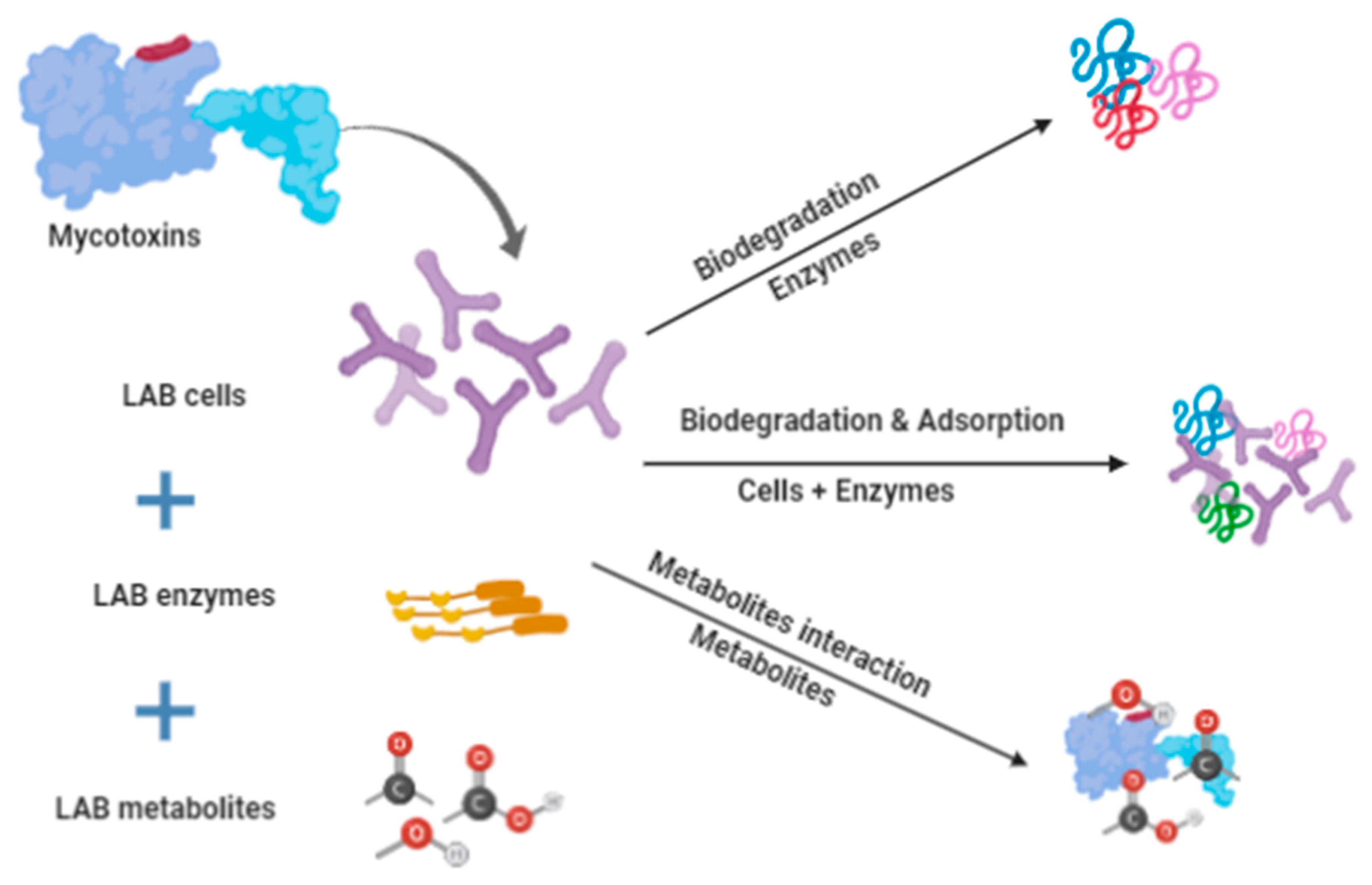
2. Biological Detoxification
3. Lactic Acid Bacteria Detoxification Activity
3.1. Aflatoxins Degradation by LAB
3.2. Ochratoxin A Degradation by LAB
3.3. Patulin Degradation by LAB
4. Application of LAB in Foods
5. Conclusions and Future Study
Author Contributions
Funding
Conflicts of Interest
References and Note
- Kokkonen, M.; Jestoi, M.; Rizzo, A. The effect of substrate on mycotoxin production of selected Penicillium strains. Int. J. Food Microbiol. 2005, 99, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarc’h, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. 2011, 28, 1590–1609. [Google Scholar] [CrossRef] [PubMed]
- da Rocha, M.E.B.; Freire, F.D.C.O.; Maia, F.E.F.; Guedes, M.I.F.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control. 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Dussort, P. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.; Zhou, T.; Yin, Y.; Xie, M.; Ruan, Z.; Young, J. Microbial strategies to control aflatoxins in food and feed. World Mycotoxin J. 2011, 4, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Sforza, S.; Dall’Asta, C.; Marchelli, R. Recent advances in mycotoxin determination in food and feed by hyphenated chromatographic techniques/mass spectrometry. Mass Spectrom. Rev. 2006, 25, 54–76. [Google Scholar] [CrossRef] [PubMed]
- Jelinek, C.F.; Pohland, A.E.; Wood, G.E. Worldwide occurrence of mycotoxins in foods and feeds--an update. J. Assoc. Off. Anal. Chem. 1989, 72, 223–230. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.; O’Kiely, P.; Forristal, P.D.; Fuller, H.T. Quantification and identification of fungal propagules in well-managed baled grass silage and in normal on-farm produced bales. Anim. Feed. Sci. Technol. 2007, 132, 283–297. