
Simplified LC-MS Method for Analysis of Sterols in Biological Samples



Abstract
:1. Introduction
2. Results and Discussion
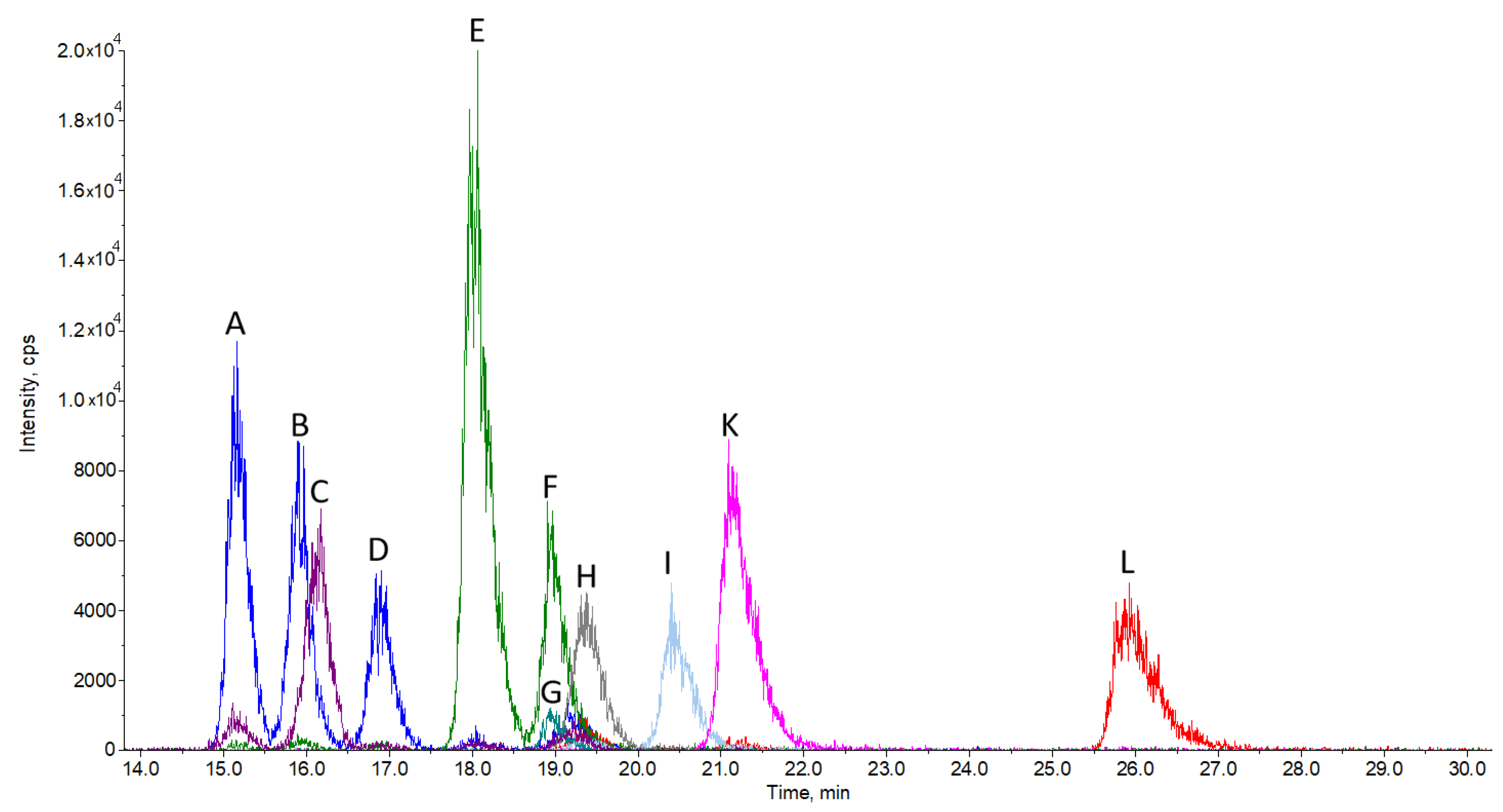
2.1. Chromatographic Separation and Detection
2.2. Sample Analysis
3. Materials and Methods
3.1. Materials
3.2. Sample Preparation
3.2.1. Isolation and Culture of Human Hepatocytes
3.2.2. Sterol Isolation
3.3. LC-MS Analysis
3.4. Quantitation, Accuracy, Repeatability, Linearity, and Stability
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Simons, K.; Ikonen, E. How cells handle cholesterol. Science 2000, 290, 1721–1726. [Google Scholar] [CrossRef] [Green Version]
- Cerqueira, N.M.; Oliveira, E.F.; Gesto, D.S.; Santos-Martins, D.; Moreira, C.; Moorthy, H.N.; Ramos, M.J.; Fernandes, P.A. Cholesterol biosynthesis: A mechanistic overview. Biochemistry 2016, 55, 5483–5506. [Google Scholar] [CrossRef] [PubMed]
- Ačimovič, J.; Rozman, D. Steroidal triterpenes of cholesterol synthesis. Molecules 2013, 18, 4002–4017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsche, M.A.; McDonald, J.G.; Hobbs, H.H.; Cohen, J.C. Flux analysis of cholesterol biosynthesis in vivo reveals multiple tissue and cell-type specific pathways. eLife 2015, 4, 1–21. [Google Scholar] [CrossRef]
- Belič, A.; Pompon, D.; Monostory, K.; Kelly, D.; Kelly, S.; Rozman, D. An algorithm for rapid computational construction of metabolic networks: A cholesterol biosynthesis example. Comput. Biol. Med. 2013, 43, 471–480. [Google Scholar] [CrossRef]
- Skubic, C.; Rozman, D. Sterols from the post-lanosterol part of cholesterol synthesis: Novel signaling players. In Mammalian Sterols: Novel Biological Roles of Cholesterol Synthesis Intermediates, Oxysterols and Bile Acids; Rozman, D., Gebhardt, R., Eds.; Springer International Publishing: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Porter, F.D.; Herman, G.E. Malformation syndromes caused by disorders of cholesterol synthesis. J. Lipid Res. 2011, 52, 6–34. [Google Scholar] [CrossRef] [Green Version]
- Hubler, Z.; Allimuthu, D.; Bederman, I.; Elitt, M.S.; Madhavan, M.; Allan, K.C.; Shick, H.E.; Garrison, E.; Karl, M.T.; Factor, D.C.; et al. Accumulation of 8,9-unsaturated sterols drives oligodendrocyte formation and remyelination. Nature 2018, 560, 372–376. [Google Scholar] [CrossRef]
- Santori, F.R.; Huang, P.; Van De Pavert, S.A.; Douglass, E.F.; Leaver, D.J.; Haubrich, B.A.; Keber, R.; Lorbek, G.; Konijn, T.; Rosales, B.N.; et al. Identification of natural ligands that regulate the development of lymphoid cells. Cell Metab. 2015, 21, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Ma, M.Y.; Sun, M.; Jiang, L.Y.; Zhao, X.T.; Fang, X.X.; Lam, S.M.; Shui, G.H.; Luo, J.; Shi, X.J.; et al. Endogenous sterol intermediates of the mevalonate pathway regulate HMGCR degradation and SREBP-2 processing. J. Lipid Res. 2019, 60, 1765–1775. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Abdel-Khalik, J.; Yutuc, E.; Morgan, A.H.; Gilmore, I.; Hearn, T.; Wang, Y. Cholesterolomics: An update. Anal. Biochem. 2017, 524, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.C.; Axelsen, P.H. Mass spectrometric analysis of long-chain lipids. Mass Spectrom. Rev. 2011, 30, 579–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, I.H.K.; Wilson, S.R.; Roberg-Larsen, H. Chromatography of oxysterols. Biochimie 2018, 153, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, J. Rapid measurement of free phytosterols in tobacco by short-column gc/ms/ms. j. Agric. Food Chem. 1997, 45, 3932–3935. [Google Scholar] [CrossRef]
- Toivo, J.; Piironen, V.; Kalo, P.; Varo, P. Gas chromatographic determination of major sterols in edible oils and fats using solid-phase extraction in sample preparation. Chromatographia 1998, 48, 745–750. [Google Scholar] [CrossRef]
- Acimovic, J.; Lövgren-Sandblom, A.; Monostory, K.; Rozman, D.; Golicnik, M.; Lutjohann, D.; Björkhem, I. Combined gas chromatographic/mass spectrometric analysis of cholesterol precursors and plant sterols in cultured cells. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 2081–2086. [Google Scholar] [CrossRef]
- Moreau, R.A.; Nyström, L.; Whitaker, B.D.; Winkler-Moser, J.K.; Baer, D.J.; Gebauer, S.K.; Hicks, K.B. Phytosterols and their derivatives: Structural diversity, distribution, metabolism, analysis, and health-promoting uses. Prog. Lipid Res. 2018, 70, 35–61. [Google Scholar] [CrossRef]
- Baila-Rueda, L.; Cenarro, A.; Cofán, M.; Orera, I.; Barcelo-Batllori, S.; Pocoví, M.; Ros, E.; Civeira, F.; Nerín, C.; Domeno, C. Simultaneous determination of oxysterols, phytosterols and cholesterol precursors by high performance liquid chromatography tandem mass spectrometry in human serum. Anal. Methods 2013, 5, 2249–2257. [Google Scholar] [CrossRef]
- Cañabate-Díaz, B.; Segura Carretero, A.; Fernández-Gutiérrez, A.; Belmonte Vega, A.; Garrido Frenich, A.; Martínez Vidal, J.L.; Duran Martos, J. Separation and determination of sterols in olive oil by HPLC-MS. Food Chem. 2007, 102, 593–598. [Google Scholar] [CrossRef]
- Rocco, A.; Fanali, S. Analysis of phytosterols in extra-virgin olive oil by nano-liquid chromatography. J. Chromatogr. A 2009, 1216, 7173–7178. [Google Scholar] [CrossRef]
- Carretero, A.S.; Carrasco-Pancorbo, A.; Cortacero, S.; Gori, A.; Cerretani, L.; Fernández-Gutiérrez, A. A simplified method for HPLC-MS analysis of sterols in vegetable oil. Eur. J. Lipid Sci. Technol. 2008, 110, 1142–1149. [Google Scholar] [CrossRef]
- Kim, D.; Park, J.B.; Choi, W.-K.; Lee, S.J.; Lim, I.; Bae, S.K. Simultaneous determination of β-sitosterol, campesterol, and stigmasterol in rat plasma by using LC-APCI-MS/MS: Application in a pharmacokinetic study of a titrated extract of the unsaponifiable fraction of Zea mays L. J. Sep. Sci. 2016, 39, 4060–4070. [Google Scholar] [CrossRef]
