
Bioactive Compounds and Total Sugar Contents of Different Open-Pollinated Beetroot Genotypes Grown Organically


Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Dry Matter Content
2.2. Total Soluble Sugar Content
2.3. Nitrate Content
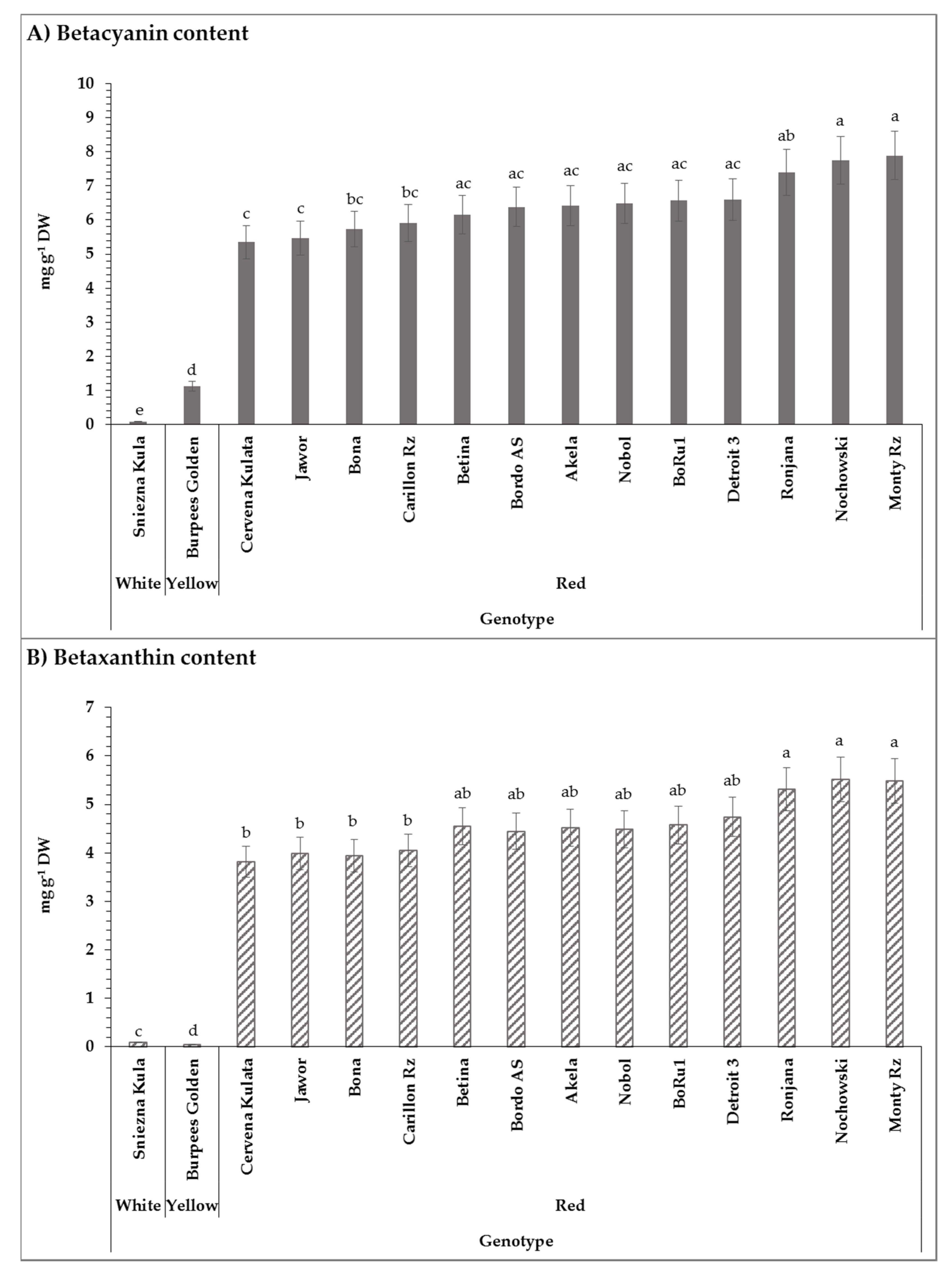
2.4. Betalain Content
2.5. Total Phenolic Content
3. Methods and Materials
3.1. Plant Materials and Sample Preparation
3.2. Chemicals and Reagents
3.3. Total Dry Matter Content
3.4. Total Soluble Sugar Content
3.5. Quantitation of Nitrate Content
3.6. Betalain Content
3.6.1. Extraction of Betalains
3.6.2. Betalains Content Determination
3.7. Total Phenolic Content
3.7.1. Extraction of Total Phenolic Compounds
3.7.2. Total Phenolic Content Quantification
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, R.H. Health-Promoting Components of Fruits and Vegetables in the Diet. Adv. Nutr. 2013, 4, 384S–392S. [Google Scholar] [CrossRef] [PubMed]
- Wruss, J.; Waldenberger, G.; Huemer, S.; Uygun, P.; Lanzerstorfer, P.; Müller, U.; Höglinger, O.; Weghuber, J. Compositional characteristics of commercial beetroot products and beetroot juice prepared from seven beetroot varieties grown in Upper Austria. J. Food Compos. Anal. 2015, 42, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive compounds of beetroot and utilization in food processing industry: A critical review. Food Chem. 2019, 272, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E. The Potential Benefits of Red Beetroot Supplementation in Health and Disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef]
- Vasconcellos, J.; Conte-Junior, C.A.; Silva, D.; Pierucci, A.P.T.R.; Paschoalin, V.; Alvares, T.S. Comparison of total antioxidant potential, and total phenolic, nitrate, sugar, and organic acid contents in beetroot juice, chips, powder, and cooked beetroot. Food Sci. Biotechnol. 2016, 25, 79–84. [Google Scholar] [CrossRef]
- Harsh Chawla Milind Parle, K.S. Beetroot: A Health Promoting Functional Food. Inven. Rapid Nutraceuticals 2016, 1, 1–5. [Google Scholar]
- Hepaksoy, S.; Eroğul, D.; Sen, F.; Aksoy, U. Antioxidant Activity and Total Phenolic Content of Some Turkish Pomegranate Varieties. Acta Hortic. 2009, 37, 241–248. [Google Scholar] [CrossRef]
- Ülger, T.G.; Songur, A.N.; Çırak, O.; Çakıroğlu, F.P. Role of Vegetables in Human Nutrition and Disease Prevention. In Vegetables—Importance of Quality Vegetables to Human Health; IntechOpen: London, UK, 2018; pp. 7–32. [Google Scholar]
- Roshan, B.; Ravindranath, S. Free Radical Pharmacology and Its Role in Various Diseases. J. Drug Deliv. Ther. 2019, 9, 661–668. [Google Scholar]
- Kujala, T.; Vienola, M.; Klika, K.; Loponen, J.; Pihlaja, K. Betalain and phenolic compositions of four beetroot (Beta vulgaris) cultivars. Eur. Food Res. Technol. 2002, 214, 505–510. [Google Scholar] [CrossRef]