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojuri, O.T.; Ezekiel, C.N.; Sulyok, M.; Ezeokoli, O.T.; Oyedele, O.A.; Ayeni, K.I.; Nwangburuka, C.C. Assessing the mycotoxicological risk from consumption of complementary foods by infants and young children in Nigeria. Food Chem. Toxicol. 2018, 121, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Rao, G.J.; Govindaraju, G.; Sivasithamparam, N.; Shanmugasundaram, E.R.B. Uptake, translocation and persistence of mycotoxins in rice seedlings. Plant. Soil 1982, 66, 121–123. [Google Scholar] [CrossRef]
- Stadler, D.; Berthiller, F.; Suman, M.; Schuhmacher, R.; Krska, R. Novel analytical methods to study the fate of mycotoxins during thermal food processing. Anal. Bioanal. Chem. 2020, 412, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadler, D.; Lambertini, F.; Bueschl, C.; Wiesenberger, G.; Hametner, C.; Schwartz-Zimmermann, H.; Suman, M. Untargeted LC–MS based 13C labelling provides a full mass balance of deoxynivalenol and its degradation products formed during baking of crackers, biscuits and bread. Food Chem. 2019, 279, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [Green Version]
- Richardson, K.E.; Hamilton, P.B. Enhanced production of pancreatic digestive enzymes during aflatoxicosis in egg-type chickens. Poult. Sci. 1987, 66, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Loureiro-Bracarense, A.P.; Lucioli, J.; Pacheco, G.D.; Cossalter, A.M.; Moll, W.D.; Oswald, I.P. Individual and combined effects of subclinical doses of deoxynivalenol and fumonisins in piglets. Mol. Nutr. Food Res. 2011, 55, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Martel, A.; Pasmans, F.; Ducatelle, R.; Verbrugghe, E.; Vandenbroucke, V.; Croubels, S. The impact of Fusarium mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 2014, 6, 430–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RASFF. The Rapid Alert System for Food and Feed-2011 Annual Report. 2011.
