1. Introduction
Hyaluronan (HA) is one of the important physiological bio macromolecules which is ubiquitous in the body. It is a high molecular weight anionic and linear polysaccharide consisting of repeating disaccharide units of β-1,3-
d-glucuronic acid and β-1,4-
N-acetyl-
d-glucosamine. The pKa of the monomer
d-glucuronic acid and the polymer has been estimated to be 3.23 and 2.9, respectively, and the distance between adjacent charges along the polymer chain is about 1 nm [
1]. Hyaluronan is regarded as a wormlike polyelectrolyte with an intrinsic persistence length ranging from 4.5 to 9 nm in high ionic strength solutions [
2]. The role of hyaluronan is quite diverse, ranging from being important for cell proliferation, cell migration, the connective tissue and the synovial fluid. The latter one is a multi-component aqueous system that contains, in addition to hyaluronan, a complex mixture of phospholipids, proteins, proteoglycans and ions [
3,
4]. The reason for the interest in the working mechanisms of the synovial joints is due to their excellent lubrication properties with friction coefficients as low as 0.002–0.006 under high shear and load conditions [
3,
5]. The secret of this performance is thought to be a fluid water layer in between the sliding surfaces and friction reduction occurs via hydration repulsion generated by water interacting with biomolecules adsorbed from the synovial fluid to the cartilage surface, e.g., lubricin, phospholipids and HA [
4,
6,
7,
8,
9]. Different investigations suggest that not only one component but rather a mixture of specific constitutes has to be present to allow for the exceptional good lubrication properties in joints as, for example, the combined presence of albumin, γ-globulin, DPPC and HA leads to reduction by a factor of 4 compared solutions with the single components [
10,
11]. One suggestion is that the sliding occurs via boundary lubrication in between multilayer phospholipid structures, which form at the cartilage surface [
3,
12]. The important sliding plane seems to be in between the phospholipid head group structures where a thin layer of water is maintained [
12,
13,
14]. The question arises how the association structures adapt to stable well-defined structures that promote a bond but yet retain a fluid and easily sheared layer of water under high loads and shear conditions.
Among the different components in the synovial fluid, hyaluronan and phospholipids are thought to be of major importance for lubrication. This view arises from the fact that phospholipids are very efficient in reducing boundary friction [
15,
16,
17]. On the other hand, hyaluronan has been shown to be able to relieve osteoarthritis and degenerated articular cartilage by injection into the knee [
18]. The effect is mainly due to its ability to control the viscosity of the synovial fluid and retain water [
5]. Hydrophobic regions representing repeating structures of eight CH groups along the HA chains have been identified [
19], and these hydrophobic sites are perceived as being important for network formation and aggregation of HA [
20]. HA is able to reduce friction on its own to some extent, and it is important for the rheological properties of the synovial fluid as it controls viscosity [
10,
21,
22].
For these reasons the interaction of 1,2-dipalmitoyl-sn-glycero-3-phosphocholine (DPPC) and HA has been the scope of different theoretical and experimental investigation to reveal the associate structures formed, their interactions and their behavior under load. Experimental investigations have reported on the accumulation of HA in-between dipalmitoylphosphaditcholin bilayers [
23] and adsorption of HA to DPPC bilayers [
17,
24,
25,
26,
27]. On the other hand, the study performed by Herzog et al. using HA with a high molecular weight (MW) could not observe any adsorption of HA to DPPC [
28]. This observation is not in objection to the former cited studies, as an investigation reporting on the effect of the MW on the interaction shows that HA with high MW has a weak interaction with DPPC [
29]. The reason for this might be the intrinsic structure of HA, where low MW species may form more intermolecular interactions as opposed to high MW species where more intramolecular interactions are expected.
Both Na
+ and Cl
− ions have been reported to bind to DPPC membrane bilayers. Since the binding of Cl
− is weaker than that of Na
+, the DPPC bilayer in NaCl electrolyte is expected to have a net positive surface charge [
30]. Investigations of ion-membrane interactions suggest that calcium ions can be found at low concentrations of 1.2 mM to 2.4 mM in the synovial fluid [
31,
32], and it is well-known that CaCl
2 is able to alter the structure of DPPC bilayers [
33,
34,
35,
36,
37,
38,
39] and the charge density of HA [
40]. Na
+ and Ca
2+ ions bind to phospholipid head groups via coordination of the cations by three and four lipid carbonyl oxygens, respectively [
41]. The stoichiometry between calcium ions and phospholipids has been considered in several studies, with somewhat conflicting results. For instance, for 1-Palmitoyl-2-oleoylphosphatidylchol, the lipid/Ca
2+ ratio was reported to be 1:2 in 5 M CaCl
2 [
39]. A binding stoichiometry of Ca
2+/DPPC of 1:1 has also been proposed, and the binding constants of Ca
2+ and Na
+ were reported to be 37 M
−1 and 0.25 M
−1, respectively, signifying significantly stronger binding of Ca
2+ ions. Still, other binding stoichiometry have been reported by Disalvo et al., where the phospholipid/Ca
2+ ratio of 2:1 was obtained for the inner monolayer and 1:1 for the outer monolayer [
42]. The interaction between metal ions and phospholipids is also temperature dependent, and the data suggest stronger binding in the gel phase than in the fluid phase. Unlike many anionic biopolymers, hyaluronan does not precipitate at high Ca
2+ concentrations, which is related to its strong water binding capacity. Small angle X-ray scattering (SAXS) measurements in the semi dilute regime also suggest minor effects of calcium ions [
40]. However, anomalous SAXS measurements have shown that in mixed solutions containing Na
+ and Ca
2+ ions, the calcium ions are preferentially located next to the hyaluronan chain [
40].
It has also been shown that low concentration of calcium ions increases the interaction between HA and DPPC [
29,
43]. It was speculated that a possible mechanism is a charge reversal of the phosphate group by calcium ions resulting in a positive charge of DPPC. Indeed, theoretical calculations confirm that calcium ions promote long-lasting interactions between the carboxylate group of HA and the phosphate group of the phospholipid by creating a bridging connection [
44]. Further, a reinforcement of the HA network by the adsorption of phospholipids has been observed [
45,
46].
To gain understanding of the interactions of the synovial fluid under loading conditions is challenging as only a limited number of techniques allow exploration of this situation on the molecular level. Investigations on the phase behavior of solid supported HA–DPPC composite bilayers demonstrated that the phase behavior is changed in comparison to that of DPPC bilayers alone. The reports indicate that HA–DPPC bilayers exhibit an increased robustness against high hydrodynamic pressure as compared to DPPC bilayers alone, allowing for a reversible phase change [
27,
47]. Further, a decoupling of the two DPPC leaflets could be observed for low MW of HA in the presence of calcium ions.
This investigation aims to further elucidate the effects of calcium ions and molecular weight on the molecular interactions between HA and DPPC by a combination of simulations and experimental approaches. To this end, we utilized DPPC vesicles as here no solid–liquid interfaces are present, which simplifies the system. The samples were studied under near physiological conditions of 150 mM sodium chloride and with 10 mM calcium chloride added. In former studies [
29] it was observed that the MW of HA had an impact on the phase behavior and bilayer structures of DPPC, therefore, the phase behavior of HA was investigated by differential scanning calorimetry (DSC) and dynamic light scattering (DLS) in the MW range from 10 kDa to 2500 kDa. Based on these results, we decided to use HA of 10 kDa and 1500 kDa for small angle neutron scattering (SANS) measurements as this study focused on the formed structures and the structural arrangement of DPPC and HA to elucidate if the observed changes in the DSC could be attributed to a changed phospholipid packing. The HA with an MW of 10 kDa was chosen as we expected a stronger effect due to the small size if it would penetrate the lipid bilayer than that achieved by HA with an MW of 1500 kDa. To further investigate the impact of Ca
2+ ions on the HA–DPPC arrangement, SAXS measurements on vesicles were performed. The decision on the 250 kDa for these measurements was based on the other measurements as the impact for low MW of HA is stronger. However, the usage of a too short HA polymer chain would have made it difficult to see an adsorbed layer like that experienced in former studies [
29]. The electrophoretic mobility measurements were performed to achieve an understanding of the Ca
2+ binding to the vesicles and how this influences the association with HA, and in these measurements HA with sufficiently large molecular weight are needed, and our choice fell on 800 kDa. To obtain further insight on the molecular level all-atom molecular dynamics (MD), simulations were performed and used as an aide for interpreting the experimental findings. With the available computational resources, modeling long chains of HA, with atomic mass over dozens kDa at the atomic level, is very difficult, if not impossible. To model longer HA chains, coarse-grained MD simulations could be utilized, although they could be insufficient to obtain the necessary level of details [
48].
