
MALDI-HRMS Imaging Maps the Localization of Skyrin, the Precursor of Hypericin, and Pathway Intermediates in Leaves of Hypericum Species




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (1)
- In which plant tissues is skyrin (7) localized and accumulated?
- (2)
- In which tissues are the precursors of skyrin (7) localized and accumulated?
- (3)
- How does the spatial distribution of skyrin (7) correlate with other precursors, intermediates, and the produced naphthodianthrones?
2. Results and Discussion
2.1. Detection of the Selected Phytochemicals by HPLC-HRMS
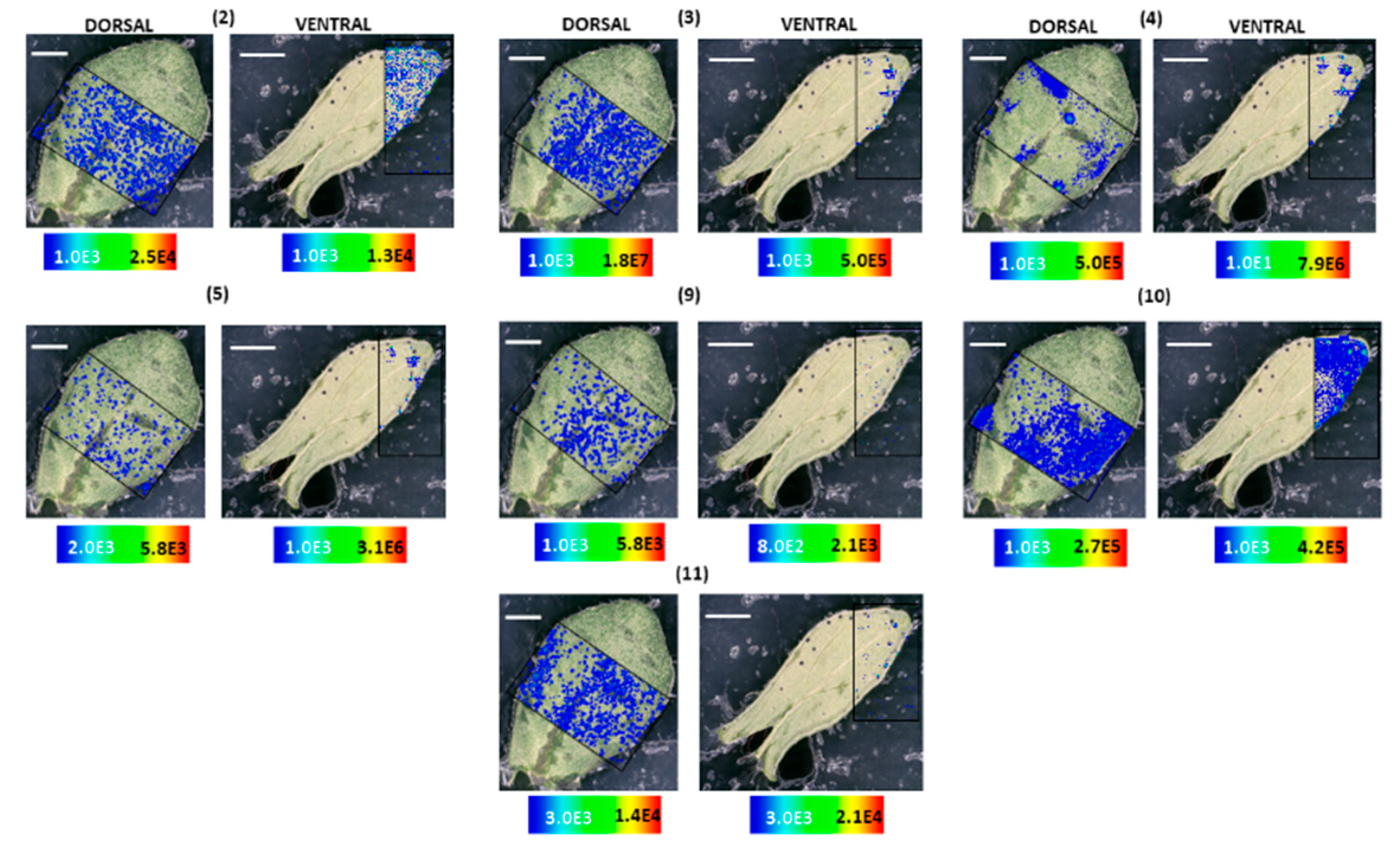
2.2. MALDI-HRMS Imaging Reveals That Skyrin Is Localized in the Dark Glands in H. humifusum and H. tetrapterum
2.3. MALDI-HRMS Imaging Reveals A Scattered Distribution of Skyrin in H. annulatum, H. bupleuroides, and H. rumeliacum
3. Materials and Methods
3.1. Plant Material and Growth Conditions
3.2. Extraction of Metabolites From Leaves
3.3. HPLC-HRMS Instrumentation and Measurement Conditions
3.4. Sample Preparation for MALDI-HRMS Imaging
3.5. MALDI-HRMS Imaging
4. Conclusions and Outlook
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Osbourn, A.; Ma, P. MYB transcription factors as regulators of Phenylpropanoid metabolism in plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheynier, V.; Tomas-Barberan, F.A.; Yoshida, K. Polyphenols: From plants to a variety of food and nonfood uses. J. Agric. Food. Chem. 2015, 63, 7589–7594. [Google Scholar] [CrossRef] [PubMed]
- Crockett, S.L.; Robson, N.K. Taxonomy and chemotaxonomy of the genus Hypericum. Med. Aromat. Plant Sci. Biotechnol. 2011, 5, 1–13. [Google Scholar] [PubMed]
- Wölfle, U.; Seelinger, G.; Schempp, C.M. Topical application of St. John’s wort (Hypericum perforatum). Planta Med. 2014, 80, 109–120. [Google Scholar]
- Kladar, N.; Mrđanović, J.; Anačkov, G.; Solajic, S.; Gavaric, N.; Srdenovic, B.; Bozin, B. Hypericum perforatum: Synthesis of active principles during flowering and Fruitification-Novel aspects of biological potential. Evid. Based Complement. Alternat. Med. 2017, 2017, 2865610. [Google Scholar] [CrossRef] [Green Version]
- Soták, M.; Czeranková, O.; Klein, D.; Jurčacková, Z.; Li, L.; Čellárová, E. Comparative Transcriptome reconstruction of four Hypericum species focused on Hypericin biosynthesis. Front. Plant Sci. 2016, 7, 1039. [Google Scholar] [CrossRef] [Green Version]
- Kusari, S.; Sezgin, S.; Nigutová, K.; Čellárová, E.; Spiteller, M. Spatial chemo-profiling of hypericin and related phytochemicals in Hypericum species using MALDI-HRMS imaging. Anal. Bioanal. Chem. 2015, 407, 4779–4791. [Google Scholar] [CrossRef]
