
Glucose-Uptake Activity and Cytotoxicity of Diterpenes and Triterpenes Isolated from Lamiaceae Plant Species

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
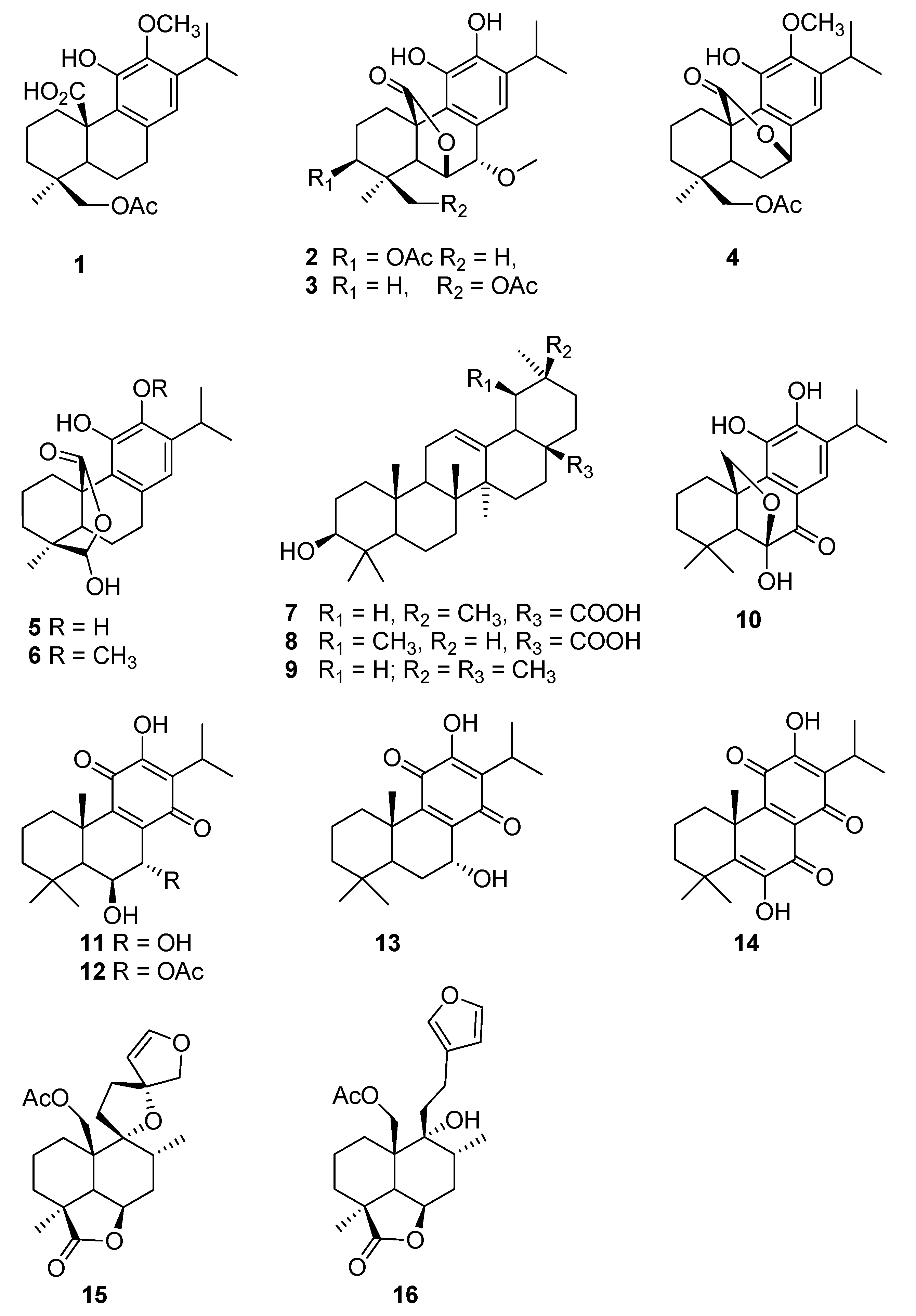
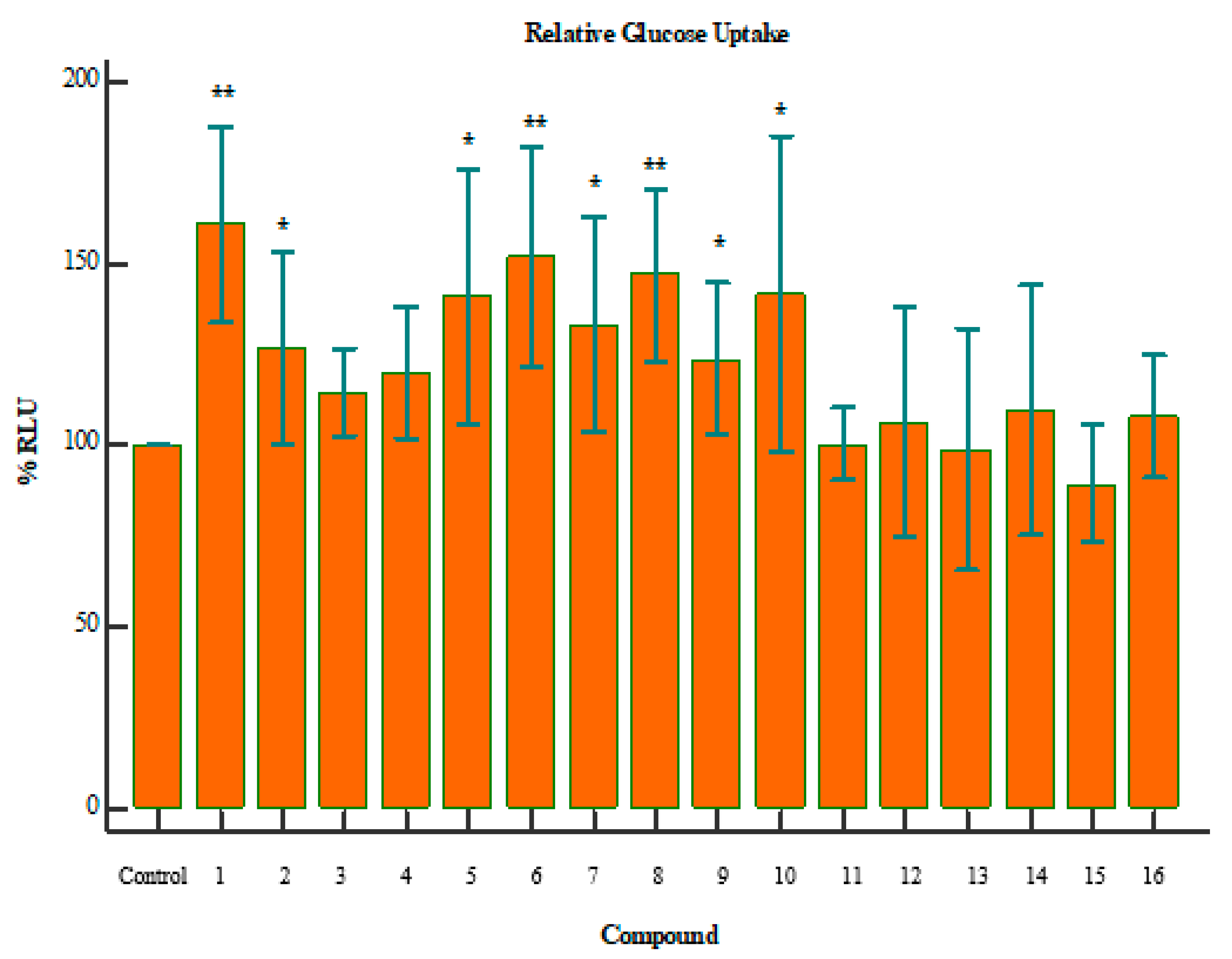
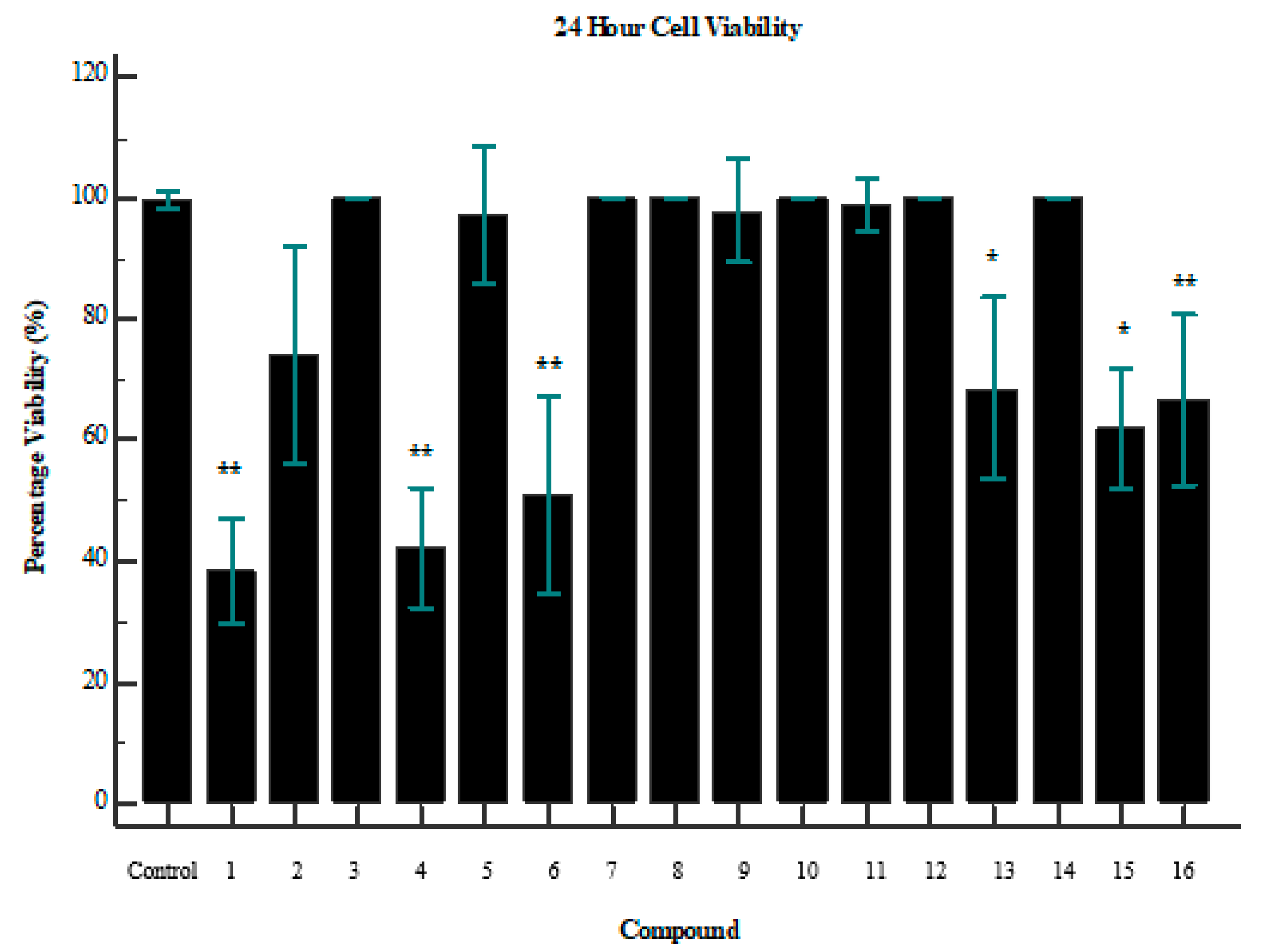
2. Results
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
4.3. Extraction and Purification of Chemical Constituents
4.4. Spectroscopic Data of Compounds 10–16
- Compound 10.1H-NMR (400 MHz, CDCl3), δH 7.65 (s, H-14), 4.29 (d, J = 7.5 Hz, Hβ-20), 3.37 (d, J = 7.5 Hz, Hα-20), 3.02, (sept, J = 7.1 Hz, H-15), 1.31 (s, Me-19), 1.17 (d, J = 7.1 Hz, H-16), 1.16 (d, J = 7.1 Hz, H-17), 1.04 (s, Me-18). 13C-NMR (100 MHz, CDCl3), δC 192.8 (C-7), 148.3 (C-12), 140.5 (C-11), 137.7 (C-9), 133.3 (C-13), 120.1 (C-14), 121.4 (C-8), 105.2 (C-6), 72.0 (C-20), 58.2 (C-5), 51.47 (C-10), 41.33 (C-3), 33.7 (C-18), 32.4 (C-4), 29.6 (C-1), 27.1 (C-15), 22.5 (C-17), 22.4 (C-16), 22.2 (C-19), 18.5 (C-2).
