
Effect of Oligosaccharide Degree of Polymerization on the Induction of Xylan-Degrading Enzymes by Fusarium oxysporum f. sp. Lycopersici




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
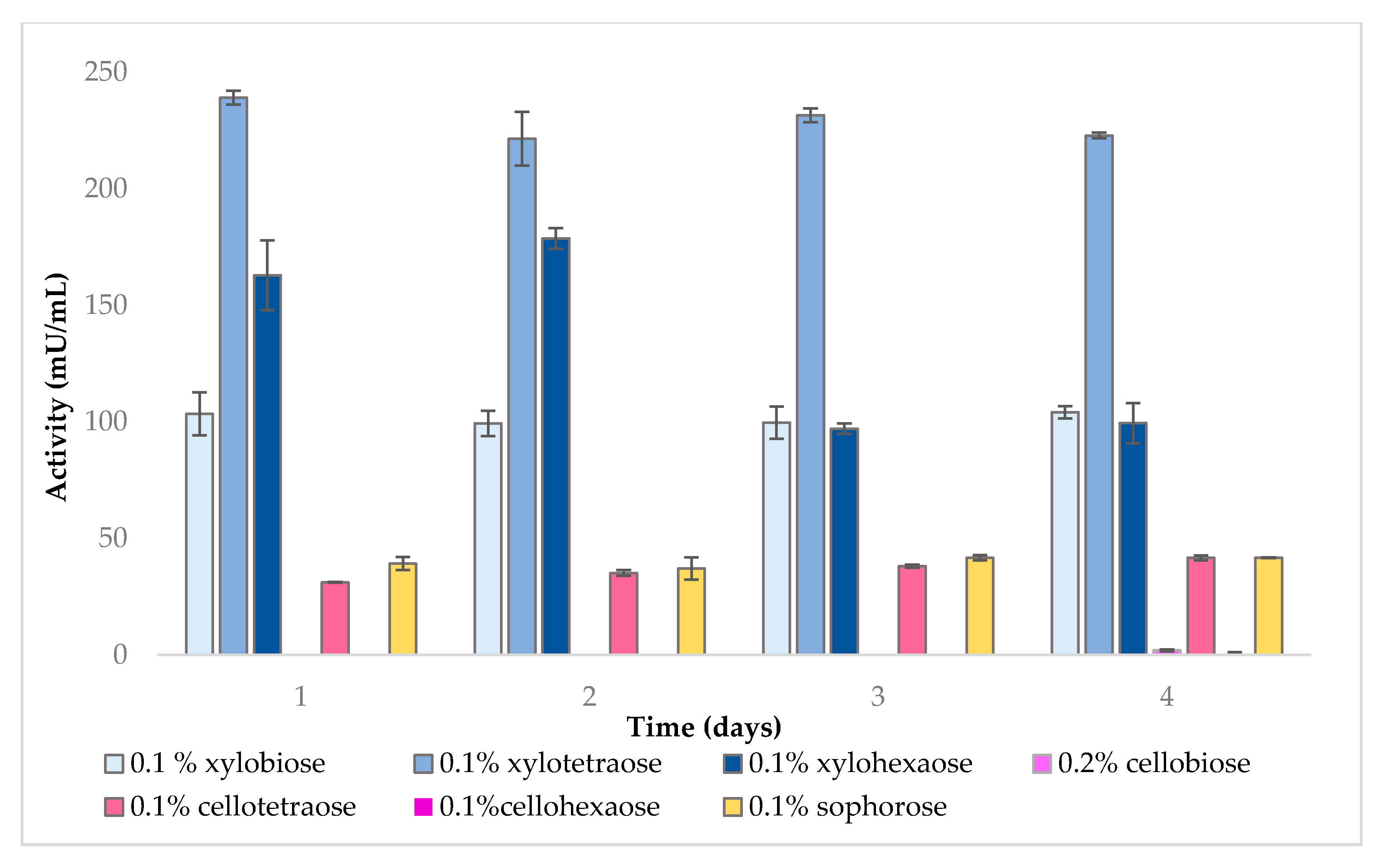
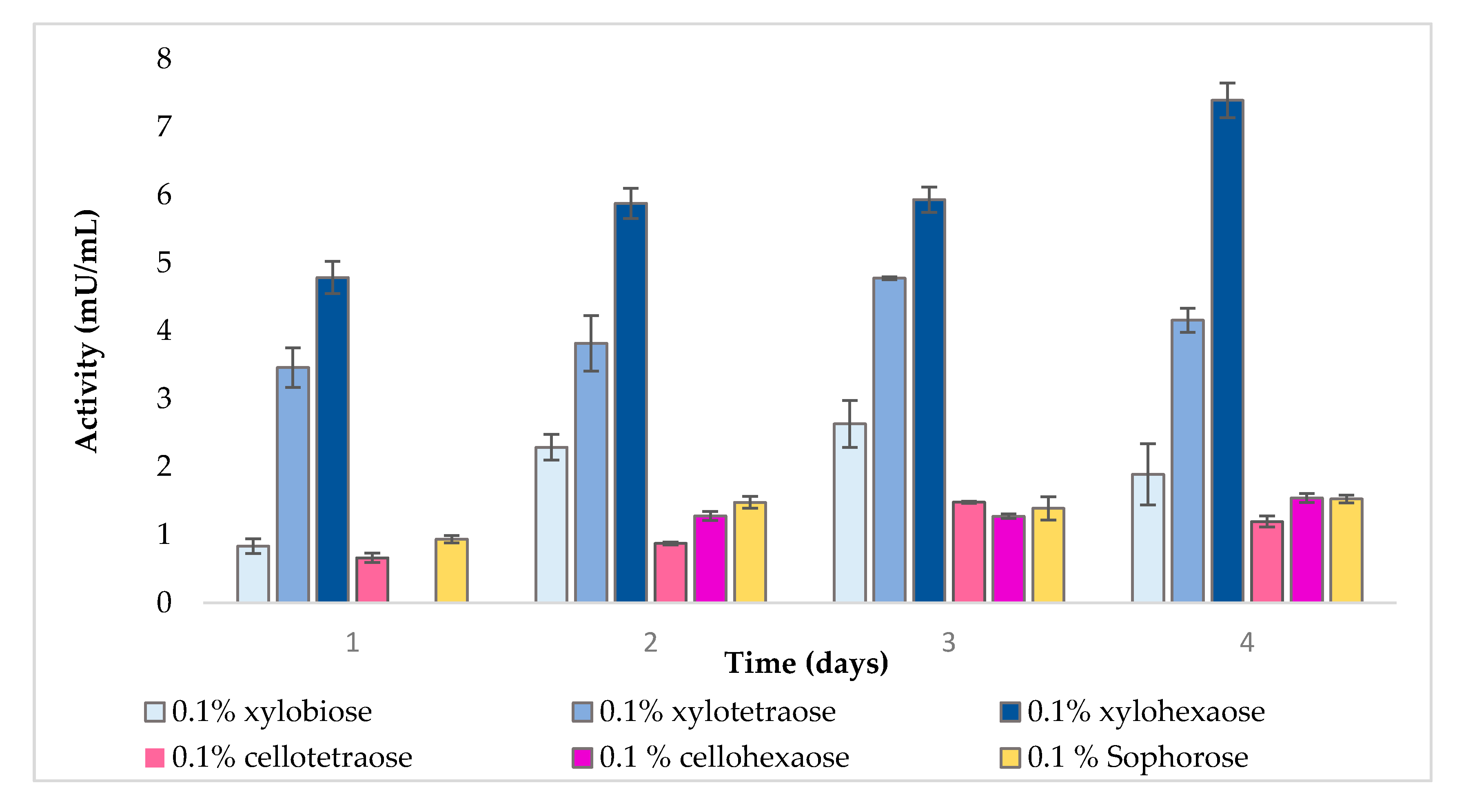
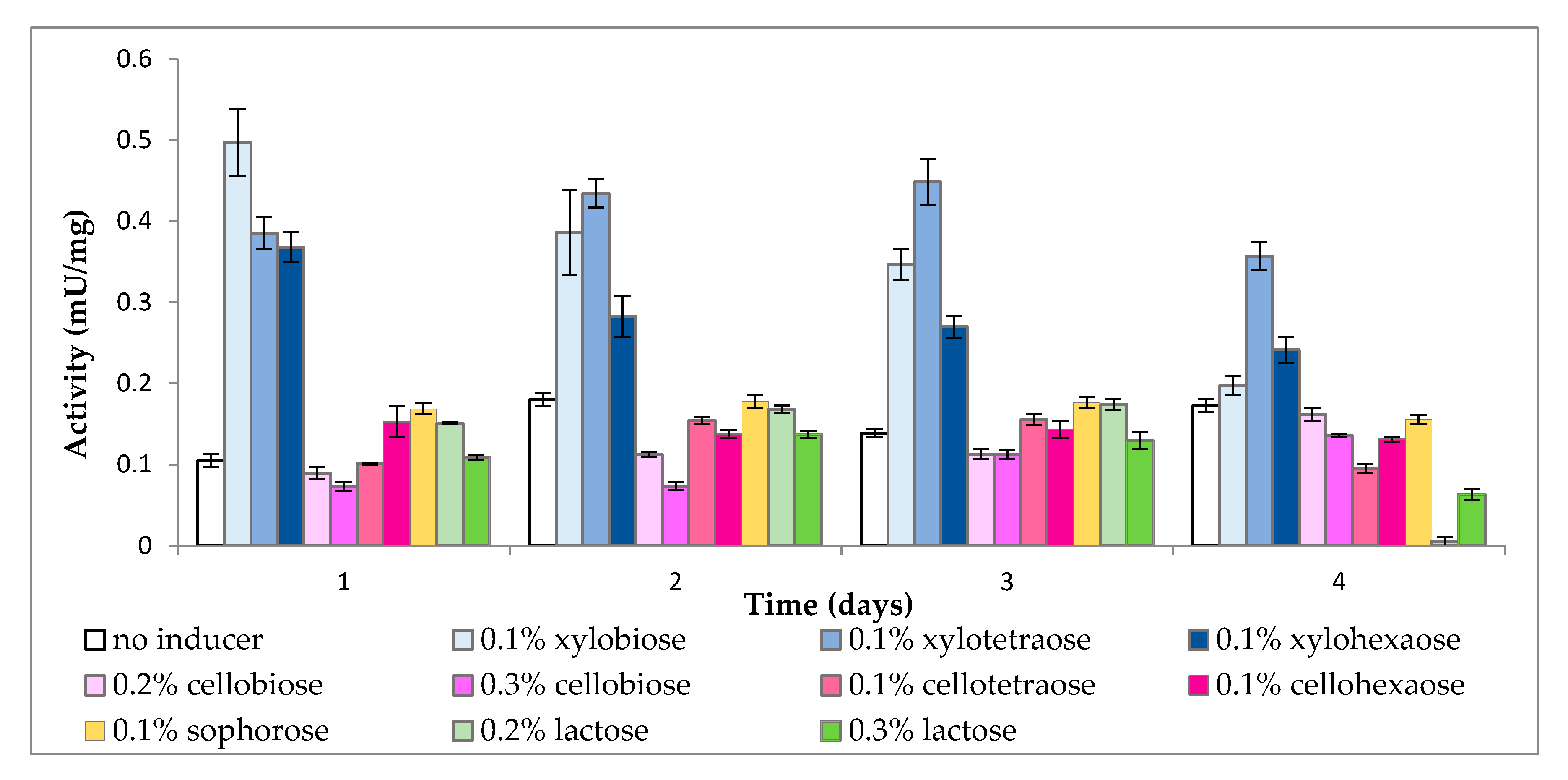