- Bavec, M.; Turinek, M.; Grobelnik-Mlakar, S.; Slatnar, A.; Bavec, F. Influence of Industrial and Alternative Farming Systems on Contents of Sugars, Organic Acids, Total Phenolic Content, and the Antioxidant Activity of Red Beet (Beta vulgaris L. ssp. Vulgaris Rote Kugel). J. Agric. Food Chem. 2010, 58, 11825–11831. [Google Scholar] [CrossRef]
- Worthington, V. Nutritional Quality of Organic Versus Conventional Fruits, Vegetables, and Grains. J. Altern. Complement. Med. 2001, 7, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Bourn, D.; Prescott, J. A Comparison of the Nutritional Value, Sensory Qualities, and Food Safety of Organically and Conventionally Produced Foods. Crit. Rev. Food Sci. Nutr. 2002, 42, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Rembiałkowska, E. Quality of plant products from organic agriculture. J. Sci. Food Agric. 2007, 87, 2757–2762. [Google Scholar] [CrossRef]
- Velimirov, A.; Huber, M.; Lauridsen, C.; Rembiałkowska, E.; Seidel, K.; Bügel, S. Feeding trials in organic food quality and health research. J. Sci. Food Agric. 2009, 90, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Rembialkowska, E. Wholesomeness and Sensory Quality of the Potatoes and Selected Vegetables from the Organic Farms. Ph.D. Thesis, Warsaw Agricultural University, Warsaw, Poland, 2000. [Google Scholar]
- Goksoy, A.; Turkec, A.; Turan, Z. Determination of some agronomic characteristics and hybrid vigor of new improved synthetic varieties in sunflower (helianthus annuus L.). Helia 2002, 25, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Torricelli, R.; Ciancaleoni, S.; Negri, V. Performance and stability of homogeneous and heterogeneous broccoli (Brassica oleracea L. var. italica Plenck) varieties in organic and low-input conditions. Euphytica 2014, 199, 385–395. [Google Scholar] [CrossRef]
- Nizioł-Łukaszewska, Z.; Gawęda, M. Selected Indicators of the Root Quality of Fifteen Cultivars of Red Beet (Beta Vulgaris L.). J. Hortic. Res. 2015, 23, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Kosson, R.; Elkner, K.; Szafirowska, A. Quality of Fresh and Processed Red Beet from Organic and Conventional Cultivation. Veg. Crop. Res. Bull. 2011, 75, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Brandt, K.; Mølgaard, J.P. Organic agriculture: Does it enhance or reduce the nutritional value of plant foods? J. Sci. Food Agric. 2001, 81, 924–931. [Google Scholar] [CrossRef]
- Hallmann, E.; Rembiałkowska, E. Characterisation of antioxidant compounds in sweet bell pepper (Capsicum annuum L.) under organic and conventional growing systems. J. Sci. Food Agric. 2012, 92, 2409–2415. [Google Scholar] [CrossRef]
- Sikora, M.; Hallmann, E.; Rembiałkowska, E. Comparison of the nutritional value of red beet roots from organic and conventional production. In Proceedings of the Bioacademy 2008—Proceedings, New Developments in Science and Research on Organic Agriculture; Lenice na Moravě, Czech Republic, 3–5 September 2008; pp. 154–156. [Google Scholar]