- Compound 11.1H-NMR (400 MHz, CDCl3), δH 4.53 (d, J = 1.5 Hz, H-7), 4.46 (br s, H-6), 3.18 (sept, J = 7.1 Hz, H-15), 1.62 (s, Me-20), 1.27 (s, Me-19), 1.23 (d, J = 7.1 Hz, H-16), 1.23 (d, J = 7.1 Hz, H-17), 1.06 (s, Me-18). 13C-NMR (100 MHz, CDCl3), δC 189.5 (C-14), 183.1 (C-11), 151.2 (C-12), 147.6 (C-9), 140.9 (C-8), 124.3 (C-13), 69.3 (C-6), 69.1 (C-7), 49.5 (C-5), 42.3 (C-3), 38.6 (C-10), 38.4 (C-1), 33.7 (C-4), 33.5 (C-18), 24.3 (C-19), 24.0 (C-15), 21.6 (C-20), 19.9 (C-17), 19.8 (C-16), 19.0 (C-2).
- Compound 12.1H-NMR (400 MHz, CDCl3), δH 5.60 (d, J = 1.8 Hz, H-7), 4.24 (s, H-6), 3.09 (sept, J = 7.1 Hz, H-15), 1.98 (s, CH3CO), 1.55 (s, Me-20), 1.16 (s, Me-19), 1.13 (d, J = 7.1 Hz, H-16), 1.11 (d, J = 7.1 Hz, H-17), 0.92 (s, Me-18). 13C-NMR, δC (100 MHz, CDCl3) 186.0 (C-14), 183.1 (C-11), 151.2 (C-12), 150.1 (C-9), 137.0 (C-8), 124.3 (C-13), 69.0 (C-7), 66.4 (C-6), 49.7 (C-5), 42.33 (C-3) 38.6 (C-4) 38.3 (C-1), 33.6 (C-10), 33.5 (C-18), 18.9 (C-2), 24.1 (C-15), 23.6 (C-19), 21.3 (C-20), 19.8 (C-17), 19.6 (C-16), 21.0/170.1 (CH3CO).
- Compound 13.1H-NMR (400 MHz, CDCl3), δH 4.73 (d, J = 1.5 Hz, H-7), 3.16 (sept, J = 7.1 Hz, H-15), 1.22 (s, Me-20), 1.21 (d, J = 7.1Hz, H-16), 1.20 (d, J = 7.1 Hz, H-17), 0.98 (s, Me-18), 0.90 (s, Me-19). 13C-NMR (100 MHz, CDCl3) δC 189.2 (C-14), 183.9 (C-11), 151.1 (C-12), 147.8 (C-9), 143.3 (C-8), 124.2 (C-13), 63.2 (C-7), 45.8 (C-5), 41.13 (C-3), 39.8 (C-10), 35.8 (C-1), 33.2 (C-4), 33.1 (C-18), 25.8 (C-6), 24.0 (C-15), 21.7 (C-19), 19.9 (C-16), 19.8 (C-17), 19.0 (C-2), 18.4 (C-20).
- Compound 14.1H-NMR (400 MHz, CDCl3), δH 3.22 (sept, J = 7.0 Hz, H-15), 1.64 (s, Me-20), 1.43 (s, Me-18), 1.42 (s, Me-19), 1.25 (d, J = 7.1 Hz, H-16), 1.24 (d, J = 7.0 Hz, H-17). 13C-NMR (100 MHz, CDCl3), δC 184.3 (C-14), 183.6 (C-11), 177.5 (C-7), 155.1 (C-9), 150.7 (C-12), 146.8 (C-6), 143.3 (C-5), 126.8 (C-8), 126.0 (C-13), 41.4 (C-10), 36.4 (C-4), 36.3 (C-3), 30.8 (C-1), 29.1 (C-19), 27.5 (C-20), 27.2 (C-18), 24.4 (C-15), 19.8 (C-16), 19.8 (C-17), 17.7 (C-2).