3. Discussion
The DSC measurements show two characteristic peaks, one for the pre-transition and one for the main transition which is in agreement with the literature [
49,
50]. However, the data indicate that an interaction of HA with the glycerol backbone at the hydrophobic–hydrophilic interface, as a shoulder, is present at the main transition [
51]. This is a realistic scenario for HA as it has been reported that HA has hydrophobic patches, which can bind to phospholipids [
20,
52]. However, in the simulation results reported here, hydrophobic interactions between HA and DPPC are identified exclusively with the CH
3-groups present in the head group. An interaction of HA with the alkyl chains of the lipids would induce a change of the main transition temperature and, in parallel, an increase in the width of the main transition [
51]. As we only observe a slight shift of the main transition along with a change in the transition enthalpy it can be ruled out that HA interacts with the tail group of DPPC.
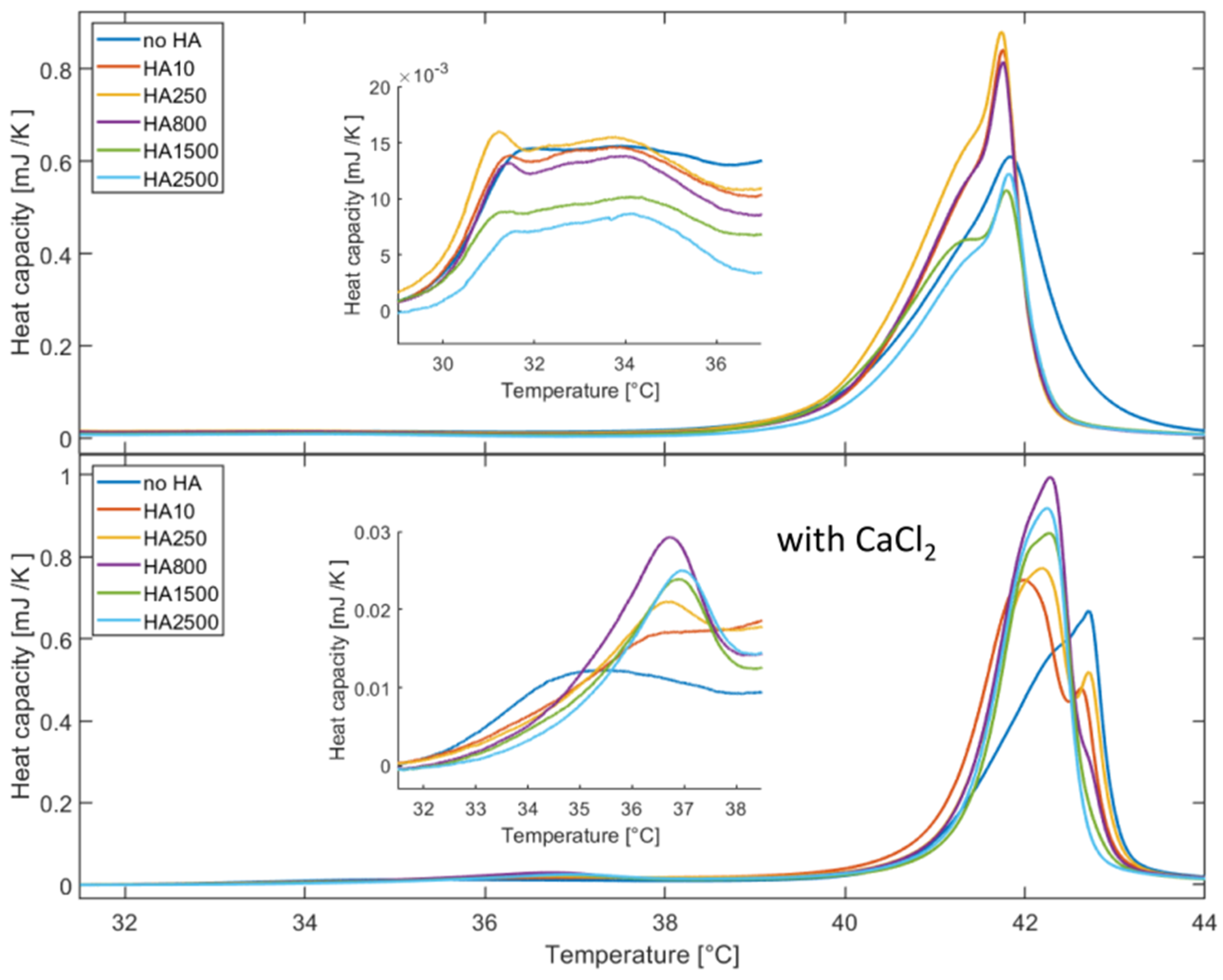
Further, the DSC data also show that calcium ions significantly change the interactions in the DPPC and DPPC/HA systems. Our data demonstrate that Ca
2+ binds to the DPPC bilayer and thereby affects the phase behavior. The addition of HA leads to a strong decrease in the pre- and main-transition, which is significantly stronger for HA with an MW lower than 800 kDa. Such changes are normally regarded as a sign for impurities disturbing the alkyl chain packing [
50]. However, as also the shapes of the transition peaks are changed, while in parallel a second peak at the main transition occurs; thus, it can be deduced that HA only interacts with the outer headgroup [
51] as detailed in the simulation work reported here. We speculate that this interaction changes the mobility and degree of freedom of the DPPC molecules in the outer layer, making the system more stiff and inflexible. The appearance of the second peak can be understood in terms of two phases being present: one in the inner DPPC layer of the vesicles not interacting with HA and one associated with the outer layer affected by HA adsorption. This leads to a different phase behavior and a decoupling of the bilayer leaflets as observed in other studies [
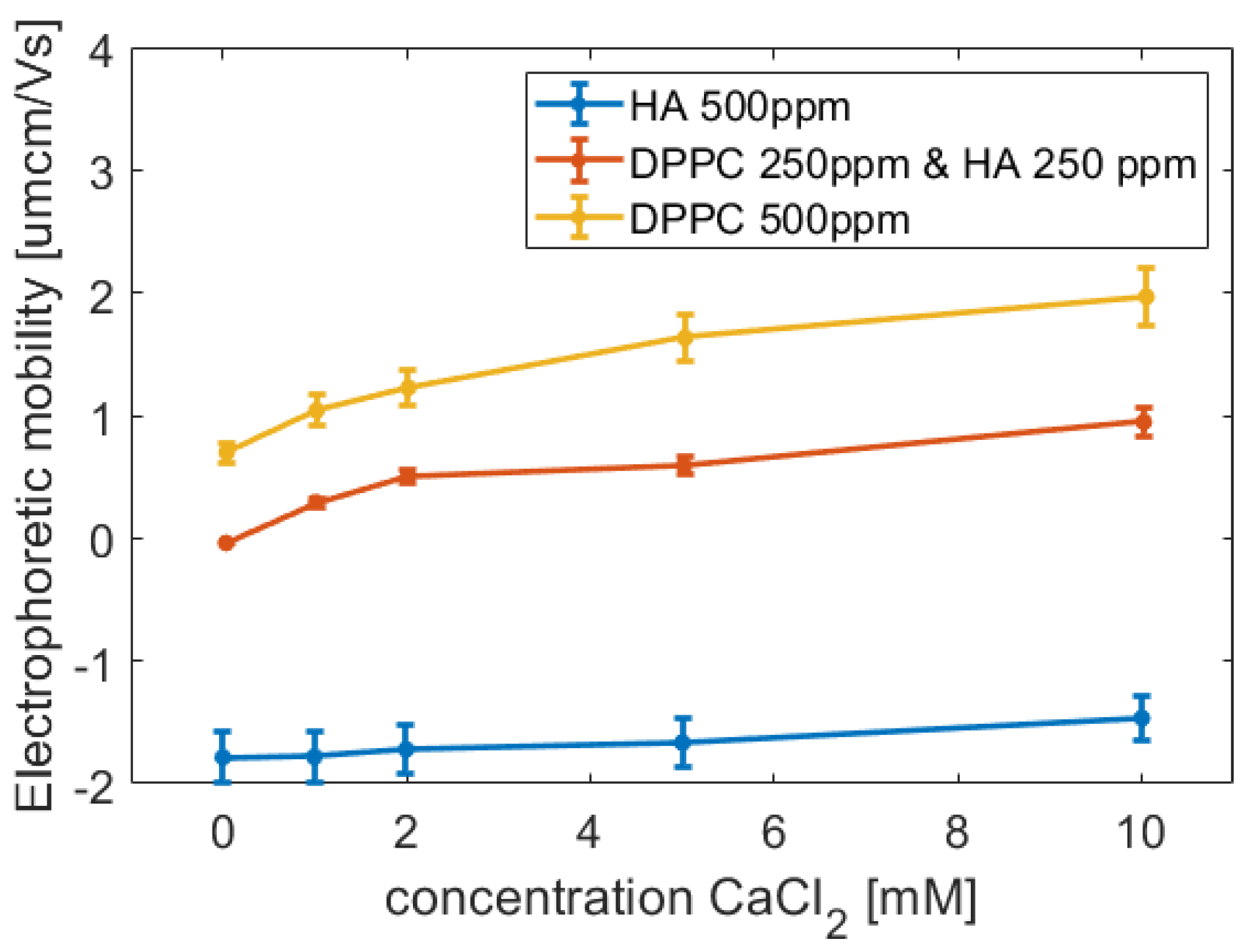
47]. On the basis of the electrophoretic mobility measurements, we postulate that the binding to the outer layer is favoured by electrostatic interactions, but MD simulations also demonstrate hydrogen bonding and hydrophobic interactions between HA and DPPC bilayers. In solutions containing both HA and DPPC, the charge of the aggregates is less than that of the vesicles in absence of HA, indicating partial charge compensation only possibly by the binding of HA to DPPC.