- Moretti, A.; Logrieco, A.F.; Susca, A. Mycotoxins: An underhand food problem. In Mycotoxigenic Fungi; Humana Press: New York, NY, USA, 2017; pp. 3–12. [Google Scholar]
- Ashiq, S. Natural occurrence of mycotoxins in food and feed: Pakistan perspective. Compr. Rev. Food Sci. Food Saf. 2015, 14, 159–175. [Google Scholar] [CrossRef]
- Kensler, T.W.; Roebuck, B.D.; Wogan, G.N.; Groopman, J.D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicol. Sci. 2011, 120, S28–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rheeder, J.P.; Marasas, W.F.; Vismer, H.F. Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattanzio, V.M.; Pascale, M. Determination of T-2 and HT-2 Toxins in Oats and Oat-Based Breakfast Cereals by Liquid-Chromatography Tandem Mass Spectrometry. In Oat; Humana Press: New York, NY, USA, 2017; pp. 127–136. [Google Scholar]
- Larsen, T.O.; Svendsen, A.; Smedsgaard, J. Biochemical characterization of ochratoxin A-producing strains of the genus Penicillium. Appl. Environ. Microbiol. 2001, 67, 3630–3635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, S.S. (Ed.) Toxic metals, radionuclides, and food packaging contaminants. In Hand Book of Food Toxicology; Marcel Dekker, Inc.: New York, NY, USA, 2002; pp. 783–810. [Google Scholar]
- Murugesan, G.R.; Ledoux, D.R.; Naehrer, K.; Berthiller, F.; Applegate, T.J.; Grenier, B.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poult. Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef] [PubMed]
- Tihomir, K.; Ivana, B.; Marija, K.; Ante, L.; Frane, Č.K.; Aleksandar, D.; Bojan, Š. Impact of fullerol C 60 (OH) 24 nanoparticles on the production of emerging toxins by Aspergillus flavus. Sci. Rep. 2020, 10. [Google Scholar]
- Kovač, T.; Borišev, I.; Crevar, B.; Kenjerić, F.Č.; Kovač, M.; Strelec, I.; Šarkanj, B. Fullerol C 60 (OH) 24 nanoparticles modulate aflatoxin B 1 biosynthesis in Aspergillus flavus. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Tóth, B. Novel strategies to control mycotoxins in feeds: A review. Acta Vet. Hung. 2005, 53, 189–203. [Google Scholar] [CrossRef] [PubMed]
- Karlovsky, P. Biological detoxification of fungal toxins and its use in plant breeding, feed and food production. Nat. Toxins 1999, 7, 1–23. [Google Scholar] [CrossRef]
- Ji, C.; Fan, Y.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Nichea, M.; Palacios, S.; Chiacchiera, S.; Sulyok, M.; Krska, R.; Chulze, S.; Ramirez, M. Presence of multiple mycotoxins and other fungal metabolites in native grasses from a wetland ecosystem in Argentina intended for grazing cattle. Toxins 2015, 7, 3309–3329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberts, J.F.; Gelderblom, W.C.A.; Botha, A.; Van Zyl, W.H. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef]
- Zhang, W.; Xue, B.; Li, M.; Mu, Y.; Chen, Z.; Li, J.; Shan, A. Screening a strain of Aspergillus niger and optimization of fermentation conditions for degradation of aflatoxin B1. Toxins 2014, 6, 3157–3172. [Google Scholar] [CrossRef] [PubMed]
- Sato, I.; Ito, M.; Ishizaka, M.; Ikunaga, Y.; Sato, Y.; Yoshida, S.; Tsushima, S. Thirteen novel deoxynivalenol-degrading bacteria are classified within two genera with distinct degradation mechanisms. FEMS Microbiol. Lett. 2012, 327, 110–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smiley, R.D.; Draughon, F.A. Preliminary evidence that degradation of aflatoxin B1 by Flavobacterium aurantiacum is enzymatic. J. Food Prot. 2000, 63, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Sangare, L.; Zhao, Y.; Folly, Y.; Chang, J.; Li, J.; Selvaraj, J.; Liu, Y. Aflatoxin B1 degradation by a Pseudomonas strain. Toxins 2014, 6, 3028–3040. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, M.; Shi, Z.Q.; Ghassempour, A.; Sedaghat, N.; Ahmadzadeh, M.; Mirabolfathy, M.; Javan-Nikkhah, M. Aflatoxin B1 degradation by Bacillus subtilis UTBSP1 isolated from pistachio nuts of Iran. Food Control. 2012, 23, 100–106. [Google Scholar] [CrossRef]