- Hecke, K.; Herbinger, K.; Veberič, R.; Trobec, M.; Toplak, H.; Štampar, F.; Keppel, H.; Grill, D. Sugar, acid and phenol contents in apple cultivars from organic and integrated fruit cultivation. Eur. J. Clin. Nutr. 2006, 60, 1136–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szopińska, A.A.; Gawęda, M. Comparison of Yield and Quality of Red Beet Roots Cultivated Using Conventional, Integrated and Organic Method. J. Hortic. Res. 2013, 21, 107–114. [Google Scholar] [CrossRef]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2005, 86, 10–17. [Google Scholar] [CrossRef]
- Andac, M.; Eren, H.; Coldur, F. Determination of Nitrate in Leafy Vegetables by Flow Injection Analysis with Potentiometric Detection. J. Food Drug Anal. 2011, 19, 457–462. [Google Scholar]
- Hirata, S.; Amma, B.V.; Karthikeyan, S.; Toda, K. Determination of Nitrite by Flow Injection Spectrophotometry Using a Home-Made Flow Cell Detector. Anal. Sci. 2003, 19, 1687–1689. [Google Scholar] [CrossRef] [Green Version]
- Bedale, W.; Sindelar, J.J.; Milkowski, A.L. Dietary Nitrate and Nitrite: Benefits, Risks, and Evolving Perceptions. Meat Sci. 2016, 120, 85–92. [Google Scholar] [CrossRef]
- Cermak, N.M.; Gibala, M.J.; Van Loon, L.J.C. Nitrate Supplementation’s Improvement of 10-Km Time-Trial Performance in Trained Cyclists. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 64–71. [Google Scholar] [CrossRef]
- Bailey, S.J.; Winyard, P.; Vanhatalo, A.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Tarr, J.; Benjamin, N.; Jones, A.M. Dietary Nitrate Supplementation Reduces the O2 Cost of Low-Intensity Exercise and Enhances Tolerance to High-Intensity Exercise in Humans. J. Appl. Physiol. 2009, 107, 1144–1155. [Google Scholar] [CrossRef] [Green Version]
- Hernández, A.; Schiffer, T.A.; Ivarsson, N.; Cheng, A.J.; Bruton, J.D.; Lundberg, J.O.; Weitzberg, E.; Westerblad, H. Dietary Nitrate Increases Tetanic [Ca2+] i and Contractile Force in Mouse Fast-Twitch Muscle. J. Physiol. 2012, 590, 3575–3583. [Google Scholar] [CrossRef]
- Grzebelus, D.; Baranski, R. Identification of Accessions Showing Low Nitrate Accumulation in a Germplasm Collection of Garden Beet. Acta Hortic. 2001, 563, 253–257. [Google Scholar] [CrossRef]
- Feller, C.; Fink, M. Nitrate Content, Soluble Solids Content, and Yield of Table Beet as Affected by Cultivar, Sowing Date and Nitrogen Supply. HortScience 2004, 39, 1255–1259. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.W.C.; Tran, J.C.H.; Tong, K.S.K.; Chen, M.Y.Y.; Xiao, Y.; Ho, Y.Y.; Chan, C.H.Y. Nitrate and Nitrite Levels in Commonly Consumed Vegetables in Hong Kong. Food Addit. Contam. Part. B Surveill. 2011, 4, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Siomos, A.S.; Dogras, C.C. Nitrates in Vegetables Produced in Greece. J. Veg. Crop. Prod. 2008, 5, 3–13. [Google Scholar] [CrossRef]