2.1. Effect of Inducers on Enzyme Activity
2.1.1. Endo-β-1,4-Xylanase
2.1.2. Extracellular β-Xylosidase
2.1.3. Cell-Bound β-Xylosidase
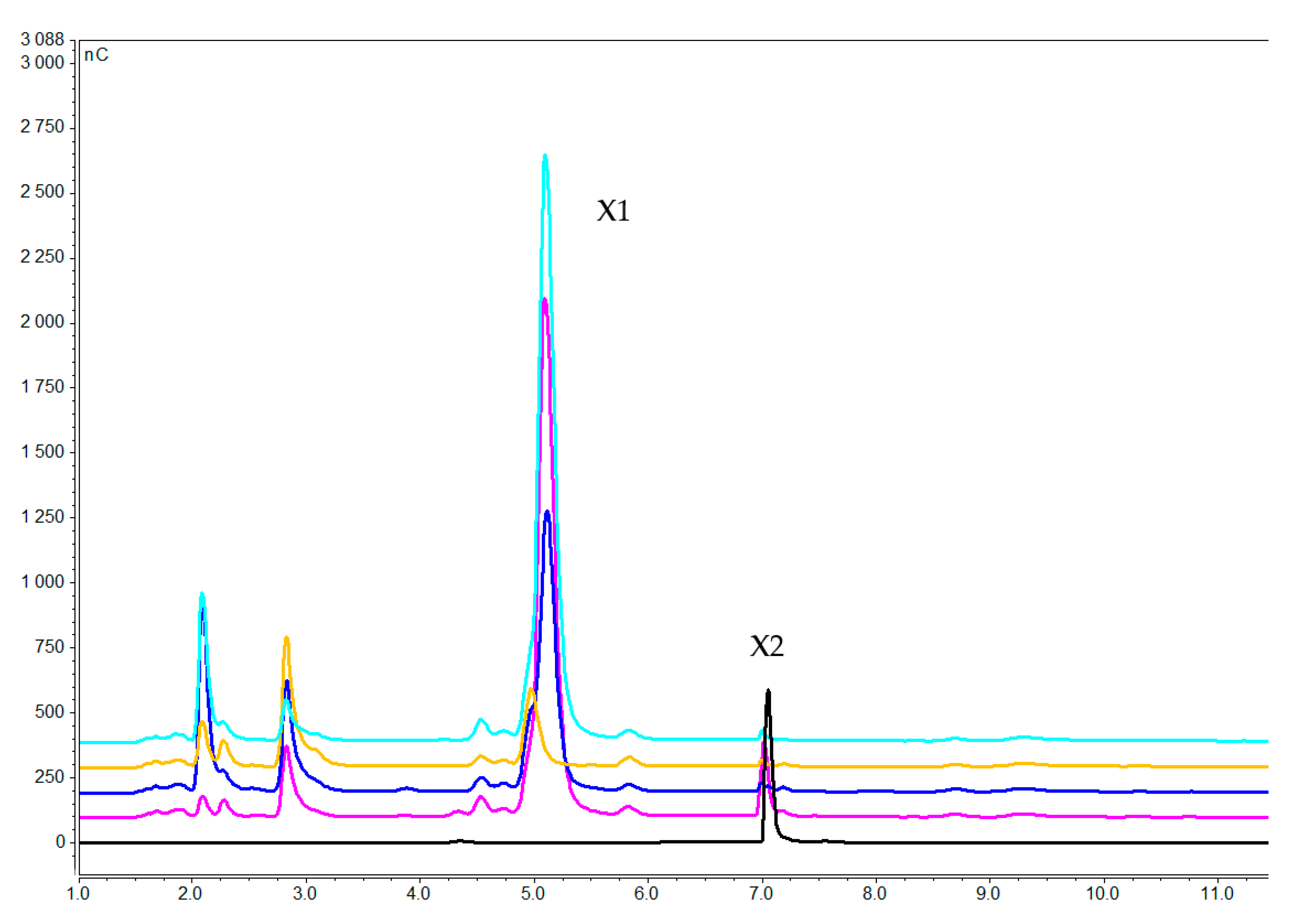
