
Dose-Dependent Effects of Dietary Xylooligosaccharides Supplementation on Microbiota, Fermentation and Metabolism in Healthy Adult Cats

 , ,
, , _Ran.png) ,
, 
Abstract
:1. Introduction
2. Results
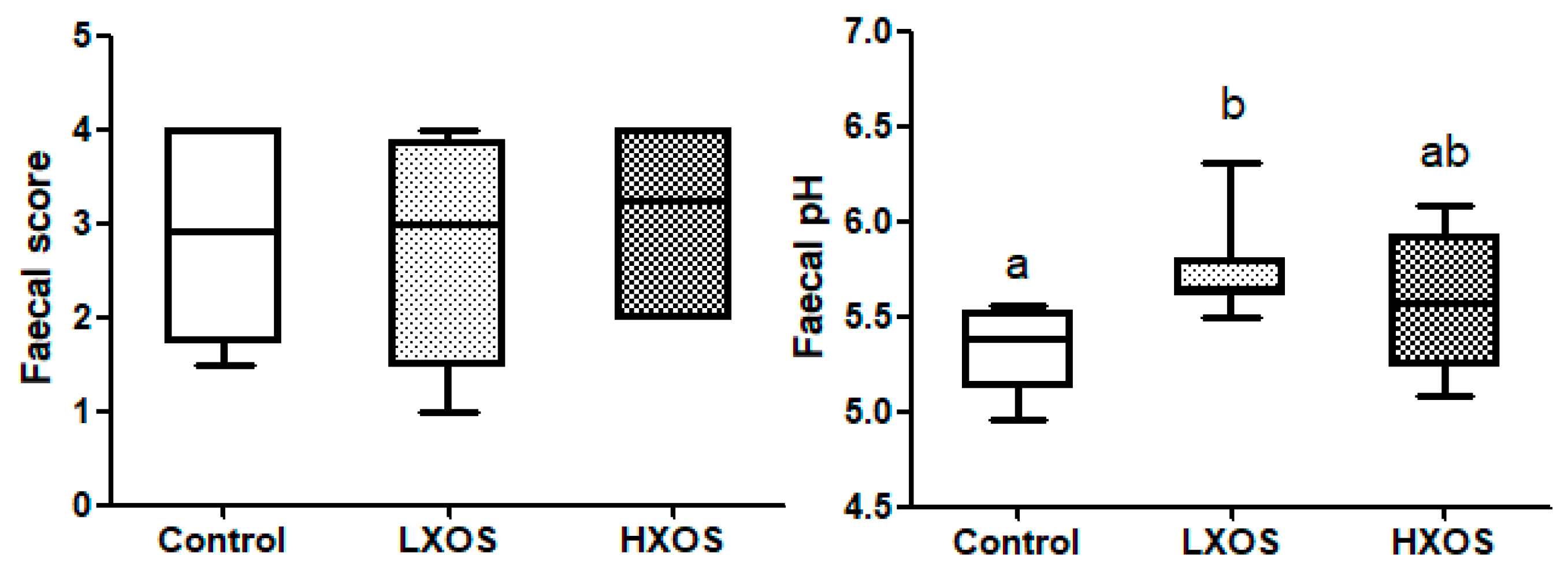
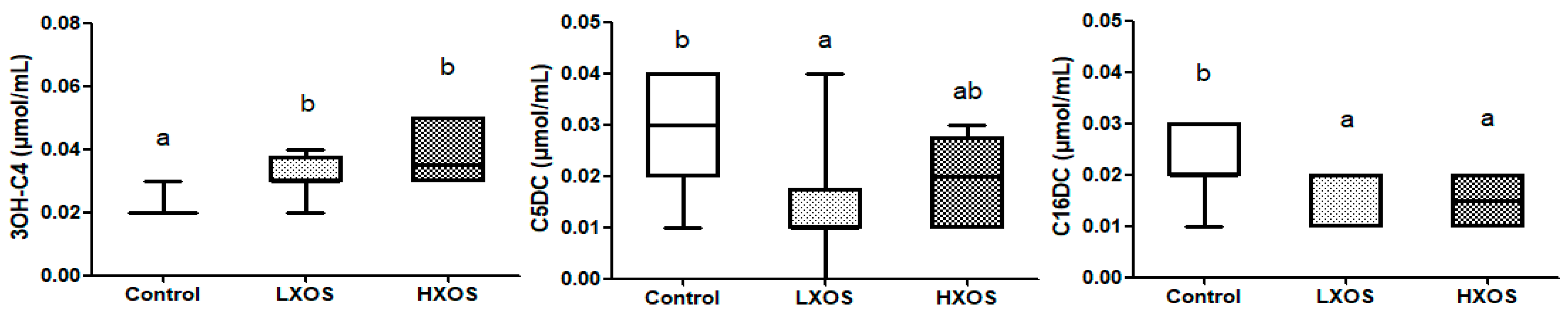
2.1. Fecal Parameters
2.2. Blood Parameters
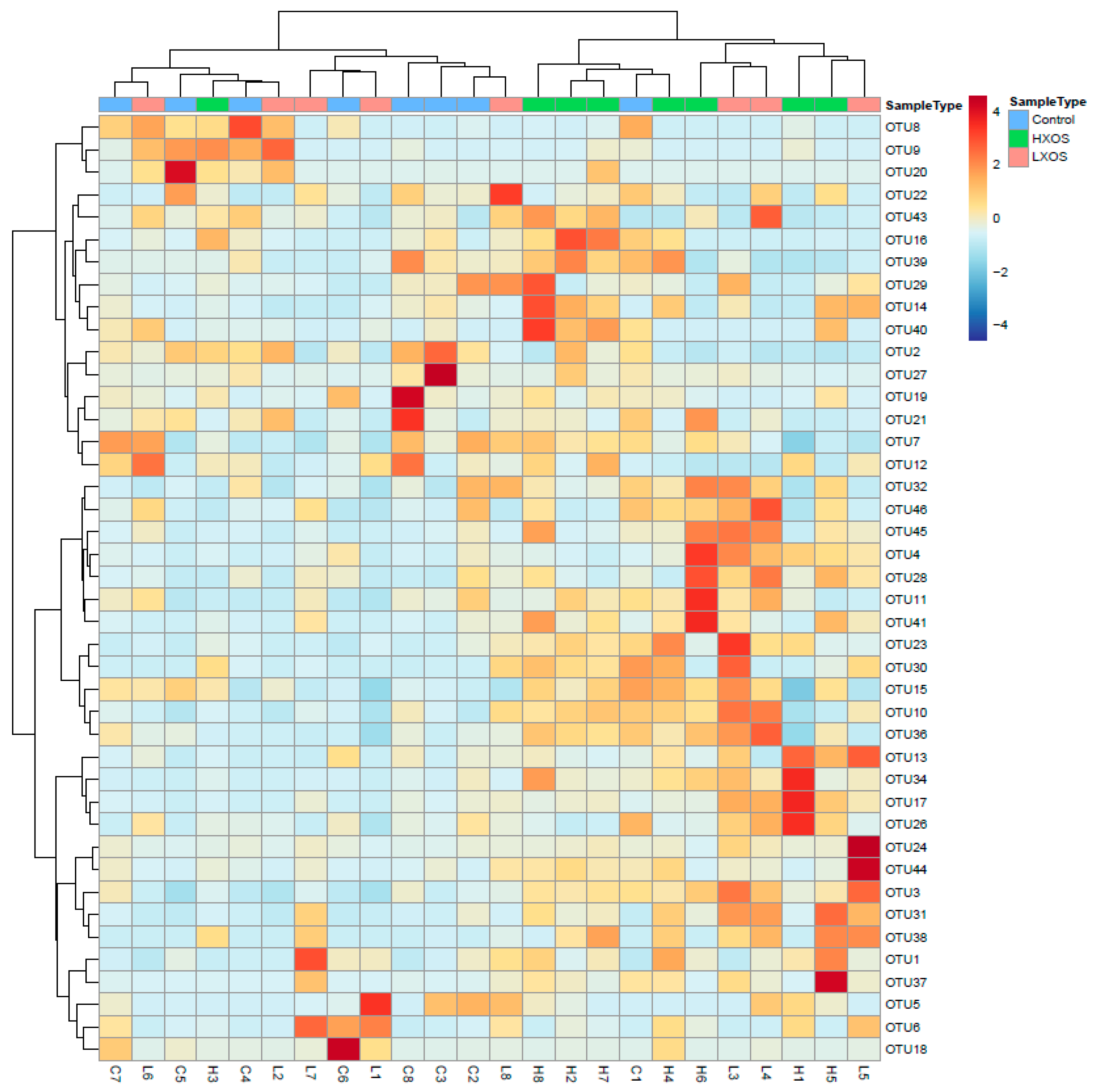
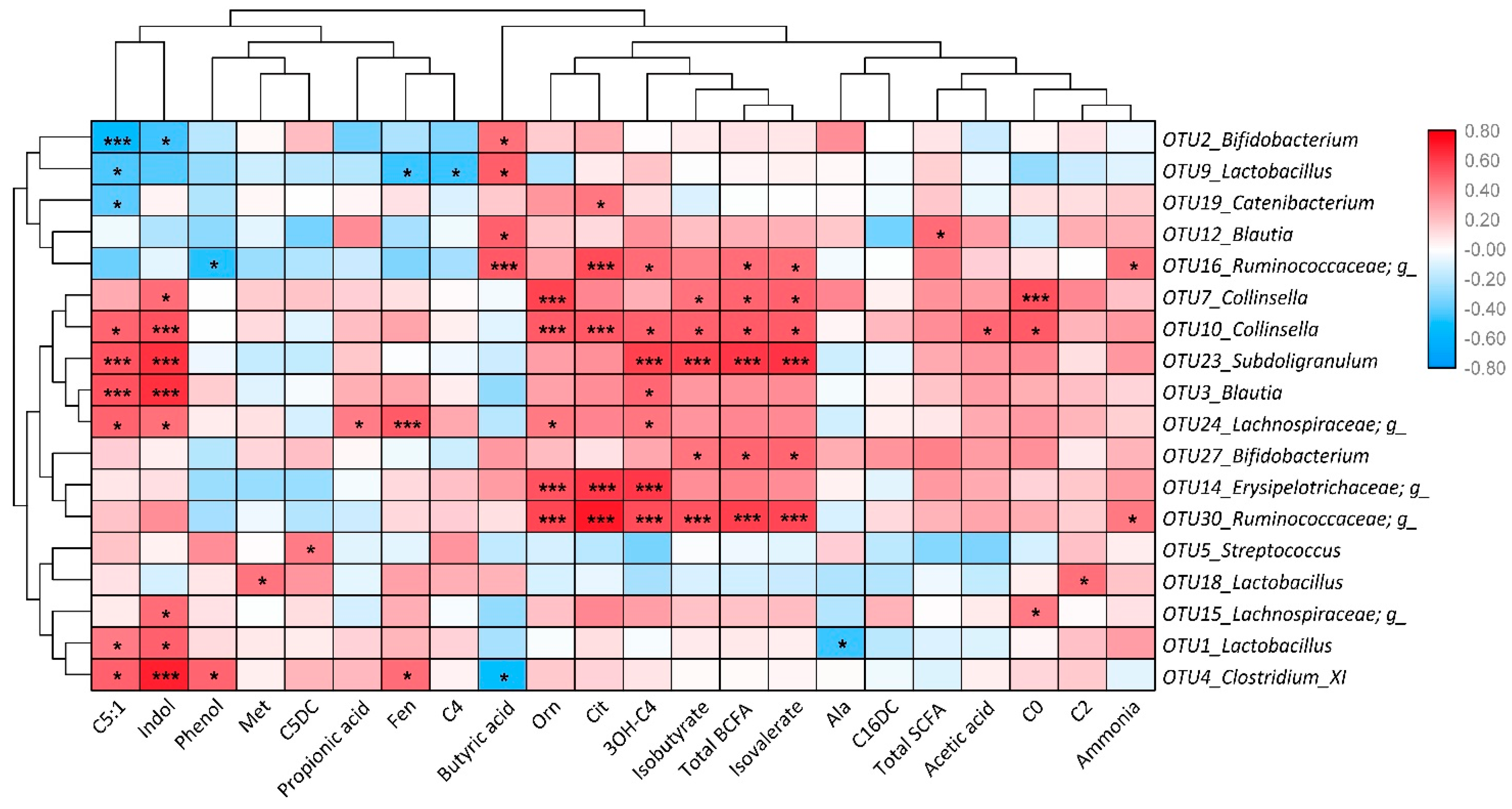
2.3. Microbiome Profile
3. Discussion
3.1. Both XOS Treatments Influenced Intestinal Metabolism and Microbiome
3.2. High XOS Supplementation could Promote Intestinal Fermentation
3.3. Low XOS Supplementation could Enhance Carbohydrate Metabolism
3.4. Could a Low Dose be Better?
4. Materials and Methods
4.1. Animals and Diets
4.2. Sample Collection
4.3. Analytical Methods
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Esgalhado, M.; Kemp, J.A.; Damasceno, N.R.; Fouque, D.; Mafra, D. Short-chain fatty acids: A link between prebiotics and microbiota in chronic kidney disease. Future Microbiol. 2017, 12, 1413–1425. [Google Scholar] [CrossRef] [PubMed]
- Dewulf, E.M.; Cani, P.D.; Claus, S.P.; Fuentes, S.; Puylaert, P.G.; Neyrinck, A.M.; Bindels, L.B.; de Vos, W.M.; Gibson, G.R.; Thissen, J.P.; et al. Insight into the prebiotic concept: Lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 2013, 62, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Lecourt, E.; Dewulf, E.M.; Sohet, F.M.; Pachikian, B.D.; Naslain, D.; De Backer, F.; Neyrinck, A.M.; Delzenne, N.M. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am. J. Clin. Nutr. 2009, 90, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Ghouri, Y.; Richards, M.; Rahimi, E.; Krill, J.; Jelinek, K.A.; Dupont, A.W. Systematic review of randomized controlled trials of probiotics, prebiotics, and synbiotics in inflammatory bowel disease. Clin. Exp. Gastroenterol. 2014, 7, 473–487. [Google Scholar]
- Respondek, F.; Swanson, K.S.; Belsito, K.R.; Vester, B.M.; Wagner, A.; Istasse, L.; Diez, M. Short-chain fructooligosaccharides influence insulin sensitivity and gene expression of fat tissue in obese dogs. J. Nutr. 2008, 138, 1712–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Respondek, F.; Myers, K.; Smith, T.L.; Wagner, A.; Geor, R.J. Dietary supplementation with short-chain fructo-oligosaccharides improves insulin sensitivity in obese horses. J. Anim. Sci. 2011, 89, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Che, T.M.; Johnson, R.W.; Kelley, K.W.; Van