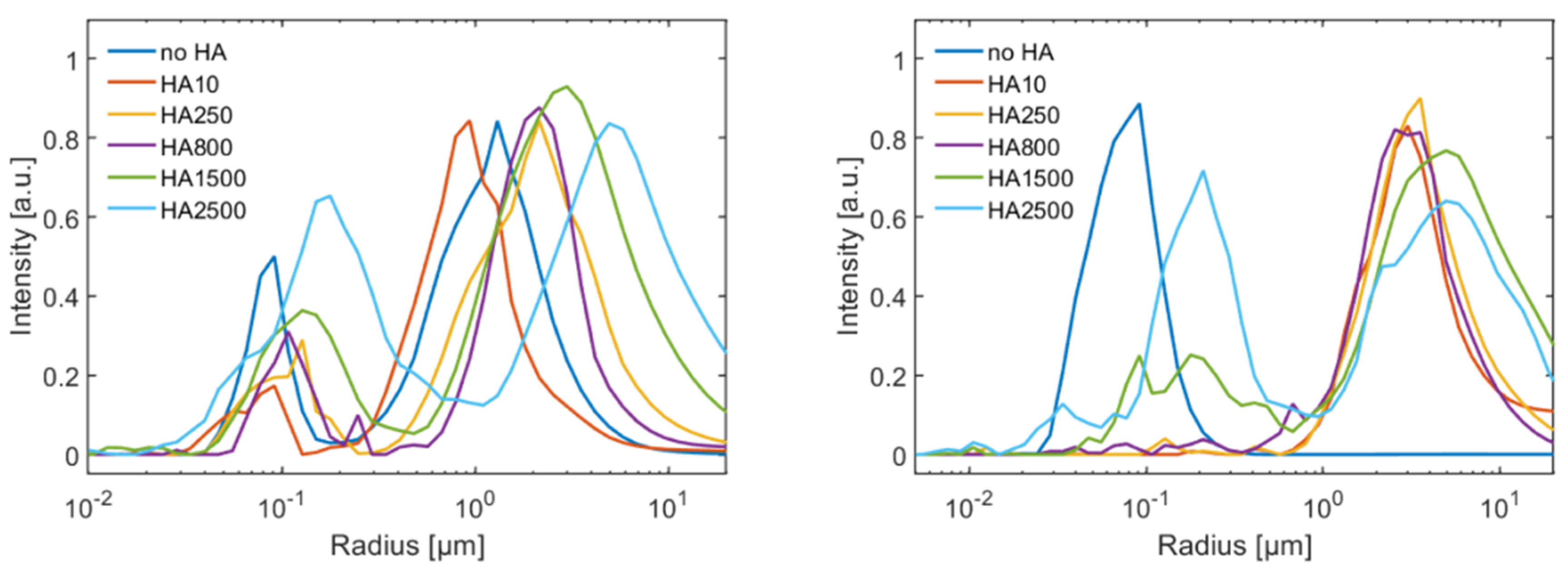
The DLS measurements show that addition of HA, both in presence and absence of calcium ions, leads to the increase in the hydrodynamic radius of the vesicles. Further vesicle aggregation occurs, and the extent of aggregation depends on the MW of HA. As discussed above, the DSC also indicates a change in the interaction depending on the MW. The transition enthalpy decreases for HA with an MW higher than 800 kDa, which indicates a modified interaction mechanisms. This can be understood by the change of the polymer chain conformation as high and low MW of HA have the same structure and number of charged carboxylate groups per monomer. At low MW, HA can be regarded as a short chain having no or few intramolecular interactions. This changes as the chain gets longer and becomes progressively more able to coil and develop intramolecular interactions that reduce the ability of HA to interact with the DPPC bilayer. Further, different hydrodynamic radii of the vesicles as a function of the MW of HA can be understood by the different conformations of the HA polymer chain. HA with an increased MW will lead to a larger increase in the hydrodynamic radius due to the larger extension of the polymer chain into the solution. Our data also suggest that a critical MW for this change in the interaction is around 800 kDa, but it is as yet not clear why this transition occurs at such a high molecular weight.