- Compound 15.1H-NMR (200 MHz, CDCl3), δH 6.34 (br d, J= 2.6 Hz, H-15), 5.04 (br d, J= 2.6 Hz, H-14), 4.59 (br t, J= 4.0 Hz, H-6), 4.26 and 4.17 (d each, J= 12.5 Hz, CH2-20), 4.28 and 3.95 (d, each, J = 10.6 Hz, H-16a, b), 1.12 (s, Me-18), 0.73 (d, J = 6.4 Hz, Me-17). 13C-NMR (50 MHz CDCl3) δC 182.3 (C-19), 147.9 (C-15), 107.0 (C-14), 91.3 (C-13), 88.7 (C-9), 79.7 (C-16), 75.5 (C-6), 65.6 (C-20), 46.4 (C-5), 43.2 (C-4), 42.0 (C-10), 37.0 (C-12), 32.4 (C-8), 31.5 (C-7), 31.4 (C-11), 28.0 (C-3), 23.4 (C-18), 22.2 (C-1), 17.6 (C-2), 17.4 (C-17), 20.7/169.8 (COCH3).
- Compound 16.1H-NMR (200 MHz, CDCl3) δH 7.34 (br s, H15), 7.21 (br s, H-16), 6.24 (br s, H-14), 5.13 (br s, H-6), 4.27/4.62 (br d each J= 12.3 Hz, CH2-20), 1.01 (s, Me-18), 0.96 (d, J = 6.3 Hz, Me-17). 13C-NMR (50 MHz, CDCl3) δC 175.9 (C-19), 143.4 (C-15), 138.4 (C-16), 124.2 (C-13), 110.3 (C-14), 75.8 (C-20), 74.7 (C-9), 69.3 (C-6), 46.5 (C-5), 43.9 (C-11), 41.0 (C-10), 40.8 (C-4), 39.6 (C-3), 33.3 (C-8), 30.1 (C-1), 32.8 (C-7), 22.3 (C-18), 22.4 (C-2), 20.3 (C-12), 15.6 (C-17), 20.9/170.2 (COCH3).
4.5. Glucose-Uptake Assay
4.6. Cytotoxicity Assay (MTT)
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tripathy, J.P. Burden and risk factors of diabetes and hyperglycemia in India: Findings from the Global Burden of Disease Study 2016. Diabetes Metab. Syndr. Obes. Targets Ther. 2018, 11, 381–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFronzo, R.A.; Tripathy, D. Skeletal Muscle Insulin Resistance Is the Primary Defect in Type 2 Diabetes. Diabetes Care 2009, 32, S157–S163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilcox, G. Insulin and insulin resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar] [PubMed]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zúñiga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef]
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czech, M.P. Insulin action and resistance in obesity and type 2 diabetes. Nat. Med. 2017, 23, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Dal, S.; Sigrist, S. The Protective Effect of Antioxidants Consumption on Diabetes and Vascular Complications. Diseases 2016, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Etsassala, N.G.; Cupido, C.N.; Iwuoha, E.; Hussein, A.A. Review: Abietane diterpenes as potential candidates for the management of type 2 diabetes. Curr. Pharm. Des. 2020, 26, 1–13. [Google Scholar] [CrossRef]
- Alemu, A.; Tamiru, W.; Nedi, T.; Shibeshi, W. Analgesic and Anti-Inflammatory Effects of 80% Methanol Extract of Leonotis ocymifolia (Burm.f.) Iwarsson Leaves in Rodent Models. Evidence-Based Complement. Altern. Med. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kubinova, R.; Pořízková, R.; Navrátilová, A.; Farsa, O.; Hanaková, Z.; Bačinská, A.; Cizek, A.; Valentová, M. Antimicrobial and enzyme inhibitory activities of the constituents of Plectranthus madagascariensis (Pers.) Benth. J. Enzym. Inhib. Med. Chem. 2014, 29, 749–752. [Google Scholar] [CrossRef]
- Yin, Z.; Zhang, W.; Feng, F.; Zhang, Y.; Kang, W. α-Glucosidase inhibitors isolated from medicinal plants. Food Sci. Hum. Wellness 2014, 3, 136–174. [Google Scholar] [CrossRef] [Green Version]
- Manning, J.; Goldblatt, P. Plants of the Greater Cape Floristic Region 1: The Core Cape Flora; South African National Biodiversity Institute: Pretoria, South Africa, 2012; ISBN 9781919976747. [Google Scholar]