- Hölscher, D.; Shroff, R.; Knop, K.; Gottschaldt, M.; Crecelius, A.; Schneider, B.; Heckel, D.G.; Schubert, U.S.; Svatos, A. Matrix-free UV-laser desorption/ionization (LDI) mass spectrometric imaging at the single-cell level: Distribution of secondary metabolites of Arabidopsis thaliana and Hypericum species. Plant J. 2009, 60, 907–918. [Google Scholar] [CrossRef]
- Rizzo, P.; Altschmied, L.; Stark, P.; Rutten, T.; Gündel, A.; Scharfenberg, S.; Franke, K.; Bäumlein, H.; Wessjohann, L.; Koch, M.; et al. Discovery of key regulators of dark gland development and hypericin biosynthesis in St. John’s Wort (Hypericum perforatum). Plant Biotechnol. J. 2019, 17, 2299–2312. [Google Scholar] [CrossRef] [Green Version]
- Jendželovská, Z.; Jendželovský, R.; Kuchárová, B.; Fedoročko, P. Hypericin in the light and in the dark: Two sides of the same coin. Front. Plant Sci. 2016, 7, 560. [Google Scholar] [CrossRef] [Green Version]
- Bais, H.P.; Vepachedu, R.; Lawrence, C.B.; Stermitz, F.R.; Vivanco, J.M. Molecular and biochemical characterization of an enzyme responsible for the formation of hypericin in St. John’s wort (Hypericum perforatum L.). J. Biol. Chem. 2003, 278, 32413–32422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Košuth, J.; Smelcerovic, A.; Borsch, T.; Zuehlke, S.; Karppinen, K.; Spiteller, M.; Hohtola, A.; Čellárová, E. The hyp-1 gene is not a limiting factor for hypericin biosynthesis in the genus Hypericum. Funct. Plant Biol. 2010, 38, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Košuth, J.; Katkovčinová, Z.; Olexová, P.; Čellárová, E. Expression of the hyp-1 gene in early stages of development of Hypericum perforatum L. Plant Cell Rep. 2007, 26, 211–217. [Google Scholar] [CrossRef]
- Karppinen, K.; Derzsó, E.; Jaakola, L.; Hohtola, A. Molecular cloning and expression analysis of hyp-1 Type PR-10 family genes in Hypericum perforatum. Front. Plant Sci. 2016, 7, 526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalska, K.; Fernandes, H.; Sikorski, M.; Jaskolski, M. Crystal structure of Hyp-1, a St. John’s wort protein implicated in the biosynthesis of hypericin. J. Struct. Biol. 2010, 169, 161–171. [Google Scholar] [CrossRef]
- Sliwiak, J.; Dauter, Z.; Jaskolski, M. Crystal Structure of Hyp-1, a Hypericum perforatum PR-10 Protein, in Complex with Melatonin. Front Plant Sci. 2016, 7, 668. [Google Scholar] [CrossRef] [Green Version]
- Kimáková, K.; Kimáková, A.; Idkowiak, J.; Stobiecki, M.; Rodziewicz, P.; Marczak, L.; Čellárová, E. Phenotyping the genus Hypericum by secondary metabolite profiling: Emodin vs. skyrin, two possible key intermediates in hypericin biosynthesis. Anal. Bioanal. Chem. 2018, 410, 7689–7699. [Google Scholar] [CrossRef] [Green Version]