Alstine, W.G.; Dawson, K.A.; Moran, C.A.; Pettigrew, J.E. Mannan oligosaccharide modulates gene expression profile in pigs experimentally infected with porcine reproductive and respiratory syndrome virus. J. Anim. Sci. 2011, 89, 3016–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.H.; Kim, I.H. Effects of lactulose supplementation on performance, blood profiles, excreta microbial shedding of Lactobacillus and Escherichia coli, relative organ weight and excreta noxious gas contents in broilers. J. Anim. Physiol. Anim. Nutr. 2014, 98, 424–430. [Google Scholar] [CrossRef]
- Vázquez, M.J.; Alonso, J.L.; Domínguez, H.; Parajó, J.C. Xylooligosaccharides: Manufacture and applications. Trends Food Sci. Tech. 2000, 11, 387. [Google Scholar] [CrossRef]
- Jain, I.; Kumar, V.; Satyanarayana, T. Xylooligosaccharides: An economical prebiotic from agroresidues and their health benefits. Indian J. Exp. Biol. 2015, 53, 131–142. [Google Scholar]
- Aachary, A.A.; Prapulla, S.G. Xylooligosaccharides (xos) as an emerging prebiotic: Microbial synthesis, utilization, structural characterization, bioactive properties, and applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 2–16. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Courtin, C.M.; Verbeke, K.; Van de Wiele, T.; Verstraete, W.; Delcour, J.A. Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 2011, 51, 178–194. [Google Scholar] [CrossRef] [PubMed]
- Gobinath, D.; Madhu, A.N.; Prashant, G.; Srinivasan, K.; Prapulla, S.G. Beneficial effect of xylo-oligosaccharides and fructo-oligosaccharides in streptozotocin-induced diabetic rats. Brit. J. Nutr. 2010, 104, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaizumi, K.; Nakatsu, Y.; Sato, M.; Sedarnawati, Y.; Sugano, M. Effects of xylooligosaccharides on blood glucose, serum and liver lipids and cecum short-chain fatty acids in diabetic rats. Agri. Biol. Chem. 1991, 55, 199–205. [Google Scholar]
- Sheu, H.H.; Lee, I.T.; Chen, W.; Chan, Y.C. Effects of xylooligosaccharides in type 2 diabetes mellitus. J. Nutr. Sci. Vitaminol. 2008, 54, 396–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, M.; Kim, W. Effects of xylooligosaccharide intake on fecal Bifidobacteria, lactic acid and lipid metabolism in Korean young women. Korean J. Nutr. 2007, 40, 154–161. [Google Scholar]