- He, J.W.; Bondy, G.S.; Zhou, T.; Caldwell, D.; Boland, G.J.; Scott, P.M. Toxicology of 3-epi-deoxynivalenol, a deoxynivalenol-transformation product by Devosia mutans 17–2-E-8. Food Chem. Toxicol. 2015, 84, 250–259. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Streit, E. Global occurrence of mycotoxins in the food and feed chain: Facts and toxins. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Ahad, R.; Zhou, T.; Lepp, D.; Pauls, K.P. Microbial detoxification of eleven food and feed contaminating trichothecene mycotoxins. BMC Biotechnol. 2017, 17, 30. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, Z.; Yuan, Y.; Cai, R.; Niu, C.; Yue, T. Identification of key factors involved in the biosorption of patulin by inactivated lactic acid bacteria (LAB) cells. PloS ONE 2015, 10, e0143431. [Google Scholar] [CrossRef]
- Shima, J.; Takase, S.; Takahashi, Y.; Iwai, Y.; Fujimoto, H.; Yamazaki, M.; Ochi, K. Novel detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated by enrichment culture. Appl. Environ. Microbiol. 1997, 63, 3825–3830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, E.M.; Binder, J. Method of using a feedstuff additive. U.S. Patent No. 7,347,997, 25 March 2008. [Google Scholar]
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1–7. [Google Scholar] [CrossRef]
- Gerez, C.L.; Torino, M.I.; Rollán, G.; de Valdez, G.F. Prevention of bread mould spoilage by using lactic acid bacteria with antifungal properties. Food Control. 2009, 20, 144–148. [Google Scholar] [CrossRef]
- Juodeikiene, G.; Salomskiene, J.; Eidukonyte, D.; Vidmantiene, D.; Narbutaite, V.; Vaiciulyte-Funk, L. The impact of novel fermented products containing extruded wheat material on the quality of wheat bread. Food Technol. Biotechnol. 2011, 49, 502–510. [Google Scholar]
- Abrunhosa, L.; Paterson, R.R.; Venâncio, A. Biodegradation of ochratoxin A for food and feed decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Nezami, H.; Kankaanpaa, P.; Salminen, S.; Ahokas, J. Ability of dairy strains of lactic acid bacteria to bind a common food carcinogen, aflatoxin B1. Food Chem. Toxicol. 1998, 36, 321–326. [Google Scholar] [CrossRef]
- Peltonen, K.; El-Nezami, H.; Haskard, C.; Ahokas, J.; Salminen, S. Aflatoxin B1 binding by dairy strains of lactic acid bacteria and bifidobacteria. J. Dairy Sci. 2001, 84, 2152–2156. [Google Scholar] [CrossRef]
- Niderkorn, V.; Morgavi, D.P.; Aboab, B.; Lemaire, M.; Boudra, H. Cell wall component and mycotoxin moieties involved in the binding of fumonisin B1 and B2 by lactic acid bacteria. J. Appl. Microbiol. 2009, 106, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Sezer, Ç.; Güven, A.; Oral, N.B.; Vatansever, L. Detoxification of aflatoxin B_1 by bacteriocins and bacteriocinogenic lactic acid bacteria. Turk. J. Vet. Anim. Sci. 2013, 37, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Elsanhoty, R.M.; Salam, S.A.; Ramadan, M.F.; Badr, F.H. Detoxification of aflatoxin M1 in yoghurt using probiotics and lactic acid bacteria. Food Control. 2014, 43, 129–134. [Google Scholar] [CrossRef]