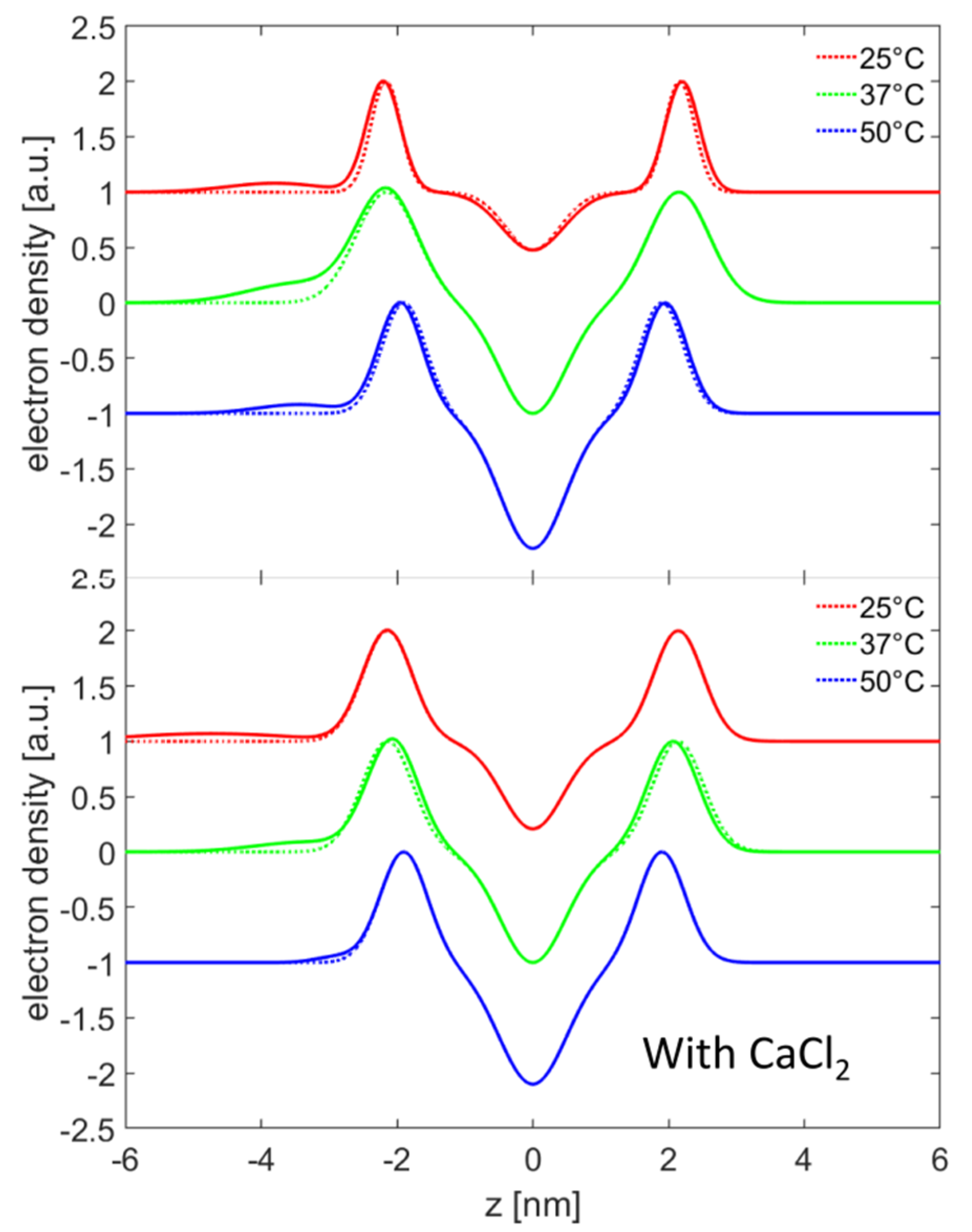
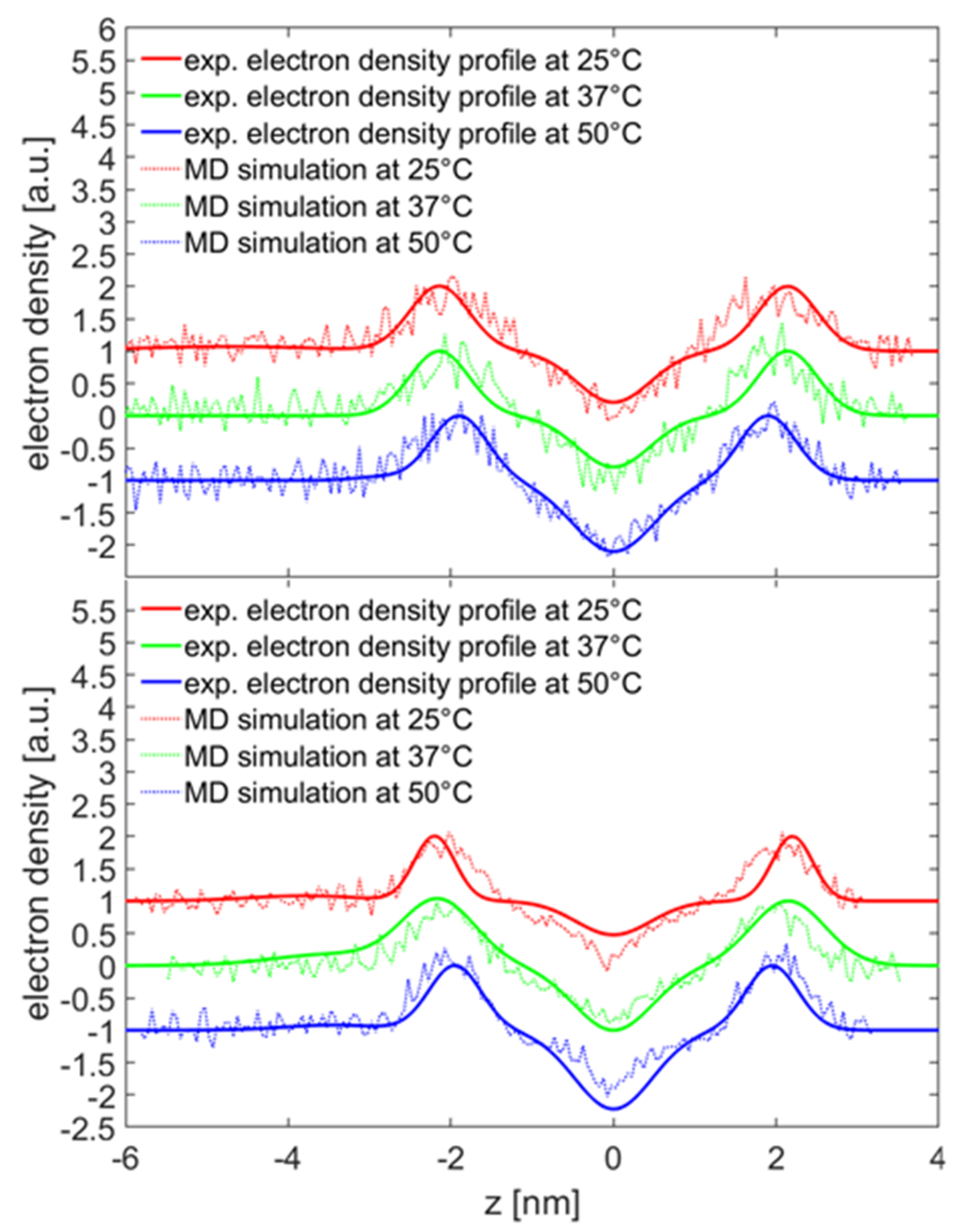
The SAXS measurements on DPPC vesicles with HA were performed at three bilayer phases: gel phase (L
β0), rippled phase (P
β0) and fluid phase (L
α) [
53]. The different phases have different structural conformations, and the most clearly observed change was a reduction in the d-spacing and head-to-head distance as the bilayer makes a transition to the L
α phase. The obtained values for the head-to-head distance in the different phases agree well with the literature data [
54]. However, the results for the d-spacing do not match. A possible reason might be the preparation method as in this study vesicles with a diameter of 100 nm were used. Such vesicles could have high stresses due to the high curvature which is expected to have an impact on the d-spacing [
55]. The fits show that the electron density ρ
T of the phospholipids increase as the L
α phase is reached which can be due to a changed packing of the DPPC molecules in the vesicles due to a higher disorder in the bilayer as a result of the melting of the alkyl chains. Such an increased volume per molecule is also reported in the literature [
56]. Upon the addition of HA, no detectable change of the vesicle bilayer structure can be observed as the scattering curves are similar to each other and, thus, the obtained electron density profiles are indistinguishable. A diffuse layer of HA can, however, be seen, which is most compact at 37 °C where the DPPC vesicles are in the rippled phase. However, in the presence of HA an increased d-spacing can be recognized for the P
β0 and the L
α phases. This hints at an accumulation of HA in between the bilayer as reported by Kreuzer at al. [
23]. It remains an open question how HA enters the inter bilayer region, as vesicles consist of closed lipid shells and an HA-induced disruption of the bilayer seems unlikely. A scenario where HA penetrates thought the membranes can be ruled out by the DSC measurements. Some parts of the multillamellar structure could arise from aggregates of vesicles where it is possible for the HA to accumulate between bilayers.
In the presence of 10 mM, only CaCl
2 unilamellar vesicles can be found as also reported in the literature [
39]. In contrast to the cited work, oligo lamellar structures could be detected at 50 °C, which could be due to the high concentration of NaCl used in our study [
47]. In addition to promoting the formation of unilamellar vesicles, the addition of Ca
2+ also has a strong effect on the electron density in the tail group region. The higher electron density level indicates a higher packing of the lipids induced by Ca
2+, as already suggested by Kataoka et al. 1985 [
34] and Aruga et al. 1985 [
33]. The extracted head-to-head distance in the three different phases for bilayers in contact with 150 mM NaCl solutions, in absence and presence of 10 mM CaCl
2, showed no effect due to calcium ions. This is only partially consistent with the reports in the literature [
39]. Here, an induced increase in the bilayer thickness in the L
α phase of about 0.2 nm was observed in the presence of 10 mM CaCl
2. The origin was speculated to be a changed packing induced by the electrostatic potential of the adsorbed Ca
2+ ions. However, other studies do not report any change due to the presence of Ca
2+ [
36].