- Bouhnik, Y.; Vahedi, K.; Achour, L.; Attar, A.; Salfati, J.; Pochart, P.; Marteau, P.; Flourié, B.; Bornet, F.; Rambaud, J.C. Short-chain fructo-oligosaccharide administration dose-dependently increases fecal bifidobacteria in healthy humans. J. Nutr. 1999, 129, 113–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M.; Deguchi, Y.; Miyamori, A.; Matsumoto, K.; Kikuchi, H.; Matsumoto, K.; Kobayashi, Y.; Yajima, T.; Kan, T. Effects of Administration of Galactooligosaccharides on the Human Faecal Microflora, Stool Weight and Abdominal Sensation. Microb. Ecol. Health Dis. 1990, 3, 285–292. [Google Scholar]
- Lin, X.; Ning, J.; Xu, G. Application of Xylo-oligosaccharide in modifying human intestinal function. Afr. J. Microbiol. Res. 2012, 6, 2116–2119. [Google Scholar]
- Rochus, K.; Janssens, G.P.; Hesta, M. Dietary fibre and the importance of the gut microbiota in feline nutrition: A review. Nutr. Res. Rev. 2014, 27, 295–307. [Google Scholar] [CrossRef]
- Brosey, B.P.; Hill, R.C.; Scott, K.C. Gastrointestinal volatile fatty acid concentrations and pH in cats. Am. J. Vet. Res. 2000, 61, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Zoran, D.L. The carnivore connection to nutrition in cats. J. Am. Vet. Med. Assoc. 2002, 221, 1559–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbrugghe, A.; Hesta, M.; Daminet, S.; Janssens, G. Nutritional modulation of insulin resistance in the true carnivorous cat: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Hesta, M.; Janssens, G.P.J.; Debraekeleer, J.; Wilde, R.D. The effect of oligofructose and inulin on faecal characteristics and nutrient digestibility in healthy cats. J. Anim. Physiol. Anim. Nutr. 2001, 85, 7. [Google Scholar] [CrossRef] [PubMed]
- Hesta, M.; Hoornaert, E.; Verlinden, A.; Janssens, G.P.J. The effect of oligofructose on urea metabolism and faecal odour components in cats. J. Anim. Physiol. Anim. Nutr. 2005, 89, 7. [Google Scholar] [CrossRef] [PubMed]
- Verbrugghe, A.; Hesta, M.; Gommeren, K.; Daminet, S.; Wuyts, B.; Buyse, J.; Janssens, G.P.J. Oligofructose and inulin modulate glucose and amino acid metabolism through propionate production in normal-weight and obese cats. Brit. J. Nutr. 2009, 102, 694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbrugghe, A.; Hesta, M.; Van Weyenberg, S.; Papadopoulos, G.A.; Gommeren, K.; Daminet, S.; Bosmans, T.; Polis, I.; Buyse, J.; Janssens, G.P.J. The glucose and insulin response to isoenergetic reduction of dietary energy sources in a true carnivore: The domestic cat (felis catus). Br. J. Nutr. 2010, 104, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Kanakupt, K.; Vester