- Rubóczki, T.; Raczkó, V.; Takácsné Hájos, M. Evaluation of Morphological Parameters and Bioactive Compounds in Different Varieties of Beetroot (Beta Vulgaris L. Ssp. Esculenta GURKE Var. Rubra L.). Int. J. Hortic. Sci. 2015, 21, 31–35. [Google Scholar] [CrossRef]
- Santamaria, P.; Gonnella, M.; Elia, A.; Parente, A.; Serio, F. Ways of Reducing Rocket Salad Nitrate Content. In International Symposium on Growing Media and Hydroponics; Maloupa, E., Gerasopoulos, D., Eds.; ISHS Acta Horticulturae 548: Kassandra, Macedonia, 2001; pp. 529–537. [Google Scholar]
- Felczynski, K.; Elkner, K. Effect of Long-Term Organic and Mineral Fertilization on the Yield and Quality of Red Beet (Beta Vulgaris L.). Veg. Crop. Res. Bull. 2008, 68, 111–125. [Google Scholar] [CrossRef]
- Lidder, S.; Webb, A.J. Vascular Effects of Dietary Nitrate (as Found in Green Leafy Vegetables and Beetroot) via the Nitrate-Nitrite-Nitric Oxide Pathway. Br. J. Clin. Pharmacol. 2013, 75, 677–696. [Google Scholar] [CrossRef] [Green Version]
- Harada, H.; Yoshimura, Y.; Sunaga, Y.; Hatanaka, T.; Sugita, S. Breeding of Italian Ryegrass (Lolium Multiflorum Lam.) for a Low Nitrate Concentration by Seedling Test. Euphytica 2003, 129, 201–209. [Google Scholar] [CrossRef]
- Hord, N.G.; Tang, Y.; Bryan, N.S. Food Sources of Nitrates and Nitrites: The Physiologic Context for Potential Health Benefits. Am. J. Clin. Nutr. 2009, 90, 1–10. [Google Scholar] [CrossRef]
- Wang, X.; Cai, X.; Xu, C.; Wang, S.; Dai, S.; Wang, Q. Nitrate Accumulation and Expression Patterns of Genes Involved in Nitrate Transport and Assimilation in Spinach. Molecules 2018, 23, 2231. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, H.M.C. Betalains: Properties, Sources, Applications, and Stability-A Review. Int. J. Food Sci. Technol. 2009, 44, 2365–2376. [Google Scholar] [CrossRef] [Green Version]
- Slatnar, A.; Stampar, F.; Veberic, R.; Jakopic, J. HPLC-MSn Identification of Betalain Profile of Different Beetroot (Beta Vulgaris L. Ssp. Vulgaris) Parts and Cultivars. J. Food Sci. 2015, 80, C1952–C1958. [Google Scholar] [CrossRef]
- Strack, D.; Vogt, T.; Schliemann, W. Recent Advances in Betalain Research. Phytochemistry 2003, 62, 247–269. [Google Scholar] [CrossRef]
- Gengatharan, A.; Dykes, G.A.; Choo, W.S. Betalains: Natural Plant Pigments with Potential Application in Functional Foods. Lwt 2015, 64, 645–649. [Google Scholar] [CrossRef]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. Purification and Antiradical Properties of the Structural Unit of Betalains. J. Nat. Prod. 2012, 75, 1030–1036. [Google Scholar] [CrossRef]
- Sawicki, T.; Bączek, N.; Wiczkowski, W. Betalain Profile, Content and Antioxidant Capacity of Red Beetroot Dependent on the Genotype and Root Part. J. Funct. Foods 2016, 27, 249–261. [Google Scholar] [CrossRef]