When both calcium ions and HA are present, a layer of HA could be detected which had a more compact structure than in sodium chloride solutions, indicating a stronger interaction between the DPPC bilayer and HA chains, which is also observed in the MD simulations. The population of pure unilamellar vesicles changed and bilayer structures could be detected. The question arises if the presence of the double bilayer vesicles is a consequence of disruption and reorganization of vesicles due to HA or if HA induced an aggregation with a favorable distance between bilayers. The aggregation could also be seen by DLS. However, for a disruption of the vesicles, a strong interaction of HA and the alkyl chains has to be present, which can be ruled out by the DSC and small angle scattering measurements.
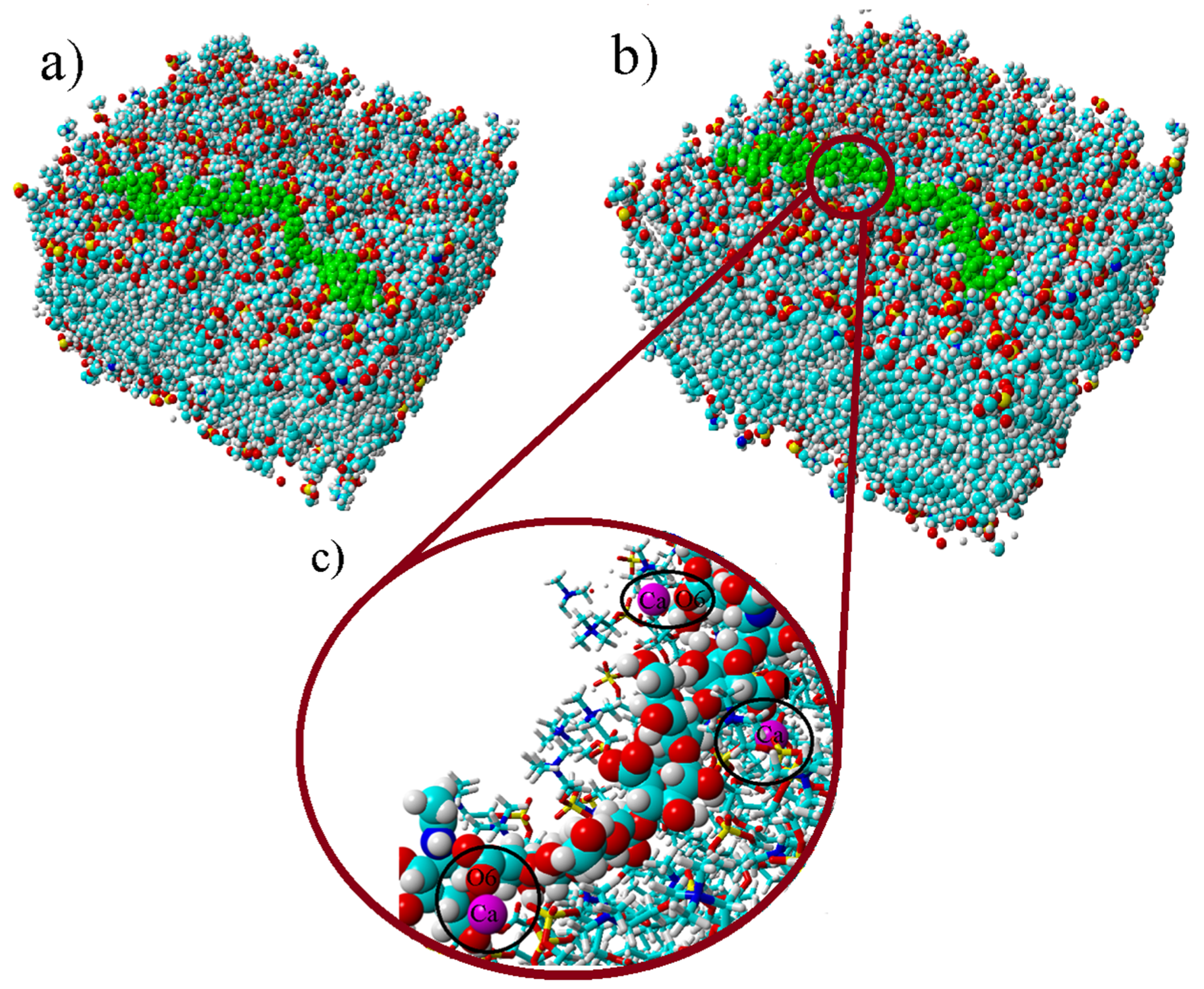
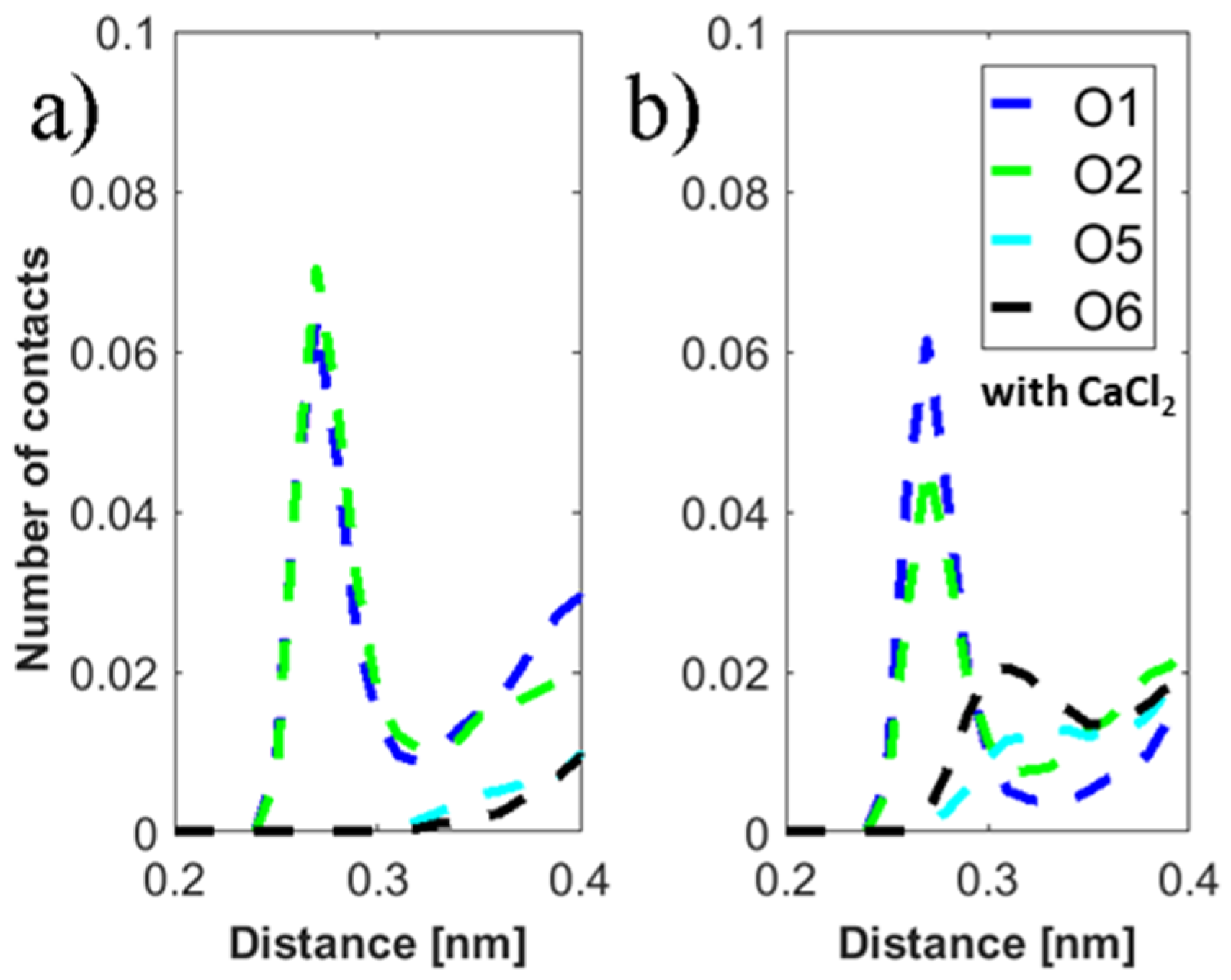
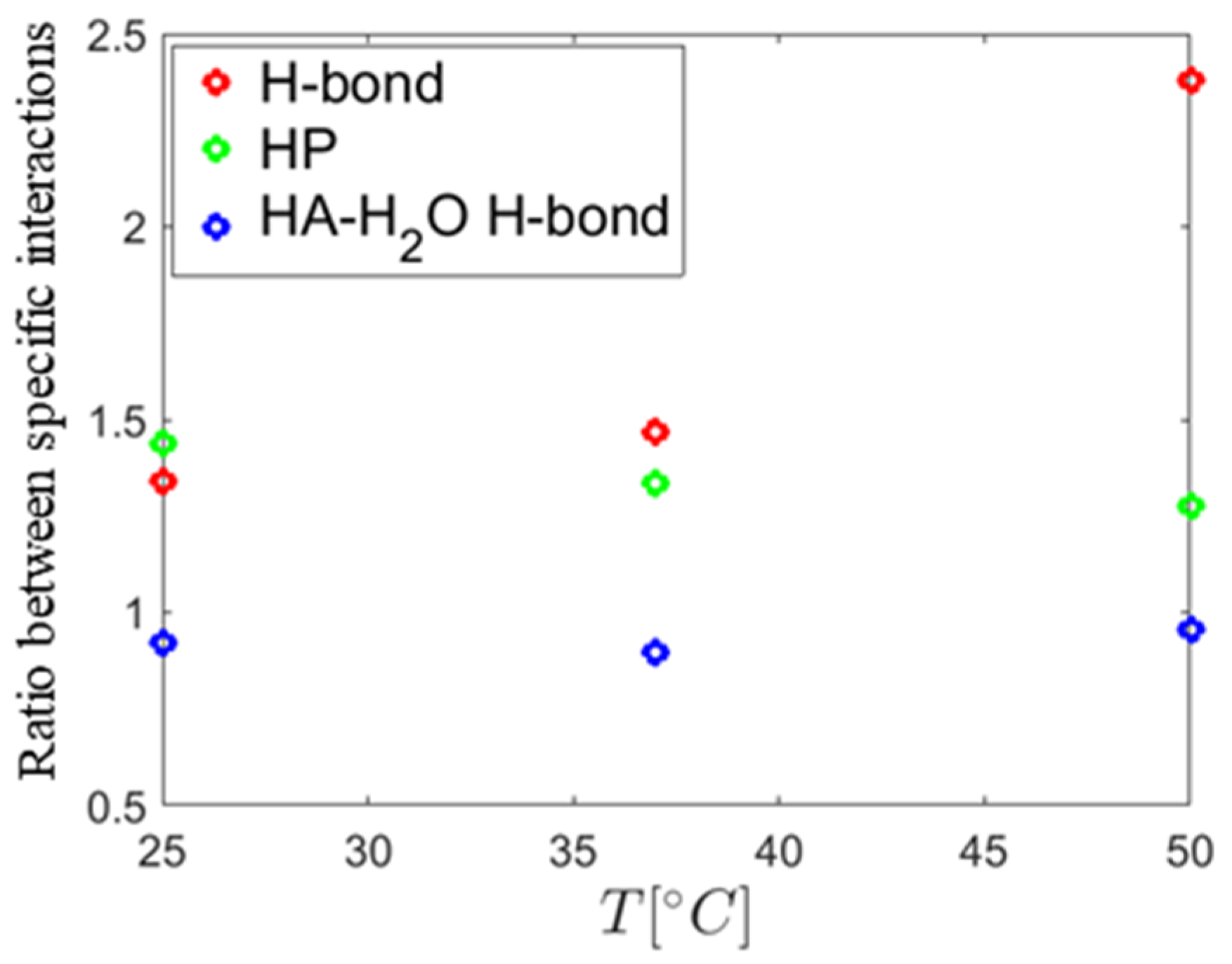
The MD simulations yield insight to the direct binding of DPPC and HA. As one can see in
Figure 8, the conformation of HA was not significantly changed during the simulation at 37 °C in calcium chloride solution. However, for other cases, HA was more detached from the surface. In general, no strong effect on the binding distribution behavior could be observed for the simulated conditions, which can be a result of the short chain used. However, for conditions where calcium is present, a total increase in the number of interaction points, either by hydrophobic contacts or HA–DPPC hydrogen bonds, is observed, which can be interpreted as adsorption of HA