Boler, B.M.; Dunsford, B.R.; Fahey, G.C., Jr. Effects of short-chain fructooligosaccharides and galactooligosaccharides, individually and in combination, on nutrient digestibility, fecal fermentative metabolite concentrations, and large bowel microbial ecology of healthy adult cats. J. Anim. Sci. 2011, 89, 1376–1384. [Google Scholar] [CrossRef] [Green Version]
- Barry, K.A.; Wojcicki, B.J.; Middelbos, I.S.; Vester, B.M.; Swanson, K.S.; Fahey, G.C. Dietary cellulose, fructooligosaccharides, and pectin modify fecal protein catabolites and microbial populations in adult cats. J. Anim. Sci. 2010, 88, 2978–2987. [Google Scholar] [CrossRef] [Green Version]
- Barry, K.A.; Middelbos, I.S.; Vester Boler, B.M.; Dowd, S.E.; Suchodolski, J.S.; Henrissat, B.; Coutinho, P.M.; White, B.A.; Fahey, G.C., Jr.; Swanson, K.S. Effects of dietary fiber on the feline gastrointestinal metagenome. J. Proteome Res. 2012, 11, 5924–5933. [Google Scholar] [CrossRef]
- Barry, K.A.; Hernot, D.C.; Van Loo, J.; Fahey, G.C.; de Godoy, M.R.C. Fructan supplementation of senior cats affects stool metabolite concentrations and fecal microbiota concentrations, but not nitrogen partitioning in excreta. J. Anim. Sci. 2014, 92, 4964–4971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.; Muoio, D.M.; Shiota, M.; Fujimoto, Y.; Cline, G.W.; Shulman, G.I.; Koves, T.R.; Stevens, R.; Millington, D.; Newgard, C.B. Hepatic expression of malonyl-CoA decarboxylase reverses muscle, liver and whole-animal insulin resistance. Nat. Med. 2004, 10, 268. [Google Scholar] [CrossRef] [PubMed]
- Ferdinandusse, S.; Denis, S.; Mooijer, P.A.; Zhang, Z.; Reddy, J.K.; Spector, A.A.; Wanders, R.J. Identification of the peroxisomal β-oxidation enzymes involved in the biosynthesis of docosahexaenoic acid. J. Lipid Res. 2001, 42, 1987–1995. [Google Scholar] [PubMed]
- O’Keefe, S.J. Tube feeding, the microbiota, and Clostridium difficile infection. World J. Gastroenterol. 2010, 16, 139. [Google Scholar] [CrossRef]
- Masuoka, H.; Shimada, K.; Kiyosue-Yasuda, T.; Kiyosue, M.; Oishi, Y.; Kimura, S.; Ohashi, Y.; Fujisawa, T.; Hotta, K.; Yamada, A.; et al. Transition of the intestinal microbiota of cats with age. PLoS ONE 2017, 12, e0181739. [Google Scholar] [CrossRef] [Green Version]