2.2. Uptake Experiment
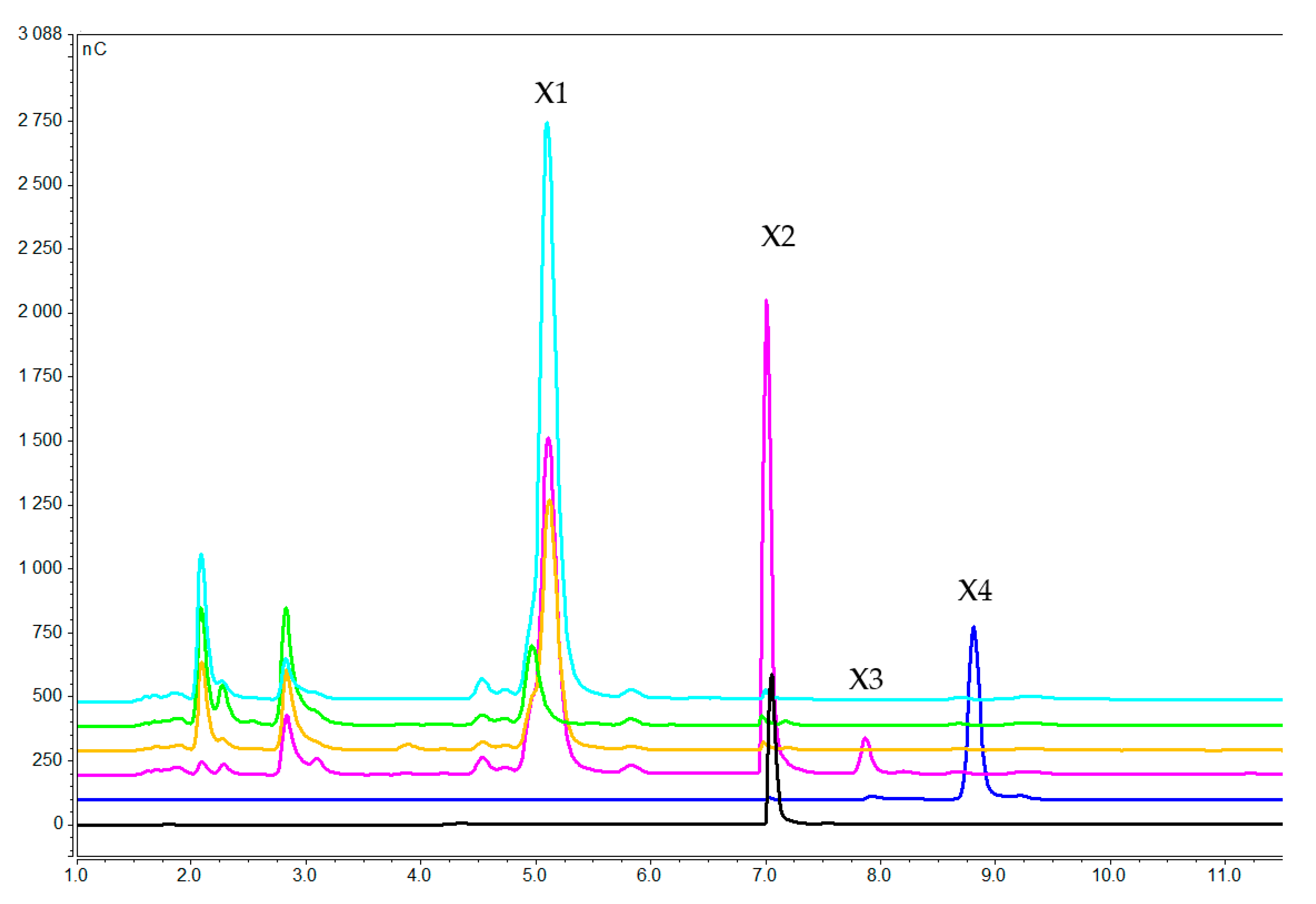
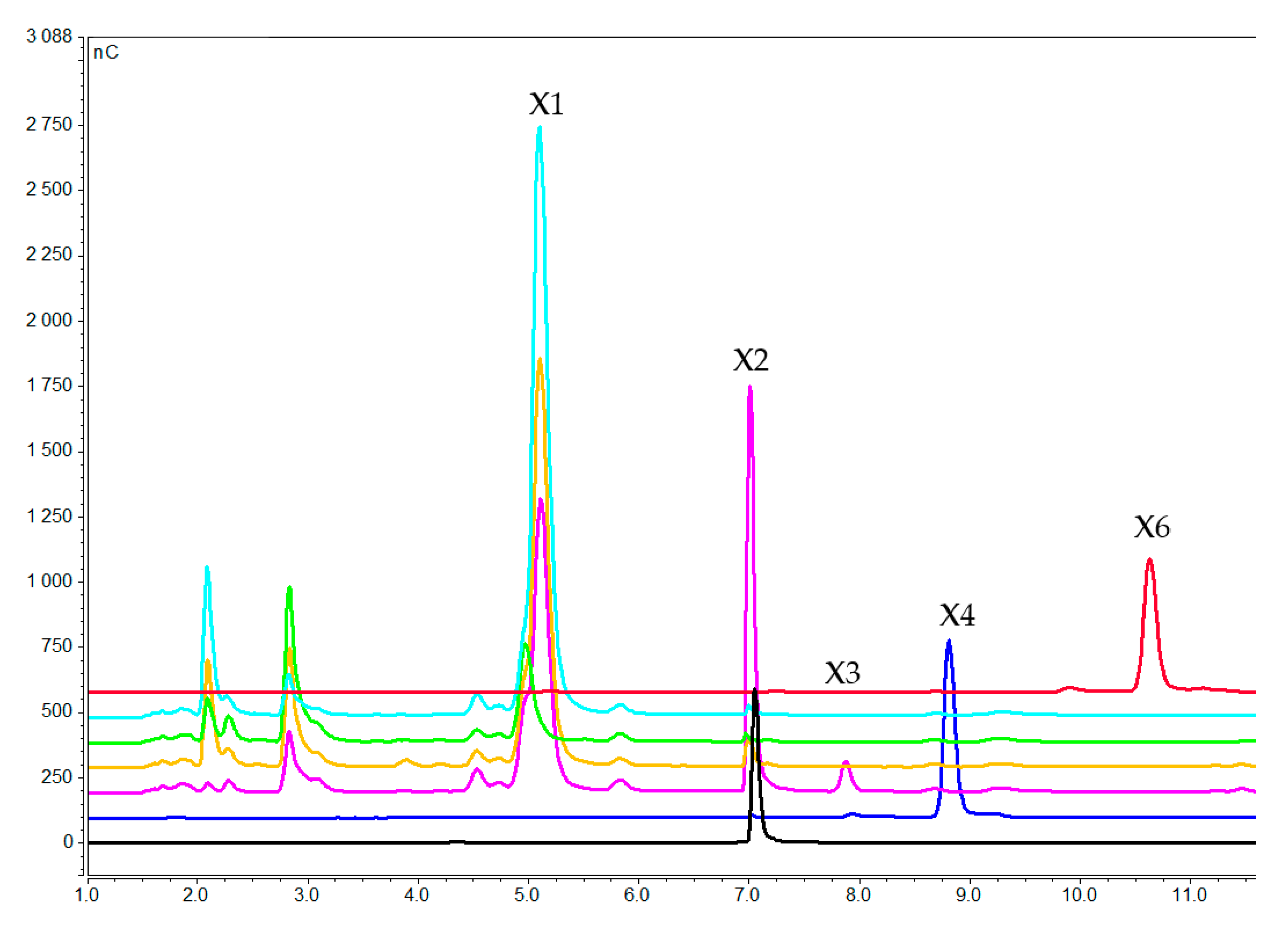