closer to the bilayer, which is consistent with the enhanced adsorption observed by the experiments. At 50 °C, the relative number of H-bonds formed in calcium ion containing solution is higher than in sodium ion containing solutions, indicating a stronger impact of calcium ions on the binding behavior at higher temperatures. In the presence of calcium ions, the number of hydrogen bonds to water for HA is decreased. This can be explained by the binding of calcium ions to HA and the closer binding of HA to the DPPC bilayer in the presence of calcium ions, which reduce the possibility for water binding [
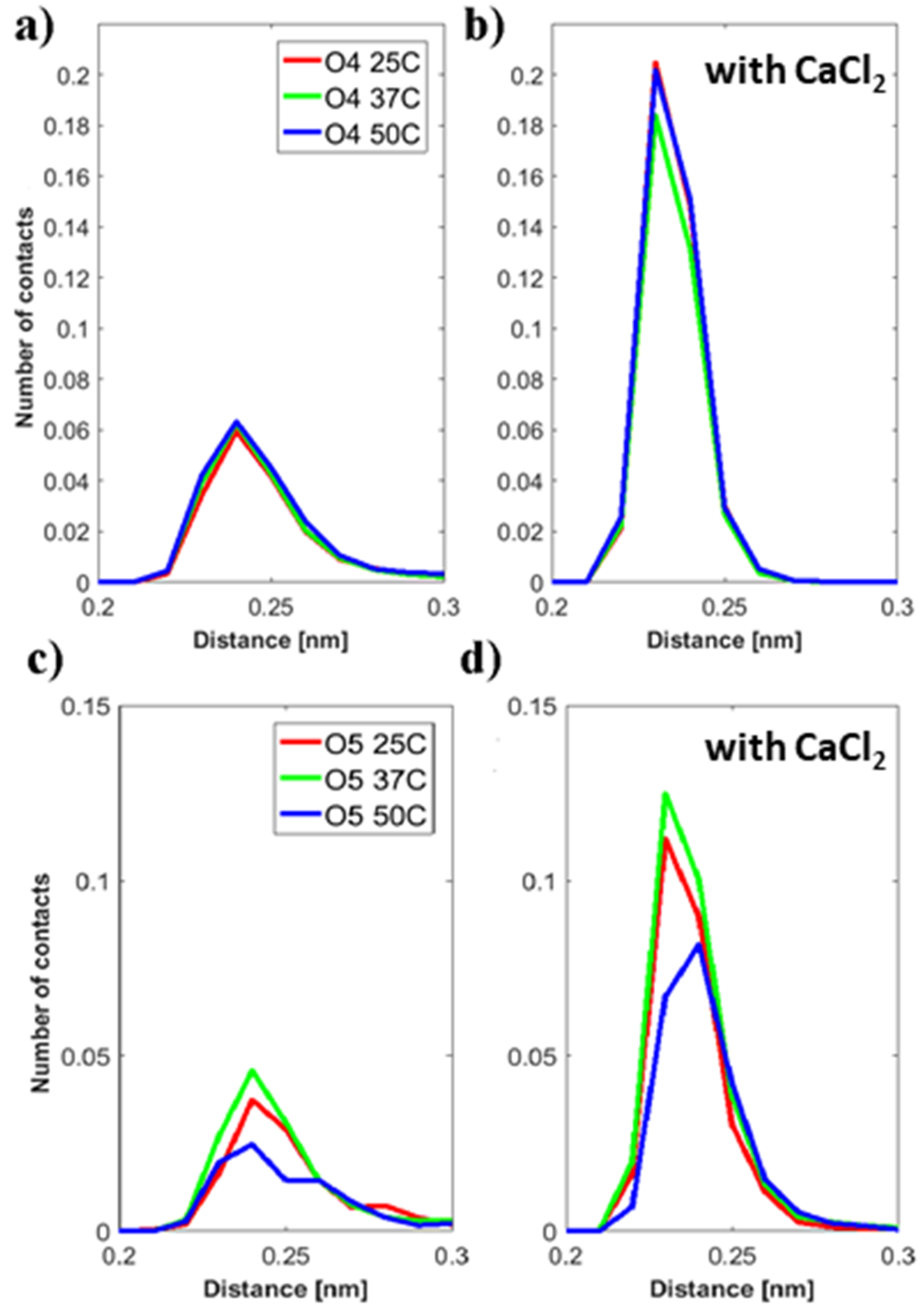
44]. The distribution of distances of ions to the HA-O5 (carboxylate) and DPPC-O4 (phosphate) atoms shows more calcium ions than sodium ions in close proximity to these groups. Further, calcium-ion-mediated ionic bridges between these groups were observed. Such bridges may stabilize the DPPC–HA binding and thereby lead to an increased number of H-bonds and hydrophobic contacts. This property might be of higher importance at elevated temperatures as indicated by the increase in the relative number of hydrogen bonds between DPPC and HA at 50 °C and might explain the change in the DSC data. For HA, a similar behavior is seen as the calcium ions are also moved closer to the carboxyl group due to electrostatic attraction. This can also explain a lower binding of water molecules to the HA-O5 atom as calcium ions are more directly involved in electrostatic interaction and, thus, result in a shift of water away from carboxyl group.