- Furet, J.P.; Firmesse, O.; Gourmelon, M.; Bridonneau, C.; Tap, J.; Mondot, S.; Dore, J.; Corthier, G. Comparative assessment of human and farm animal faecal microbiota using real-time quantitative PCR. FEMS Microbiol. Ecol. 2009, 68, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Lahti, L.; Salonen, A.; Kekkonen, R.A.; Salojärvi, J.; Jalanka-Tuovinen, J.; Palva, A.; Orešič, M.; De Vos, W.M. Associations between the human intestinal microbiota, Lactobacillus rhamnosus GG and serum lipids indicated by integrated analysis of high-throughput profiling data. Peer J. 2013, 1, e32. [Google Scholar] [CrossRef] [Green Version]
- Bag, S.; Ghosh, T.S.; Das, B. Complete genome sequence of Collinsella aerofaciens Isolated from the gut of a healthy Indian subject. Genome Announc. 2017, 5, e01361-17. [Google Scholar] [CrossRef] [Green Version]
- Tap, J.; Furet, J.P.; Bensaada, M.; Philippe, C.; Roth, H.; Rabot, S.; Lakhdari, O.; Lombard, V.; Henrissat, B.; Corthier, G.; et al. Gut microbiota richness promotes its stability upon increased dietary fibre intake in healthy adults. Environ. Microbiol. 2015, 17, 4954–4964. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Backhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Daniel, H.; Gholami, A.M.; Berry, D.; Desmarchelier, C.; Hahne, H.; Loh, G.; Mondot, S.; Lepage, P.; Rothballer, M.; Walker, A.; et al. High-fat diet alters gut microbiota physiology in mice. ISME J. 2014, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.R.; Ning, Y.; Yu, H.; Tang, N.J. A HPLC-Q-TOF-MS-based urinary metabolomic approach to identification of potential biomarkers of metabolic syndrome. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Barclay, K.D.; McKersie, B.D. Peroxidation reactions in plant membranes: Effects of free fatty acids. Lipids 1994, 29, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Coxam, V. Current data with inulin-type fructans and calcium, targeting bone health in adults. J. Nutr. 2007, 137, 2527S–2533S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, S.; Watzl, B. Inulin and oligofructose: Review of experimental data on immune modulation. J. Nutr. 2007, 137, 2563S–2567S. [Google Scholar] [CrossRef]
- Zoetendal, E.G.; Raes, J.; Van Den Bogert, B.; Arumugam, M.; Booijink, C.C.; Troost, F.J.; Bork, P.; Wels, M.; De Vos, W.M.; Kleerebezem, M. The human small intestinal microbiota is driven by rapid uptake and conversion of simple carbohydrates. ISME J. 2012, 6, 1415. [Google Scholar] [CrossRef]