- Hernandez-Mendoza, A.; Garcia, H.S.; Steele, J.L. Screening of Lactobacillus casei strains for their ability to bind aflatoxin B1. Food Chem. Toxicol. 2009, 47, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Śliżewska, K.; Smulikowska, S. Detoxification of aflatoxin B1 and change in microflora pattern by probiotic in vitro fermentation of broiler feed. J. Anim. Feed Sci. 2011, 20, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Duan, C.; Zhao, Y.; Gao, L.; Niu, C.; Xu, J.; Li, S. Reduction of aflatoxin B1 toxicity by Lactobacillus plantarum C88: A potential probiotic strain isolated from Chinese traditional fermented food “tofu”. PLoS ONE 2017, 12, e0170109. [Google Scholar] [CrossRef]
- Biernasiak, J.; Piotrowska, M.; Libudzisz, Z. Detoxification of mycotoxins by probiotic preparation for broiler chickens. Mycotoxin Res. 2006, 22, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, S.; Sontag, G.; Stidl, R.; Ehrlich, V.; Kundi, M.; Knasmüller, S. Detoxification of patulin and ochratoxin A, two abundant mycotoxins, by lactic acid bacteria. Food Chem. Toxicol. 2008, 46, 1398–1407. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Inês, A.; Rodrigues, A.I.; Guimarães, A.; Pereira, V.L.; Parpot, P.; Venâncio, A. Biodegradation of ochratoxin A by Pediococcus parvulus isolated from Douro wines. Int. J. Food Microbiol. 2014, 188, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luz, C.; Ferrer, J.; Mañes, J.; Meca, G. Toxicity reduction of ochratoxin A by lactic acid bacteria. Food Chem. Toxicol. 2018, 112, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yue, T.; Yuan, Y.; Wang, Z.; Ye, M.; Cai, R. A new insight into the adsorption mechanism of patulin by the heat-inactive lactic acid bacteria cells. Food Control. 2015, 50, 104–110. [Google Scholar] [CrossRef]
- Franco, T.S.; Garcia, S.; Hirooka, E.Y.; Ono, Y.S.; Dos Santos, J.S. Lactic acid bacteria in the inhibition of Fusarium graminearum and deoxynivalenol detoxification. J. Appl. Microbiol. 2011, 111, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Niderkorn, V.; Boudra, H.; Morgavi, D.P. Binding of Fusarium mycotoxins by fermentative bacteria in vitro. J. Appl. Microbiol. 2006, 101, 849–856. [Google Scholar] [CrossRef]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. Reduction of fumonisin B1 and zearalenone by lactic acid bacteria in fermented maize meal. J. Food Prot. 2005, 68, 2095–2099. [Google Scholar] [CrossRef]
- Čvek, D.; Markov, K.; Frece, J.; Friganović, M.; Duraković, L.; Delaš, F. Adhesion of zearalenone to the surface of lactic acid bacteria cells. Croat. J. Food Technol. Biotechnol. Nutr. 2012, 7, 49–52. [Google Scholar]
- Rogowska, A.; Pomastowski, P.; Walczak, J.; Railean-Plugaru, V.; Rudnicka, J.; Buszewski, B. Investigation of Zearalenone Adsorption and Biotransformation by Microorganisms Cultured under Cellular Stress Conditions. Toxins 2019, 11, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapot-Chartier, M.P.; Kulakauskas, S. Cell wall structure and function in lactic acid bacteria. Microb. Cell Fact. 2014, 13, S9. [Google Scholar]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria–Potential for control of mould growth and mycotoxins: A review. Food Control. 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Gomaa, E.Z.; Abdelall, M.F.; El-Mahdy, O.M. Detoxification of aflatoxin B1 by antifungal compounds from Lactobacillus brevis and Lactobacillus paracasei, isolated from dairy products. Probiotics Antimicrob. Proteins 2018, 10, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; Abdel-Galil, M.M.; El-Lithey, M. Melatonin counteracts oxidative stress in rats fed an ochratoxin A contaminated diet. J. Pineal Res. 2005, 38, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Wanigasuriya, K.P.; Peiris, H.; Ileperuma, N.; Peiris-John, R.J.; Wickremasinghe, R. Could ochratoxin A in food commodities be the cause of chronic kidney disease in Sri Lanka? Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 726–728. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Zakowska, Z. The elimination of ochratoxin A by lactic acid bacteria strains. Pol. J. Microbiol. 2005, 54, 279–286. [Google Scholar]