- McDonald, J.G.; Smith, D.D.; Stiles, A.R.; Russell, D.W. A comprehensive method for extraction and quantitative analysis of sterols and secosteroids from human plasma. J. Lipid Res. 2012, 53, 1399–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudin, M.; Imbert, L.; Libong, D.; Chaminade, P.; Brunelle, A.; Touboul, D.; Laprévote, O. Atmospheric pressure photoionization as a powerful tool for large-scale lipidomic studies. J. Am. Soc. Mass Spectrom. 2012, 23, 869–879. [Google Scholar] [CrossRef] [Green Version]
- Abidi, S.L. Chromatographic analysis of plant sterols in foods and vegetable oils. J. Chromatogr. A 2001, 935, 173–201. [Google Scholar] [CrossRef]
- McDonald, J.G.; Thompson, B.M.; McCrum, E.C.; Russell, D.W. Extraction and analysis of sterols in biological matrices by high performance liquid chromatography electrospray ionization mass spectrometry. Methods Enzymol. 2007, 432, 145–170. [Google Scholar] [CrossRef]
- Marchand, D.H.; Croes, K.; Dolan, J.W.; Snyder, L.R.; Henry, R.A.; Kallury, K.M.R.; Waite, S.; Carr, P.W. Column selectivity in reversed-phase liquid chromatography. VIII. Phenylalkyl and fluoro-substituted columns. J. Chromatogr. A 2005, 1062, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Honda, A.; Miyazaki, T.; Ikegami, T.; Iwamoto, J.; Yamashita, K.; Numazawa, M.; Matsuzaki, Y. Highly sensitive and specific analysis of sterol profiles in biological samples by HPLC-ESI-MS/MS. J. Steroid Biochem. Mol. Biol. 2010, 121, 556–564. [Google Scholar] [CrossRef]
- Lorbek, G.; Perše, M.; Horvat, S.; Björkhem, I.; Rozman, D. Sex differences in the hepatic cholesterol sensing mechanisms in mice. Molecules 2013, 18, 11067–11085. [Google Scholar] [CrossRef] [Green Version]
- Urlep, Ž.; Lorbek, G.; Perše, M.; Jeruc, J.; Juvan, P.; Matz-Soja, M.; Gebhardt, R.; Björkhem, I.; Hall, J.A.; Bonneau, R.; et al. Disrupting hepatocyte Cyp51 from cholesterol synthesis leads to progressive liver injury in the developing mouse and decreases RORC signalling. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds sterol standards available at Avanti Polar Lipids (Alabaster, AL, USA) and cell line samples are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Sterols | Accuracy (%) | Repeatability (% RSD) | Regression Coefficient | Stability 48 h (%) | LOQ (ng/mL) | LOD (ng/mL) | S/N (125 ng/mL) |
|---|---|---|---|---|---|---|---|
| Zymosterol | 105.0 | 7.5 | 0.9999 | 86.6 | 90.6 | 27.2 | 13.8 |
| Dihydrolanosterol | 97.9 | 4.2 | 0.9999 | 96.2 | 148.8 | 44.6 | 8.4 |
| Zymostenol | 106.4 | 3.5 | 0.9999 | 95.2 | 78.1 | 23.4 | 16.0 |
| FFMAS | 86.5 | 6.1 | 0.9999 | 82.5 | 178.5 | 53.6 | 7.0 |
| TMAS | 87.2 | 4.3 | 0.9998 | 82.2 | 187.7 | 56.3 | 6.7 |
| Lanosterol | 95.7 | 5.8 | 0.9998 | 92.5 | 115.7 | 34.7 | 10.8 |
| 7-dehydrodesmosterol | 109.7 | 7.4 | 0.9996 | 88.3 | 152.4 | 45.7 | 8.2 |