- Kujala, T.S.; Loponen, J.M.; Klika, K.D.; Pihlaja, K. Phenolics and Betacyanins in Red Beetroot (Beta Vulgaris) Root: Distribution and Effect of Cold Storage on the Content of Total Phenolics and Three Individual Compounds. J. Agric. Food Chem. 2000, 48, 5338–5342. [Google Scholar] [CrossRef] [PubMed]
- Gasztonyi, M.N.; Daood, H.; Hájos, M.T.; Biacs, P. Comparison of Red Beet (Beta Vulgaris Var Conditiva) Varieties on the Basis of Their Pigment Components. J. Sci. Food Agric. 2001, 81, 932–933. [Google Scholar] [CrossRef]
- Lee, E.J.; An, D.; Nguyen, C.T.T.; Patil, B.S.; Kim, J.; Yoo, K.S. Betalain and Betaine Composition of Greenhouse- or Field-Produced Beetroot (Beta Vulgaris L.) and Inhibition of HepG2 Cell Proliferation. J. Agric. Food Chem. 2014, 62, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tsao, R. Dietary Polyphenols, Oxidative Stress and Antioxidant and Anti-Inflammatory Effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Ribas-Agustí, A.; Martín-Belloso, O.; Soliva-Fortuny, R.; Elez-Martínez, P. Food Processing Strategies to Enhance Phenolic Compounds Bioaccessibility and Bioavailability in Plant-Based Foods. Crit. Rev. Food Sci. Nutr. 2018, 58, 2531–2548. [Google Scholar] [CrossRef] [Green Version]
- Lattanzio, V.; Lattanzio, V.M.; Cardinali, A. Role of Phenolics in the Resistance Mechanisms of Plants against Fungal Pathogens and Insects. Phytochemistry 2006, 661, 23–67. [Google Scholar]
- Delgado-Vargas, F.; Jiménez, A.R.; Paredes-López, O. Natural Pigments: Carotenoids, Anthocyanins, and Betalains-Characteristics, Biosynthesis, Processing, and Stability Natural Pigments: Carotenoids, Anthocyanins, and Betalains-Characteristics, Biosynthesis, Processing, and Stability. Crit. Rev. Food Sci. Nutr. 2000, 40, 173–289. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Hathan, B.S.; Devi, C.; Memorial, L.; Longowal, S. Chemical Composition, Functional Properties and Processing of Beetroot-A Review. Int. J. Sci. Eng. Res. 2014, 5, 679–684. [Google Scholar]
- Ninfali, P.; Angelino, D. Nutritional and Functional Potential of Beta Vulgaris Cicla and Rubra. Fitoterapia 2013, 89, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Kovarovič, J.; Bystrická, J.; Tomáš, J.; Lenková, M. The Influence of Variety on the Content of Bioactive Compounds in Beetroot (Beta Vulgaris L.). Potravin. Slovak J. Food Sci. 2017, 11, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Kavalcová, P.; Bystrická, J.; Tomáš, J.; Karovičová, J.; Kovarovič, J.; Lenková, M. The Content of Total Polyphenols and Antioxidant Activity in Red Beetroot. Potravinarstvo 2015, 9, 77–83. [Google Scholar] [CrossRef]
- Asami, D.K.; Hong, Y.J.; Barrett, D.M.; Mitchell, A.E. Comparison of the Total Phenolic and Ascorbic Acid Content of Freeze-Dried and Air-Dried Marionberry, Strawberry, and Corn Grown Using Conventional, Organic, and Sustainable Agricultural Practices. J. Agric. Food Chem. 2003, 51, 1237–1241. [Google Scholar] [CrossRef]