- He, Y.-H.; Li, W.; Li, Y.; Zhang, S.; Wang, Y.; Sun, C. Ursolic Acid Increases Glucose Uptake through the PI3K Signaling Pathway in Adipocytes. PLoS ONE 2014, 9, e110711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamede, M.; Mabuza, L.P.; Ngubane, P.; Khathi, A. The Effects of Plant-Derived Oleanolic Acid on Selected Parameters of Glucose Homeostasis in a Diet-Induced Pre-Diabetic Rat Model. Molecules 2018, 23, 794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teoh, S.L.; Das, S. Phytochemicals and their effective role in the treatment of diabetes mellitus: A short review. Phytochem. Rev. 2018, 17, 1111–1128. [Google Scholar] [CrossRef]
- Lipina, C.; Hundal, H.S. Carnosic acid stimulates glucose uptake in skeletal muscle cells via a PME-1/PP2A/PKB signalling axis. Cell. Signal. 2014, 26, 2343–2349. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Borji, A.; Farkhondeh, T. Evaluation of Antidiabetic Activity of Carnosol (Phenolic Diterpene in Rosemary) in Streptozotocin-Induced Diabetic Rats. Cardiovasc. Hematol. Disord. Targets 2017, 17, 11–17. [Google Scholar] [CrossRef]
- Vlavcheski, F.; Baron, D.; Vlachogiannis, I.A.; MacPherson, R.E.K.; Tsiani, E. Carnosol Increases Skeletal Muscle Cell Glucose Uptake via AMPK-Dependent GLUT4 Glucose Transporter Translocation. Int. J. Mol. Sci. 2018, 19, 1321. [Google Scholar] [CrossRef] [Green Version]
- Etsassala, N.G.; Badmus, J.; Waryo, T.; Marnewick, J.L.; Cupido, C.N.; Hussein, A.A.; Iwuoha, E.I. Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities of Novel Abietane Diterpenes from Salvia africana-lutea. Antioxidants 2019, 8, 421. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.-B.; Yang, S.-J.; Yan, Z.-R.; Zhang, X.-J.; Pu, D.-B.; Wang, L.-X.; Li, X.-L.; Zhang, R.-H.; Xiao, W.-L. Isolation, Characterization, and Structure–Activity Relationship Analysis of Abietane Diterpenoids from Callicarpa bodinieri as Spleen Tyrosine Kinase Inhibitors. J. Nat. Prod. 2018, 81, 998–1006. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Kang, S.C. Tyrosinase and α-Glucosidase Inhibitory Effects of an Abietane Type Diterpenoid Taxodone from Metasequoia glyptostroboides. Natl. Acad. Sci. Lett. 2015, 38, 399–402. [Google Scholar] [CrossRef]
- Batista, O.; Simões, M.F.; Nascimento, J.; Riberio, S.; Duarte, A.; Rodríguez, B.; De La Torre, M.C. A rearranged abietane diterpenoid from Plectranthus hereroensis. Phytochemistry 1996, 41, 571–573. [Google Scholar] [CrossRef]
- Hussein, A.A.; Meyer, J.J.M.; Jimeno, M.L.; Rodríguez, B. Bioactive Diterpenes from Orthosiphon labiatus and Salvia africana-lutea. J. Nat. Prod. 2007, 70, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Bustos-Brito, C.; Joseph-Nathan, P.; Burgueño-Tapia, E.; Martínez-Otero, D.; Nieto-Camacho, A.; Calzada, F.; Yépez-Mulia, L.; Esquivel, B.; Quijano, L. Structure and Absolute Configuration of Abietane Diterpenoids from Salvia clinopodioides: Antioxidant, Antiprotozoal, and Antipropulsive Activities. J. Nat. Prod. 2019, 82, 1207–1216. [Google Scholar] [CrossRef]
- Rasikari, H. Phytochemistry and Arthropod Bioactivity of Australian Lamiaceae. Ph.D. Thesis, Southern Cross University, Lismore, Australia, 2007. [Google Scholar]
- Naman, C.B. Phytochemical Investigation of the Medicinal Plant Taxodium distichum and Library Screening of Thalictrum Alkaloids for New Antileishmanial Drug Leads. Ph.D. Thesis, Ohio State University, Ohio, OH, USA, 2015. [Google Scholar]
- Matloubi-Moghadam, F.; Rüedi, P.; Eugster, C.H. Drüsefarbstoffe aus Labiaten: Identifizierung von 17 Abietanoiden ausPlectranthus sanguineusBRITTEN. Helvetica Chim. Acta 1987, 70, 975–983. [Google Scholar] [CrossRef]