4. Materials and Methods
4.1. Materials
DPPC was purchased from Avanti polar lipids (catalogue No. 850355P) (Alabaster, AL, USA), and HA was bought from Creative PEGworks (Durham, NC, USA). HA samples with weight averaged molecular weights of 10 kDa (Catalogue No. HA-101), 250 kDa (catalogue No. HA-103), 800 kDa (catalogue No. HA-104), 1500 kDa (catalogue No. HA-106) and 2500 kDa (catalogue No. HA-107) were used. All compounds were used as received. In the following, HA with an MW of 10 kDa will be referred to as HA10, HA with an MW of 1500 kDa as HA1500 and so on. Sodium chloride (assay ≥99.8, catalogue No. 31434, purchased from Sigma-Aldrich, Munich, Germany), calcium chloride dehydrate (assay ≥99, catalogue No. 5239, purchased from Carl Roth, Roth, Germany) and ultrapure water (Milli-Q, resistivity 18.2 MΩ cm) were used to prepare all aqueous solutions. The electrophoretic mobility measurements were performed with sodium hyaluronate (HA) with a molecular weight of about 0.9 × 106 Da, which was received as a gift from Novozymes (Nottingham, UK).
4.2. Sample Preparation
The desired amount of lipid powder was dissolved in chloroform and transferred into a glass vial. The chloroform was then evaporated under a flow of nitrogen, whereby a lipid film formed on the walls of the glass vial. Residual chloroform was removed by keeping the vials in an oven over night at 60 °C. Next, the aqueous solution was added, either 150 mM NaCl or 150 mM NaCl with 10 mM CaCl2, to achieve a target concentration of the phospholipids of 10 mg/mL. The solution was heated to 55 °C and vortexed for 5 min, which resulted in a turbid solution of multillamellar vesicles with broad size distribution. To achieve further dispersion of the phospholipid aggregates, the samples were kept at 55 °C in a thermomixer (BioShake Series, Analytik Jena, Jena, Germany) using a rotation rate of 350 rpm for 3 h, interrupted by vortexing each hour. In the final step, the multillamellar vesicles were extruded 35 times through a membrane with 0.2 µm pore size (Nuclepore Track-Etched Polycarbonate Hydrophilic Membranes, GE Healthcare Life Science, Little Chalfont, UK) making use of the Avanti Mini-Extruder (Avanti Polar Lipids Inc., Alabaster, AL, USA) in order to form almost unilamellar vesicles. Extrusion was carried out at 55 °C and the solution was kept at 55 °C until use to prevent any aggregation. The DSC data that are consistent with the literature data suggest that hydrolysis of DPPC at the high temperature is negligible.
For DPPC-vesicle/HA samples, a HA solution was prepared by adding HA to the aqueous salt solution, and the solution was stirred until HA was completely dissolved. The mass concentration of HA in the solution was set to be equal to the mass concentration of DPPC, both being 8 mg/mL. Finally, the HA solution was mixed with the vesicles solution in a ratio of 1:1 at 55 °C and kept at this temperature until use.
4.3. Small Angle X-ray Scattering and Analysis
The small angle X-ray scattering experiments were performed at the P12 beamline, Petra III, DESY, Hamburg, Germany, using an X-ray energy of 10 keV [
57]. The samples and the aqueous salt solution for background correction were measured in glass capillaries with a diameter of 1 mm and wall thickness of 0.01 mm. A Linkam (Tadworth, UK) heating stage was used for temperature control. Measurements were performed at: 25 °C, 37 °C and 50 °C. DPPC and DPPC/HA samples were studied in solutions conditions of 150 mM in absence and presence of 10 mM CaCl
2. The concentration of DPPC and HA was 4 mg/mL, respectively, and HAs with different molecular weights were used.
To fit the SAXS data, a model electron density profile was built and a scattering intensity curve was calculated. This curve was fitted to the measured datasets of the samples. The scattering curves of the DPPC-vesicles usually show two characteristic kinks at q ≈ 1 nm
−1 and q ≈ 1.81 nm
−1, which are clear signs for the existence of “multi” lamellar structures. Therefore, a combination of form factors was used to account for the simultaneous presence of “multi” and single bilayer structures. Different electron density profiles were used for the form factors. We used a model developed by Pabst et al. [
58]. Here, a single bilayer structure is described by a combination of three Gaussian curves with two curves for the heads and one curve for the tails:
The electron density profile is symmetric with its centre set to z = 0. The distance between the head group and the centre is denoted by z
H. The parameters σ
H and σ
T describe the width of the Gaussian curves for the head group and tail group, respectively. Δρ
H and Δρ
T are defined as the difference between the electron density of the head groups and the tail groups to the electron density of water Δρ
H/T = ρ
H/T − ρ
Water. For a further reduction in the parameter space, ρ
s(z) is normalized by Δρ
H, which leaves only one parameter Δρ
r describing the ratio of the electron densities of the head groups and tails:
Thus, ρ
s(z) can now be written as [
58]:
In order to account for a double bilayer or multilayer structure,
ρd, the structure of the single bilayer, was doubled, and an additional parameter
d was introduced, which describes the distance between the centres of the bilayers (repeat distance):
To calculate the scattered intensity of mixtures of lamellar structures, the different form factors have to be summed and the following formula is used:
A is a scaling factor, with
bkg, a constant, taking into account the incoherent background. The factor
n scales the amount single bilayer and multilayer structures with a number of
x layers. This term can be expanded to also include the desired number of multilayers of the system to be analyzed. This model has the advantage that the overall scattering intensity is calculated by using individual form factors for each structure, and no structure factor for multillamellar structures is needed. For mixtures with a maximum number of
x layers, the ratio of unilamellar to multilamellar vesicles
R can be calculated:
The use of a combination of a second form factor for the double bilayer instead of introducing a structure factor has the advantage that it is possible to add an extra layer to account for the adsorption of HA to the outer vesicle shell. Here, adsorption can only occur to one side of the bilayer structure as HA was added after the vesicles were formed. The adsorbed layer (HA) was also described by a Gaussian function and added to the ρ
s(z) and ρ
d(z) profiles from Equations (3) and (4) to yield:
With ρ
rHA being defined as
The fitting was performed with MATLAB (MATLAB version 8.2, MathWorks, Natick, MA, USA) making use of the non-linear curve-fitting routine “lsqcurvefit” (part of the MATLAB Optimization Toolbox), which uses a least-square method.