| 24-dehydrolathosterol | 103.1 | 9.4 | 0.9997 | 93.0 | 96.9 | 29.1 | 12.9 |
| Desmosterol | 96.1 | 4.5 | 0.9997 | 85.4 | 151.5 | 45.5 | 8.3 |
| Lathosterol | 101.2 | 5.2 | 0.9997 | 89.4 | 183.8 | 55.1 | 6.8 |
| Sterol Measured | HepG2 n = 3 | HepG2 DHCR24 KO n = 3 | Statistical Significance with Unpaired t-Test * | ||
|---|---|---|---|---|---|
| Concentration | |||||
| Mean (ng/mL) | SD | Mean (ng/mL) | SD | ||
| Zymosterol | 192.8 | 11.7 | 2446.1 | 707.1 | *** |
| Dihydrolanosterol | 38.1 | 3.0 | 0.0 | 0.0 | **** |
| Zymostenol | 365.5 | 38.1 | 25.8 | 23.5 | ** |
| T-MAS | 148.5 | 7.6 | 1535.9 | 335.6 | ** |
| Lanosterol | 73.1 | 3.8 | 305.9 | 65.1 | ** |
| 7-dehydrodesmosterol | 36.0 | 2.3 | 1411.6 | 236.3 | *** |
| 24-dehydrolathosterol | 57.0 | 3.2 | 1414.0 | 236.8 | *** |
| Desmosterol | 248.7 | 19.8 | 108156.4 | 19170.1 | *** |
| Lathosterol | 842.9 | 152.4 | 0.0 | 0.0 | *** |
| Trivial Name | Avanti Polar Lipids Number * | Chemical Name | Molar Mass | MRM * | tR (min) |
|---|---|---|---|---|---|
| Lanosterol | 700063P | 8,24-lanostadien-3β-ol | 426.72 | 409/191 | 21.4 |
| Dihydro lanosterol | 700067P | 24,25-dihydrolanosterol | 428.73 | 411/191 | 26.3 |
| FFMAS | 700077P | 14-demethyl-14-dehydrolanosterol | 410.68 | 393/214 | 19.5 |
| TMAS | 700073P | 4,4-dimethylcholest-8(9),24-dien-3β-ol | 412.69 | 395/243 | 20.7 |
| Zymosterol | 700068P | 5α-cholesta-8,24-dien- 3β-ol | 384.64 | 367/215 | 15.3 |
| Zymostenol | 700118P | 5α-cholest-8-en-3β-ol | 386.65 | 369/215 | 18.2 |
| 24-dehydro lathosterol | 700114P | 5α-cholesta-7,24-dien- 3β-ol | 384.64 | 367/215 | 16.0 |
| lathosterol | 700069P | 5α-cholest-7-en-3β-ol | 386.65 | 369/215 | 19.1 |
| 7-dehydro desmosterol | 700138P | 7-dehydrodesmosterol | 382.62 | 365/199 | 16.3 |
| Desmosterol | 700060P | 3β-hydroxy-5,24-cholestadiene | 384.64 | 367/215 | 16.9 |
| Lathosterol-d7 | 700056P | 5α-Cholest-7-en-3β-ol(25,26,26,26,27,27,27-d7) | 393.70 | 376/215 | 19.2 |
| Cholesterol | Merck (C3045) | Cholest-5-en-3β-ol | 386.70 | 369/215 | 19.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skubic, C.; Vovk, I.; Rozman, D.; Križman, M. Simplified LC-MS Method for Analysis of Sterols in Biological Samples. Molecules 2020, 25, 4116. https://doi.org/10.3390/molecules25184116
Skubic C, Vovk I, Rozman D, Križman M. Simplified LC-MS Method for Analysis of Sterols in Biological Samples. Molecules. 2020; 25(18):4116. https://doi.org/10.3390/molecules25184116
Chicago/Turabian StyleSkubic, Cene, Irena Vovk, Damjana Rozman, and Mitja Križman. 2020. "Simplified LC-MS Method for Analysis of Sterols in Biological Samples" Molecules 25, no. 18: 4116. https://doi.org/10.3390/molecules25184116
APA StyleSkubic, C., Vovk, I., Rozman, D., & Križman, M. (2020). Simplified LC-MS Method for Analysis of Sterols in Biological Samples. Molecules, 25(18), 4116. https://doi.org/10.3390/molecules25184116