- Carrillo, C.; Wilches-Pérez, D.; Hallmann, E.; Kazimierczak, R.; Rembiałkowska, E. Organic versus Conventional Beetroot. Bioactive Compounds and Antioxidant Properties. Lwt 2019, 116, 108552. [Google Scholar] [CrossRef]
- Straus, S.; Bavec, F.; Turinek, M.; Slatnar, A.; Rozman, C.; Bavec, M. Nutritional Value and Economic Feasibility of Red Beetroot (Beta Vulgaris L. Ssp. Vulgaris Rote Kugel) from Different Production Systems. African J. Agric. Res. 2012, 7, 5653–5660. [Google Scholar] [CrossRef]
- Vulić, J.; Čanadanović-Brunet, J.; Ćetković, G.; Tumbas, V.; Djilas, S.; Četojević-Simin, D.; Čanadanović, V. Antioxidant and Cell Growth Activities of Beet Root Pomace Extracts. J. Funct. Foods 2012, 4, 670–678. [Google Scholar] [CrossRef]
- Stratil, P.; Klejdus, B.; Kubáň, V. Determination of Total Content of Phenolic Compounds and Their Antioxidant Activity in Vegetables-Evaluation of Spectrophotometric Methods. J. Agric. Food Chem. 2006, 54, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Biesiada, A.; Tomczak, A. Biotic and Abiotic Factors Affecting the Content of the Chosen Antioxidant Compounds in Vegetables. Veg. Crop. Res. Bull. 2012, 76, 55–78. [Google Scholar] [CrossRef]
- Gajic, I.S.; Savic, I.; Boskov, I.; Žerajić, S.; Markovic, I.; Gajic, D. Optimization of Ultrasound-Assisted Extraction of Phenolic Compounds from Black Locust (Robiniae Pseudoacaciae) Flowers and Comparison with Conventional Methods. Antioxidants 2019, 8, 248. [Google Scholar] [CrossRef] [Green Version]
- Savic, I.M.; Savic Gajic, I.M. Optimization of Ultrasound-Assisted Extraction of Polyphenols from Wheatgrass (Triticum Aestivum L.). J. Food Sci. Technol. 2020, 57, 2809–2818. [Google Scholar] [CrossRef] [PubMed]
- Yasaminshirazi, K.; Hartung, J.; Groenen, R.; Heinze, T.; Fleck, M.; Zikeli, S.; Graeff-Hoenninger, S. Agronomic Performance of Different Open-Pollinated Beetroot Genotypes Grown Under Organic Farming Conditions. Agronomy 2020, 10, 812. [Google Scholar] [CrossRef]
- Koubaier, H.B.H.; Snoussi, A.; Essaidi, I.; Chaabouni, M.M.; Thonart, P.; Bouzouita, N. Betalain and Phenolic Compositions, Antioxidant Activity of Tunisian Red Beet (Beta Vulgaris L. Conditiva) Roots and Stems Extracts. Int. J. Food Prop. 2014, 17, 1934–1945. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzym. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Wolfinger, R. Covariance Structure Selection in General Mixed Models. Commun. Stat.-Simul. Comput. 1993, 22, 1079–1106. [Google Scholar] [CrossRef]
- Piepho, H.P. An Algorithm for a Letter-Based Representation of All-Pairwise Comparisons. J. Comput. Graph. Stat. 2004, 13, 456–466. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genotype | Nitrate (mg kg−1 DW) | Total Soluble Sugar (°Bx) | Total Phenolic Content (mg GAE 100 g−1 DW) | Total Dry Matter Content (%) |