4.4. Small Angle Neutron Scattering
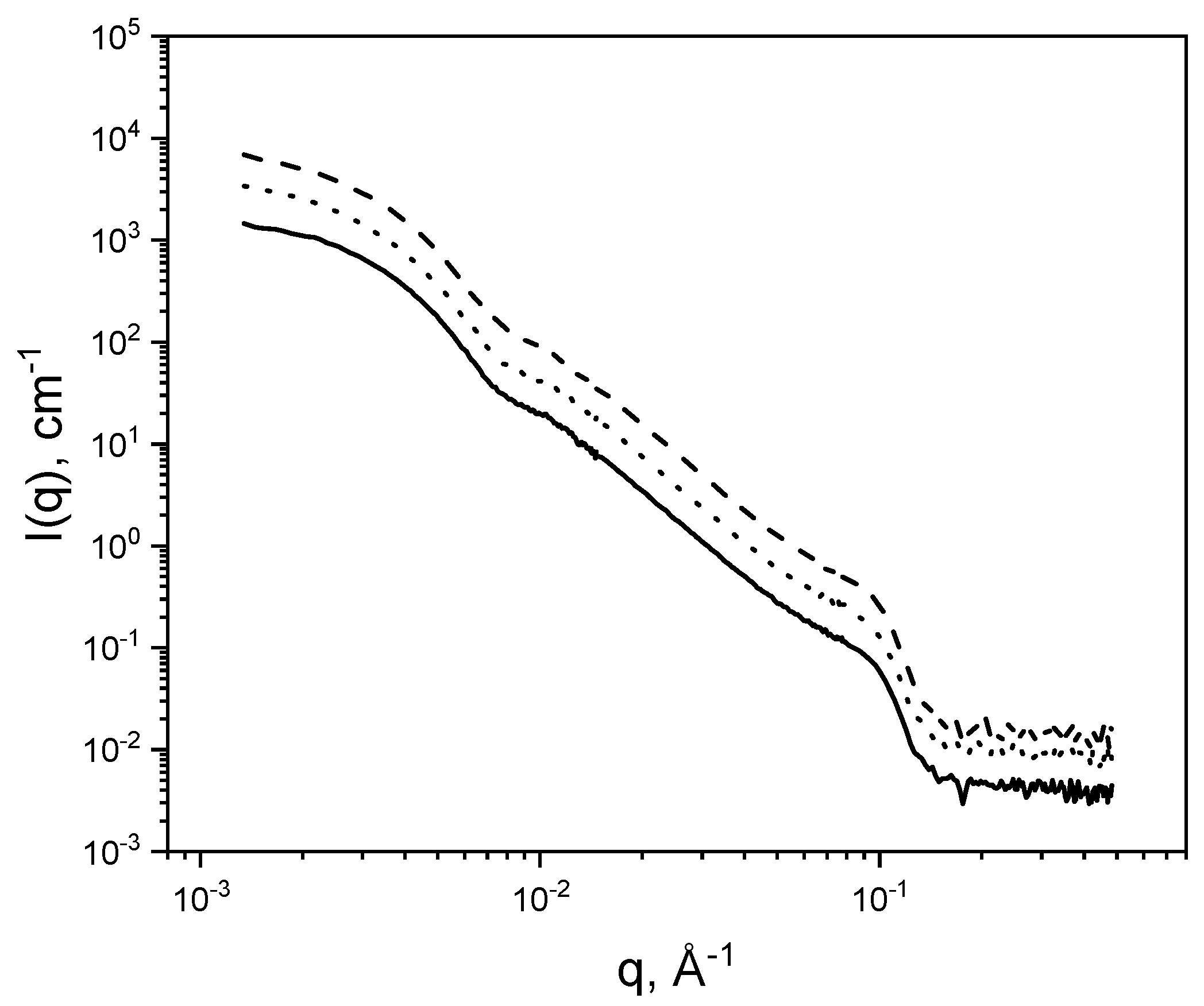
SANS experiments were performed at the D11 instrument of the Institute Laue Langevin (Grenoble, France) [
59]. Three q-ranges were explored and merged using the following wavelengths, λ, and sample-to-detector (SD) distances. The q-ranges were (1) low-q: λ = 6 Å, SD = 39 m; (2) mid-q: λ = 6 Å, SD = 8 m; (3) high-q: λ = 6 Å, SD = 1.2 m. Solutions and backgrounds were examined in standard 1 mm quartz cells. Direct beam, empty cell, and Teflon were recorded and boron carbide (B
4C) was used as neutron absorber. All samples were kept at 50 °C (above the phase transition temperature of the phospholipid) during the measurements. The scattering from the background sample was subtracted from the experimental data. Absolute values of the scattering intensity were obtained from the direct determination of the number of neutrons in the incident beam and the detector cell solid angle. The 2-D raw data were corrected for the ambient background and empty cell scattering and normalized to yield an absolute scale (cross section per unit volume) by the neutron flux on the samples. The data were then circularly averaged to yield the 1-D intensity distribution, I(q). Samples were prepared in 150 mM NaCl/D
2O (99.9%) to limit the incoherent background scattering. The concentration of DPPC and HA (M
W = 10 kDa and M
W = 1500 kDa) was the same and equal to 2 mg/mL. The full set of experimental data can be found in [
60].
4.5. Electrophoretic Experiments
Electrophoretic mobility measurements were performed at 25 °C, employing a Zetasizer 2000 instrument (Malvern Instruments, Malvern, UK). Each sample was measured 15 times, and the mean value for three independent samples is reported.
4.6. MD Simulations
All atom molecular dynamics simulations were performed using the AMBER03 force field [
24]. The HA structure was downloaded from PubChem [
26] and modified to obtain longer chains by using the YASARA (Yet Another Scientific Artificial Reality Application) Structure Software (YASARA Biosciences GmbH, Vienna, Austria). The final molecular mass of HA considered in this study was 3 kDa. The DPPC bilayer contained 288 lipid molecules. After allowing the bilayer to equilibrate, the HA molecule was placed next to the bilayer without influencing the bilayer structure. Periodic boundary conditions were applied to create an “infinite” bilayer. The TIP3P water model was used [
28]. For the isobaric–isothermal ensemble, all atom simulations were performed under the same conditions: temperatures: 298, 310 (physiological) and 323 K at a pH = 7.0 in 150 mM NaCl and CaCl
2 water solutions at a pressure of 1 bar. The time step was set to 2 fs. The simulation box contained water, one HA molecule, a DPPC bilayer, 56 cations and the corresponding number of anions to achieve electro neutrality. Simulations were carried out for 20 ns. The Berendsen barostat [
29] and thermostat with a relaxation time of 1 ps were used to maintain constant pressure and temperature. Electron density profile was calculated by using the Visual Molecular Dynamics (VMD) tool Electron Density Profile [
61]. The definition of hydrophobic contacts and hydrogen bonds, as stated in previous studies, was used [
44,
45]. Hydrogen bonds, ion-DPPC/HA contacts and HA–DPPC contacts have been extracted from the equilibrated state. For this, the data were sampled every 0.05 ns and it was checked for at which time a stable state was reached. This was typically the case between (10–20 ns).
4.7. Dynamic Light Scattering
Dynamic Light Scattering was used to obtain information about the size of the particles in solution. Therefore, it has been used to examine the interaction of lipid vesicles and HA by studying how the hydrodynamic size of the vesicles changes due to the presence of HA.
The measurements were performed using the SpectroSize 300 (Xtal concepts GmbH, Hamburg, Germany) equipped with a laser with a wavelength of 660 nm. The scattering angle was set to 90 °C and measurements were performed at 25 °C. All sample solutions were measured in the same quartz cuvette. Each sample was measured 10 times for 20 s. Six different samples (DPPC, DPPC/HA10, DPPC/HA250, DPPC/HA750, DPPC/HA1500, DPPC/HA2500) were studied at two different solution conditions (150 mM NaCl and 150 mM NaCl with 10 mM CaCl2). The concentration of DPPC and HA was 1 mg/mL, respectively.
4.8. Differential Scanning Calorimetry
Differential scanning calorimetry (DSC) is well-suited for studying phase transitions. It was used to investigate how the presence of HA changes the phase behavior of DPPC-bilayers. For the measurements, a VP-DSC MicroCalorimeter (MicroCal, Northhampton, MA, USA) was used. The instrument is equipped with two Tantaloy 61 cells (one for the sample and one for the reference). Each sample was scanned 5 times from 20 to 55 °C and five times in the inverse direction. The scanning rate was 20 °C/h for all measurements with a waiting time of 15 min between each scan. Care was taken to have the same volume (0.5 mL) in the sample and reference cell. All solutions were degassed prior to measurements to avoid air bubble formation.
Samples composed of DPPC vesicles and DPPC vesicles with HA of different molecular weights (10 kDa, 250 kDa, 750 kDa, 1500 kDa and 2500 kDa) were probed in two different salt solutions (150 mM NaCl and 150 mM NaCl with 10 mM CaCl2). The concentration of the samples was 1 mg/mL for both HA and DPPC vesicles.



 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