- Hoelzl, J.; Petersen, M. Chemical constituents of Hypericum spp. Med. Aromat. Plants Ind. Profiles 2003, 31, 77–93. [Google Scholar]
- Bigelis, R.; He, H.; Yang, H.Y.; Chang, L.P.; Greenstein, M. Production of fungal antibiotics using polymeric solid supports in solid-state and liquid fermentation. J. Ind. Microbiol. Biotechnol. 2006, 33, 815–826. [Google Scholar] [CrossRef]
- Jahn, L.; Schafhauser, T.; Wibberg, D.; Rückert, C.; Winkler, A.; Kulik, A.; Weber, T.; Flor, L.; Van Pée, K.L.; Kalinowski, J.; et al. Linking secondary metabolites to biosynthesis genes in the fungal endophyte Cyanodermella asteris: The anti-cancer bisanthraquinone skyrin. J. Biotechnol. 2017, 257, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Kusari, S.; Lamshöft, M.; Zühlke, S.; Spiteller, M. An endophytic fungus from Hypericum perforatum that produces hypericin. J. Nat. Prod. 2008, 71, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Vigneshwari, A.; Rakk, D.; Németh, A.; Kocsubé, S.; Kiss, N.; Csupor, D.; Papp, T.; Škrbić, B.; Vágvölgyi, C.; Szekeres, A. Host metabolite producing endophytic fungi isolated from Hypericum perforatum. PLoS ONE 2019, 14, e0217060. [Google Scholar] [CrossRef] [PubMed]
- Henzelyová, J.; Antalová, M.; Nigutová, K.; Logoida, M.; Schreiberova, A.; Kusari, S.; Cellarova, E. Isolation, Characterization and Targeted Metabolic Evaluation of Endophytic Fungi Harbored in 14 Seed-Derived Hypericum Species [published online ahead of print, 2020 Apr 15]. Planta Med. 2020. [Google Scholar] [CrossRef]
- Bálintová, M.; Bruňáková, K.; Petijová, L.; Čellárová, E. Targeted metabolomic profiling reveals interspecific variation in the genus Hypericum in response to biotic elicitors. Plant Physiol. Biochem. 2019, 135, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Kucharíková, A.; Kusari, S.; Sezgin, S.; Spiteller, M.; Čellárová, E. Occurrence and Distribution of Phytochemicals in the Leaves of 17 in vitro Cultured Hypericum spp. Adapted to Outdoor Conditions. Front. Plant Sci. 2016, 7, 1616. [Google Scholar] [CrossRef] [Green Version]
- Kucharíková, A.; Kimáková, K.; Janfelt, C.; Čellárová, E. Interspecific variation in localization of hypericins and phloroglucinols in the genus Hypericum as revealed by desorption electrospray ionization mass spectrometry imaging. Physiol. Plant. 2016, 157, 2–12. [Google Scholar] [CrossRef]