- Horvath, T.; Linden, A.; Yoshizaki, F.; Eugster, C.H.; Ruedi, P. Abietanes and a Novel 20-Norabietanoid from Plectranthus cyaneus (Lamiaceae). Helvetica Chim. Acta 2004, 87, 2346–2353. [Google Scholar] [CrossRef]
- McKenzie, J.M.; Green, I.R.; Mugabo, P. Leonurun. A novel labdane diterpenoid from Leonotis leonurus. S. Afr. J. Chem. 2006, 59, 114–116. [Google Scholar]
- Habtemariam, S.; Gray, A.I.; Waterman, P.G. Diterpenes from the Leaves of Leonotis ocymifolia var. raineriana. J. Nat. Prod. 1994, 57, 1570–1574. [Google Scholar] [CrossRef]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Mahomoodally, M.F. Traditional Medicines in Africa: An Appraisal of Ten Potent African Medicinal Plants. Evidence-Based Complement. Altern. Med. 2013, 2013, 617459. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.J.G.; Frederico, M.J.S.; Cazarolli, L.H.; Mendes, C.P.; Bretanha, L.C.; Schmidt, E.C.; Bouzon, Z.L.; Pinto, V.A.D.M.; Ramos, C.D.F.; Pizzolatti, M.G.; et al. The mechanism of action of ursolic acid as insulin secretagogue and insulinomimetic is mediated by cross-talk between calcium and kinases to regulate glucose balance. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2015, 1850, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Birgani, G.A.; Ahangarpour, A.; Kahorsandi, L.; Moghaddam, H.F. Anti-diabetic effect of betulinic acid on streptozotocin-nicotinamide induced diabetic male mouse model. Braz. J. Pharm. Sci. 2018, 54. [Google Scholar] [CrossRef] [Green Version]
- Sayem, A.S.M.; Arya, A.; Karimian, H.; Krishnasamy, N.; Hasamnis, A.A.; Hossain, C.F. Action of Phytochemicals on Insulin Signaling Pathways Accelerating Glucose Transporter (GLUT4) Protein Translocation. Molecules 2018, 23, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung, T.N.; Qu, Z.; Kortschak, R.D.; Adelson, D.L. Understanding the Effectiveness of Natural Compound Mixtures in Cancer through Their Molecular Mode of Action. Int. J. Mol. Sci. 2017, 18, 656. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available. |




{kind=link}
{kind=link}
{kind=link}
| Reagent | 1 Reaction (µL) | 50 Reactions (µL) |
|---|---|---|
| Luciferase reagent | 100 | 5000 |
| NADP+ | 1 | 50 |
| G6PDH | 2.5 | 125 |
| Reductase | 0.5 | 25 |
| Reductase substrate | 0.0625 | 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Etsassala, N.G.E.R.; Ndjoubi, K.O.; Mbira, T.J.; Pearce, B.; Pearce, K.; Iwuoha, E.I.; Hussein, A.A.; Benjeddou, M. Glucose-Uptake Activity and Cytotoxicity of Diterpenes and Triterpenes Isolated from Lamiaceae Plant Species. Molecules 2020, 25, 4129. https://doi.org/10.3390/molecules25184129
Etsassala NGER, Ndjoubi KO, Mbira TJ, Pearce B, Pearce K, Iwuoha EI, Hussein AA, Benjeddou M. Glucose-Uptake Activity and Cytotoxicity of Diterpenes and Triterpenes Isolated from Lamiaceae Plant Species. Molecules. 2020; 25(18):4129. https://doi.org/10.3390/molecules25184129
Chicago/Turabian StyleEtsassala, Ninon G. E. R., Kadidiatou O. Ndjoubi, Thilly J. Mbira, Brendon Pearce, Keenau Pearce, Emmanuel I. Iwuoha, Ahmed A. Hussein, and Mongi Benjeddou. 2020. "Glucose-Uptake Activity and Cytotoxicity of Diterpenes and Triterpenes Isolated from Lamiaceae Plant Species" Molecules 25, no. 18: 4129. https://doi.org/10.3390/molecules25184129
APA StyleEtsassala, N. G. E. R., Ndjoubi, K. O., Mbira, T. J., Pearce, B., Pearce, K., Iwuoha, E. I., Hussein, A. A., & Benjeddou, M. (2020). Glucose-Uptake Activity and Cytotoxicity of Diterpenes and Triterpenes Isolated from Lamiaceae Plant Species. Molecules, 25(18), 4129. https://doi.org/10.3390/molecules25184129