- Piotrowska, M. The adsorption of ochratoxin A by Lactobacillus species. Toxins 2014, 6, 2826–2839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowiak, P.; Śliżewska, K.; Nowak, A.; Chlebicz, A.; Żbikowski, A.; Pawłowski, K.; Szeleszczuk, P. Probiotic microorganisms detoxify ochratoxin A in both a chicken liver cell line and chickens. J. Sci. Food Agric. 2019, 99, 4309–4318. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, A. Fungi and mycotoxin problems in the apple industry. Curr. Opin. Food Sci. 2019, 29, 42–47. [Google Scholar] [CrossRef]
- Kadakal, Ç.; Nas, S.; Ekıncı, R. Ergosterol as a new quality parameter together with patulin in raw apple juice produced from decayed apples. Food Chem. 2005, 90, 95–100. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Serra, R.; Venâncio, A. Biodegradation of ochratoxin A by fungi isolated from grapes. J. Agric. Food Chem. 2002, 50, 7493–7496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.P. Immunotoxicity of mycotoxins. J. Dairy Sci. 1993, 76, 892–897. [Google Scholar] [CrossRef]
- Hatab, S.; Yue, T.; Mohamad, O. Removal of patulin from apple juice using inactivated lactic acid bacteria. J. Appl. Microbiol. 2012, 112, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Assaf, C.E.H.; De Clercq, N.; Van Poucke, C.; Vlaemynck, G.; Van Coillie, E.; Van Pamel, E. Effects of ascorbic acid on patulin in aqueous solution and in cloudy apple juice. Mycotoxin Res. 2019, 35, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Barukčić, I.; Bilandžić, N.; Markov, K.; Jakopović, K.L.; Božanić, R. Reduction in aflatoxin M1 concentration during production and storage of selected fermented milks. Int. J. dairy Technol. 2018, 71, 734–740. [Google Scholar] [CrossRef]
- Taheur, F.B.; Fedhila, K.; Chaieb, K.; Kouidhi, B.; Bakhrouf, A.; Abrunhosa, L. Adsorption of aflatoxin B1, zearalenone and ochratoxin A by microorganisms isolated from Kefir grains. Int. J. Food Microbiol. 2017, 251, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saladino, F.; Luz, C.; Manyes, L.; Fernández-Franzón, M.; Meca, G. In vitro antifungal activity of lactic acid bacteria against mycotoxigenic fungi and their application in loaf bread shelf life improvement. Food Control. 2016, 67, 273–277. [Google Scholar] [CrossRef]
- Ananthi, V.; Subramanian, R.S.; Arun, A. Detoxification of Aflatoxin B1 using Lactic acid bacteria. Int. J. Biosci. Technol. 2016, 9, 40. [Google Scholar]
- Khaneghah, A.M.; Chaves, R.D.; Akbarirad, H. Detoxification of aflatoxin M1 (AFM1) in dairy base beverages (acidophilus milk) by using different types of lactic acid bacteria-mini review. Curr. Nutr. Food Sci. 2017, 13, 78–81. [Google Scholar] [CrossRef]
- Juodeikiene, G.; Bartkiene, E.; Cernauskas, D.; Cizeikiene, D.; Zadeike, D.; Lele, V.; Bartkevics, V. Antifungal activity of lactic acid bacteria and their application for Fusarium mycotoxin reduction in malting wheat grains. LWT 2018, 89, 307–314. [Google Scholar] [CrossRef]


{kind=link}
| Mycotoxin | Fungi | Target Foods | Reference |
|---|---|---|---|
| Dioxynivalenol | Fusarium graminearum | Cereal products | [20] |