|---|---|---|---|---|
| Akela RZ | 8865 a,c ± 1209 | 11.94 c,e ± 0.37 | 402.55 b,c ± 28.24 | 14.95 d,e,f ± 4.56 |
| Betina | 5817 b,c,d ± 1210 | 13.88 a,b ± 0.37 | 393.94 b,c ± 28.24 | 17.12 a,b ± 4.56 |
| Bona | 9364 a,b ± 1221 | 12.39 c,d ± 0.37 | 395.55 b,c ± 28.24 | 15.44 c,e,f ± 4.56 |
| Bordo | 7023 b,c,d ± 1210 | 12.80 b,c ± 0.37 | 420.29 a,c ± 28.24 | 16.21 a,b,c,d ± 4.56 |
| BoRu1 | 4597 d ± 1211 | 12.47 c ± 0.37 | 382.73 c ± 28.24 | 15.47 c,e,f ± 4.56 |
| Burpees Golden | 7566 a,d ± 1211 | 11.94 c,e ± 0.37 | 172.89 d ± 28.78 | 15.98 b,e ± 4.56 |
| Carillon RZ | 10924 a ± 1211 | 11.19 e ± 0.37 | 352.46 c ± 28.24 | 14.12 f ± 4.56 |
| Cervena Kulata | 5768 b,c,d ± 1228 | 13.94 a,b ± 0.37 | 476.84 a,b ± 28.24 | 16.41 a,b,c,d ± 4.56 |
| Detroit 3 | 7682 a,d ± 1211 | 11.99 ce ± 0.37 | 402.61 b,c ± 28.24 | 15.06 c,e,f ± 4.56 |
| Jawor | 7855 a,d ± 1211 | 12.60 c ± 0.37 | 415.82 a,c ± 28.24 | 15.55 c,e ± 4.56 |
| Monty RZ F1 | 5487 c,d ± 1209 | 12.33 c,e ± 0.37 | 489.06 a ± 28.24 | 15.41 c,e,f ± 4.56 |
| Nobol | 9447 a,b ± 1210 | 11.28 d,e ± 0.37 | 401.23 b,c ± 28.24 | 14.66 e,f ± 4.60 |
| Nochowski | 4593 d ± 1212 | 14.67 a ± 0.37 | 432.90 a,c ± 28.24 | 17.50 a ± 4.56 |
| Ronjana | 8595 a.c ± 1210 | 12.80 b,c ± 0.37 | 405.32 a,c ± 28.24 | 15.34 c,e,f ± 4.56 |
| Sniezna Kula | 7427 a,d ± 1211 | 11.91 c,e ± 0.37 | 216.09 d ± 29.87 | 16.05 b,e ± 4.56 |
| Factor | p-Value of the F-Test of the Corresponding Factor | |||
| Location | <0.0001 | 0.0008 | n.s. 1 | 0.0013 |
| Year | n.s. | <0.0001 | <0.0001 | <0.0001 |
| Genotype | 0.0191 | 0.0028 | 0.0002 | 0.0029 |
| Location × year | 0.0495 | <0.0001 | 0.0114 | <0.0001 |
Sample Availability: Not available. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasaminshirazi, K.; Hartung, J.; Fleck, M.; Graeff-Hoenninger, S. Bioactive Compounds and Total Sugar Contents of Different Open-Pollinated Beetroot Genotypes Grown Organically. Molecules 2020, 25, 4884. https://doi.org/10.3390/molecules25214884
Yasaminshirazi K, Hartung J, Fleck M, Graeff-Hoenninger S. Bioactive Compounds and Total Sugar Contents of Different Open-Pollinated Beetroot Genotypes Grown Organically. Molecules. 2020; 25(21):4884. https://doi.org/10.3390/molecules25214884
Chicago/Turabian StyleYasaminshirazi, Khadijeh, Jens Hartung, Michael Fleck, and Simone Graeff-Hoenninger. 2020. "Bioactive Compounds and Total Sugar Contents of Different Open-Pollinated Beetroot Genotypes Grown Organically" Molecules 25, no. 21: 4884. https://doi.org/10.3390/molecules25214884
APA StyleYasaminshirazi, K., Hartung, J., Fleck, M., & Graeff-Hoenninger, S. (2020). Bioactive Compounds and Total Sugar Contents of Different Open-Pollinated Beetroot Genotypes Grown Organically. Molecules, 25(21), 4884. https://doi.org/10.3390/molecules25214884