- Kaakoush, N.O. Insights into the Role of Erysipelotrichaceae in the Human Host. Front. Cell. Infect. Microbiol. 2015, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Sunvold, G.D.; Fahey, G.C., Jr.; Merchen, N.R.; Reinhart, G.A. In vitro fermentation of selected fibrous substrates by dog and cat fecal inoculum: Influence of diet composition on substrate organic matter disappearance and short-chain fatty acid production. J. Anim. Sci. 1995, 73, 1110–1122. [Google Scholar] [CrossRef]
- Bueno, A.R.; Cappel, T.G.; Sunvold, G.D.; Moxley, R.A.; Reinhart, G.A.; Clemens, E.T. Feline colonic microbes and fatty acid transport: Effects of feeding cellulose, beet pulp and pectin/gum arabic fibers. Nutr. Res. 2000, 20, 1319–1328. [Google Scholar] [CrossRef]
- Sparkes, A.H.; Papasouliotis, K.; Sunvold, G.; Werrett, G.; Gruffydd-Jones, E.A.; Egan, K.; Gruffydd-Jones, T.J.; Reinhart, G. Effect of dietary supplementation with fructo-oligosaccharides on fecal flora of healthy cats. Am. J. Vet. Res. 1998, 59, 436–440. [Google Scholar]
- Groeneveld, E.A.; Kappert, H.J.; der Kuilen Van, J.; Beynen, A.C. Consumption of fructooligosaccharides and nitrogen excretion in cats. Int. J. Vitam. Nutr. Res. 2001, 71, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Saville, B.A.; Saville, S. Xylooligosaccharides and Arabinoxylanoligosaccharides and Their Application as Prebiotics. Appl. Food Biotechnol. 2018, 5, 121–130. [Google Scholar]
- Andersen, S.J.; Hennebel, T.; Gildemyn, S.; Coma, M.; Desloover, J.; Berton, J.; Tsukamoto, J.; Stevens, C.; Rabaey, K. Electrolytic membrane extraction enables production of fine chemicals from biorefinery sidestreams. Environ. Sci. Technol. 2014, 48, 7135–7142. [Google Scholar] [CrossRef] [PubMed]
- Zytkovicz, T.H.; Fitzgerald, E.F.; Marsden, D.; Larson, C.A.; Shih, V.E.; Johnson, D.M.; Strauss, A.W.; Comeau, A.M.; Eaton, R.B.; Grady, G.F. Tandem mass spectrometric analysis for amino, organic, and fatty acid disorders in newborn dried blood spots: A two-year summary from the New England newborn screening program. Clin. Chem. 2001, 47, 1945–1955. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.A.; Hesta, M.; Hollants, J.; Janssens, G.P.; Huys, G. Phylogenetic analysis of faecal microbiota from captive cheetahs reveals underrepresentation of Bacteroidetes and Bifidobacteriaceae. BMC Microbiol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhoutte, T.; Huys, G.; De Brandt, E.; Swings, J. Temporal stability analysis of the microbiota in human feces by denaturing gradient gel electrophoresis using universal and group-specific 16S rRNA gene primers. FEMS Microbiol. Ecol. 2004, 48, 437–446. [Google Scholar] [CrossRef]