| Aflatoxins | Aspergillus flavus, A. nomius, A. parasiticus | Wheat, corn, rice, peanut, rice, pepper, cotton, nut tree and spices | [21] |
| Fumonisins | F. proliferatum, F. verticillioides, F. nygamai | Rice, figs, beer and corn | [22] |
| HT-2 Toxins | Fusarium spp. | Oat, corn | [23] |
| Ochratoxin | A. niger, A. ochraceus, A. carbonarius, Penicillium verrucosum, Neopetromyces spp., Petromyces spp. | Fruits, coffee, spices, wine, dried cocoa, beans, corn, cereal, grains, and rice. | [24] |
| Patulin | P. patulum, P. expansum, P. urticae, A. terreus, A. clavatus, Byssochlamys nivea, P. patulin | Apricots, grapes, grape fruit, peaches, pears, apples, fruit juice, meat, cheese and cereals | [25] |
| T-2 toxin | F. sporotrichioides, F. poae, F. equiseti, F. acuminatum | Wheat, corn, oats, barley, rice, beans, and soya beans | [26] |
| Zearalenone | F. graminearum, F. culmorum, F. cerealis, F. equiseti, F. crookwellense, F. semitectum | Cereal products, wheat, barley, oat, soybean and corn | [23] |
| Microorganism | Target Mycotoxin | Degradation% | Reference |
|---|---|---|---|
| Lactobacillus rhamnosus GG, L. rhamnosus LC-705 | Aflatoxin B1 | 80% | [49] |
| L. amylovorus, L. rhamnosus | Aflatoxin B1 | 50% | [50] |
| L. casei LOCK 0920, L. brevis LOCK 0944, L. plantarum LOCK 0945 | Aflatoxines (Bl, B2, G1, G2) | ~50% | [51] |
| L. plantarum, Lactococcus lactis | Aflatoxin B1 | 81% | [52] |
| Streptococcus thermophiles, L. bulgaricus, L. plantrium | Aflatoxin M1 | 11–34% | [53] |
| L. casei | Aflatoxin B1 | 49.2% | [54] |
| L. paracasei LOCK 0920, L. brevis LOCK 0944, L. plantarum LOCK 0945 | Aflatoxin B1 | 39–55% | [55] |
| L. plantarum C88 | Aflatoxin B1 | 60% | [56] |
| Lactic acid bacteria strains | Aflatoxins B1 and B2 | ND | [57] |
| L. casei LOCK 0920, L. brevis LOCK 0944, L. plantarum LOCK 0945 | Ochratoxin A | ~50% | [51] |
| L. acidophilus VM 20, Bifidobacterium animalis VM12 | Ochratoxin A Patulin | 95% 80% | [58] |
| Pediococcus parvulus | Ochratoxin A | 90% | [59] |
| L. rhamnosus CECT 278T | Ochratoxin A | 97% | [60] |
| L. brevis 20023 | Patulin | ND | [61] |
| L. plantarum GT III | Deoxynivalenol | 56–66% | [62] |
| Lactic acid bacteria strains | Deoxynivalenol, fumonisins B1, fumonisins B2 | 55%, 82%, and 100% | [63] |
| Lactic acid bacteria | Fumonisin B1 Zearalenone | 56–67% 68–75% | [64] |
| L. rhamnosus GG (ATCC 53103), L. plantarum A1 | Zearalenone | ND | [65] |
| L. paracasei, L. lactis | Zearalenone | 55% | [66] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muhialdin, B.J.; Saari, N.; Meor Hussin, A.S. Review on the Biological Detoxification of Mycotoxins Using Lactic Acid Bacteria to Enhance the Sustainability of Foods Supply. Molecules 2020, 25, 2655. https://doi.org/10.3390/molecules25112655
Muhialdin BJ, Saari N, Meor Hussin AS. Review on the Biological Detoxification of Mycotoxins Using Lactic Acid Bacteria to Enhance the Sustainability of Foods Supply. Molecules. 2020; 25(11):2655. https://doi.org/10.3390/molecules25112655
Chicago/Turabian StyleMuhialdin, Belal J., Nazamid Saari, and Anis Shobirin Meor Hussin. 2020. "Review on the Biological Detoxification of Mycotoxins Using Lactic Acid Bacteria to Enhance the Sustainability of Foods Supply" Molecules 25, no. 11: 2655. https://doi.org/10.3390/molecules25112655
APA StyleMuhialdin, B. J., Saari, N., & Meor Hussin, A. S. (2020). Review on the Biological Detoxification of Mycotoxins Using Lactic Acid Bacteria to Enhance the Sustainability of Foods Supply. Molecules, 25(11), 2655. https://doi.org/10.3390/molecules25112655