- Zobayed, S.M.; Afreen, F.; Goto, E.; Kozai, T. Plant-environment interactions: Accumulation of hypericin in dark glands of Hypericum perforatum. Ann. Bot. 2006, 98, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Soelberg, J.; Jørgensen, L.B.; Jäger, A.K. Hyperforin accumulates in the translucent glands of Hypericum perforatum. Ann. Bot. 2007, 99, 1097–1100, reprinted in Ann. Bot. (Lond.) 2007, 100, 679. [Google Scholar] [CrossRef] [Green Version]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef]
- Izhaki, I. Emodin—A secondary metabolite with multiple ecological functions in higher plants. New Phytol. 2002, 155, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plantarum. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Nigutová, K.; Kusari, S.; Sezgin, S.; Petijová, L.; Henzelyová, J.; Bálintová, M.; Spiteller, M.; Čellárová, E. Chemometric evaluation of hypericin and related phytochemicals in 17 in vitro cultured Hypericum species, hairy root cultures and hairy root-derived transgenic plants. J. Pharm. Pharmacol. 2019, 71, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.S.; Fang, X.W.; Huang, R.; Zhang, S.P.; Wei, H.X.; Wu, S.H. A new dimeric anthraquinone from endophytic Talaromyces sp. YE3016. Nat. Prod. Res. 2016, 30, 1706–1711. [Google Scholar] [CrossRef]
- Kusari, S.; Lamshöft, M.; Kusari, P.; Gottfried, S.; Zühlke, S.; Louven, K.; Hentschel, U.; Kayser, O.; Spiteller, M. Endophytes are hidden producers of maytansine in Putterlickia roots. J. Nat. Prod. 2014, 77, 2577–2584. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the Hypericum plants are available from the group of E.C. upon request. |









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Revuru, B.; Bálintová, M.; Henzelyová, J.; Čellárová, E.; Kusari, S. MALDI-HRMS Imaging Maps the Localization of Skyrin, the Precursor of Hypericin, and Pathway Intermediates in Leaves of Hypericum Species. Molecules 2020, 25, 3964. https://doi.org/10.3390/molecules25173964
Revuru B, Bálintová M, Henzelyová J, Čellárová E, Kusari S. MALDI-HRMS Imaging Maps the Localization of Skyrin, the Precursor of Hypericin, and Pathway Intermediates in Leaves of Hypericum Species. Molecules. 2020; 25(17):3964. https://doi.org/10.3390/molecules25173964
Chicago/Turabian StyleRevuru, Bharadwaj, Miroslava Bálintová, Jana Henzelyová, Eva Čellárová, and Souvik Kusari. 2020. "MALDI-HRMS Imaging Maps the Localization of Skyrin, the Precursor of Hypericin, and Pathway Intermediates in Leaves of Hypericum Species" Molecules 25, no. 17: 3964. https://doi.org/10.3390/molecules25173964
APA StyleRevuru, B., Bálintová, M., Henzelyová, J., Čellárová, E., & Kusari, S. (2020). MALDI-HRMS Imaging Maps the Localization of Skyrin, the Precursor of Hypericin, and Pathway Intermediates in Leaves of Hypericum Species. Molecules, 25(17), 3964. https://doi.org/10.3390/molecules25173964