- Camarinha-Silva, A.; Jáuregui, R.; Chaves-Moreno, D.; Oxley, A.P.; Schaumburg, F.; Becker, K.; Wos-Oxley, M.L.; Pieper, D.H. Comparing the anterior nare bacterial community of two discrete human populations using I llumina amplicon sequencing. Environ. Microbiol. 2014, 16, 2939–2952. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item. | Control | LXOS | HXOS | SEM | p Value |
|---|---|---|---|---|---|
| Shannon index | 1.94 | 1.91 | 2.23 | 0.11 | 0.077 |
| Simpson index | 0.78 | 0.758 | 0.838 | 0.03 | 0.185 |
| Inverse Simpson index | 4.86 | 5.05 | 6.49 | 0.583 | 0.061 |
| Pielou’s evenness | 0.499 | 0.463 | 0.536 | 0.023 | 0.271 |
| Species richness | 49.4 a | 61.6 ab | 64.8 b | 4.41 | 0.022 |
| Taxonomy | Control | LXOS | HXOS | p Value |
|---|---|---|---|---|
| Firmicutes | ||||
| Bacilli; Lactobacillales; Streptococcaceae; Streptococcus | 13.468 a | 28.510 b | 7.561 a | <0.001 |
| Bacilli; Lactobacillales; Lactobacillaceae; Lactobacillus | 7.044 a | 16.591 b | 11.403 ab | <0.001 |
| Clostridia; Clostridiales; Ruminococcaceae; unclassified genus | 1.523 a | 0.871 a | 4.522 b | <0.001 |
| Clostridia; Clostridiales; Lachnospiraceae; Blautia | 0.396 a | 1.077 b | 1.870 c | <0.001 |
| Erysipelotrichia; Erysipelotrichales; Erysipelotrichaceae; unclassified genus | 1.495 a | 2.193 a | 4.294 b | 0.001 |
| Clostridia; Clostridiales; Ruminococcaceae; Subdoligranulum; | 0.240 a | 0.398 a | 0.771 b | 0.001 |
| Erysipelotrichia; Erysipelotrichales; Erysipelotrichaceae; Catenibacterium | 1.669 b | 0.587 a | 1.178 ab | 0.010 |
| Clostridia; Clostridiales; Lachnospiraceae; Blautia | 2.024 a | 4.051 b | 2.085 a | 0.029 |
| Clostridia; Clostridiales; Lachnospiraceae; Blautia | 6.423 a | 12.713 b | 9.363 ab | 0.038 |
| Clostridia; Clostridiales; Lachnospiraceae; unclassified genus | 0.127 a | 0.399 ab | 0.505 b | 0.044 |
| Negativicutes; Selenomonadales; Veillonellaceae; Megasphaera | 1.337 b | 0.790 a | 0.819 a | 0.045 |
| Clostridia; Clostridiales; Peptostreptococcaceae; Clostridium XI | 2.198 a | 3.258 b | 3.094 b | 0.047 |
| Actinobacteria | ||||
| Actinobacteria; Coriobacteriales; Coriobacteriaceae; Collinsella | 1.705 a | 2.556 b | 4.348 b | <0.001 |
| Actinobacteria; Bifidobacteriales; Bifidobacteriaceae; Bifidobacterium | 0.506 b | 0.177 a | 0.241 a | <0.001 |
| Actinobacteria; Bifidobacteriales; Bifidobacteriaceae; Bifidobacterium | 14.403 b | 16.259 b | 4.466 a | 0.006 |
| Actinobacteria; Bifidobacteriales; Bifidobacteriaceae; Bifidobacterium | 19.854 b | 5.829 a | 7.224 a | 0.011 |
| Ingredient (%) | Control | LXOS | HXOS |
| Corn | 39.11 | 39.11 | 39.11 |
| Dehydrated poultry meat | 19.80 | 19.80 | 19.80 |
| Corn gluten meal | 9.90 | 9.90 | 9.90 |
| Poultry Fat | 6.93 | 6.93 | 6.93 |
| Rice | 4.95 | 4.95 | 4.95 |
| Soy | 4.95 | 4.95 | 4.95 |
| Hydrolyzed dehydrated poultry meat | 4.95 | 4.95 | 4.95 |
| Premix | 2.48 | 2.48 | 2.48 |
| Fishmeal | 2.23 | 2.23 | 2.23 |
| Liquid aroma | 1.98 | 1.98 | 1.98 |
| Yeast | 1.24 | 1.24 | 1.24 |
| Aroma powder | 0.50 | 0.50 | 0.50 |
| Cellulose | 1.00 | 0.96 | 0.60 |
| XOS | 0.00 | 0.040 | 0.40 |
| Analysis on DM basis (%) | Control | LXOS | HXOS |
| Crude protein | 33.90 | 32.89 | 36.22 |
| Crude fat | 11.49 | 11.56 | 12.14 |
| Crude ash | 6.80 | 6.26 | 5.90 |
| Crude fiber | 1.57 | 1.42 | 1.47 |
| Total dietary fiber | 10.50 | 10.93 | 11.51 |
| Nitrogen free extract | 46.24 | 47.87 | 44.27 |
Sample Availability: Small samples (a few milligrams) of the compounds 8, 12, and 13 are available from the authors. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, Y.; Debevere, S.; Bourgeois, H.; Ran, M.; Broeckx, B.J.G.; Vanhaecke, L.; Wiele, T.V.d.; Hesta, M. Dose-Dependent Effects of Dietary Xylooligosaccharides Supplementation on Microbiota, Fermentation and Metabolism in Healthy Adult Cats. Molecules 2020, 25, 5030. https://doi.org/10.3390/molecules25215030
Lyu Y, Debevere S, Bourgeois H, Ran M, Broeckx BJG, Vanhaecke L, Wiele TVd, Hesta M. Dose-Dependent Effects of Dietary Xylooligosaccharides Supplementation on Microbiota, Fermentation and Metabolism in Healthy Adult Cats. Molecules. 2020; 25(21):5030. https://doi.org/10.3390/molecules25215030
Chicago/Turabian StyleLyu, Yang, Sandra Debevere, Hermann Bourgeois, Mavis Ran, Bart J.G. Broeckx, Lynn Vanhaecke, Tom Van de Wiele, and Myriam Hesta. 2020. "Dose-Dependent Effects of Dietary Xylooligosaccharides Supplementation on Microbiota, Fermentation and Metabolism in Healthy Adult Cats" Molecules 25, no. 21: 5030. https://doi.org/10.3390/molecules25215030
APA StyleLyu, Y., Debevere, S., Bourgeois, H., Ran, M., Broeckx, B. J. G., Vanhaecke, L., Wiele, T. V. d., & Hesta, M. (2020). Dose-Dependent Effects of Dietary Xylooligosaccharides Supplementation on Microbiota, Fermentation and Metabolism in Healthy Adult Cats. Molecules, 25(21), 5030. https://doi.org/10.3390/molecules25215030