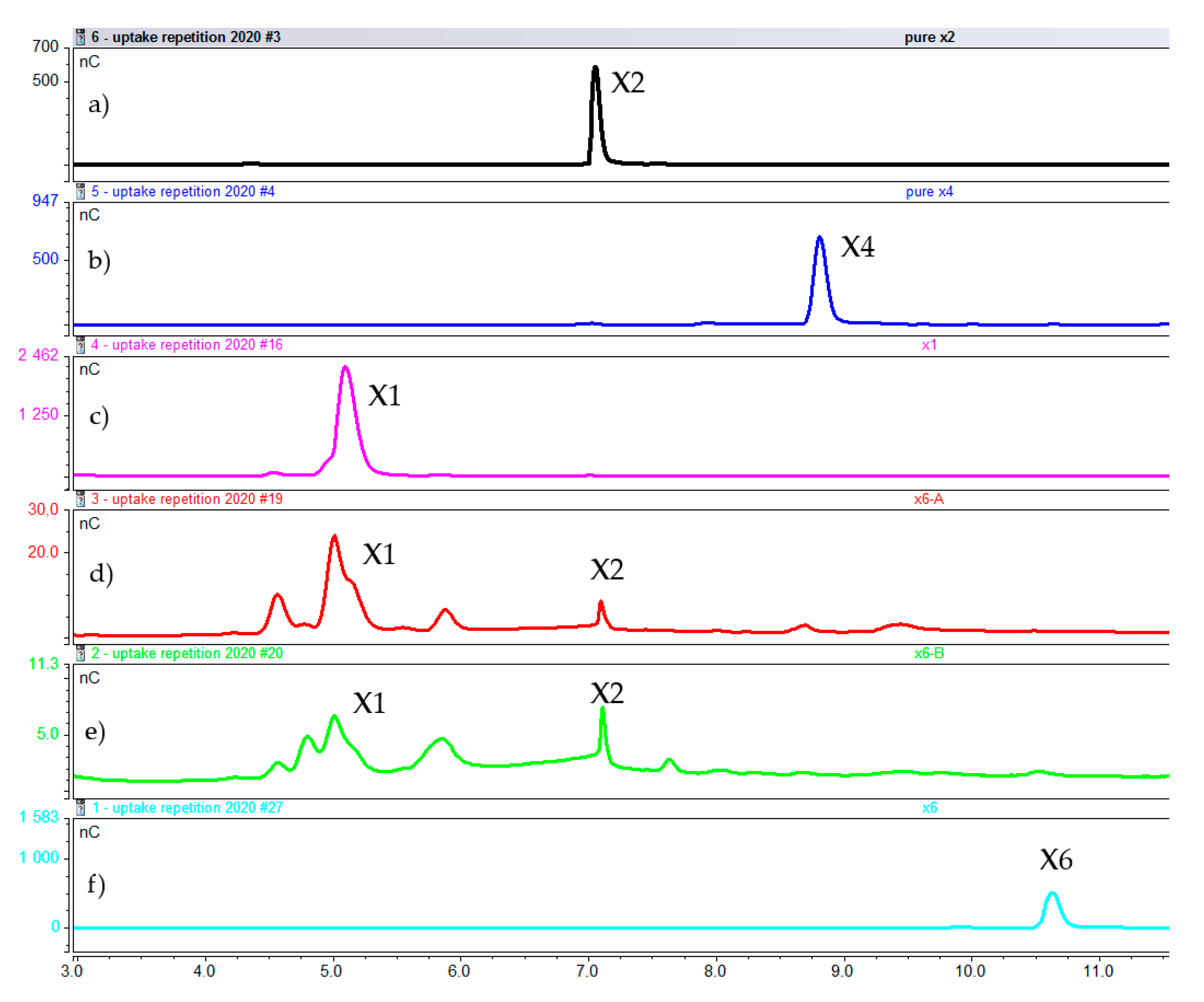
2.2.1. Concentration of Xylooligosaccharides in the Medium
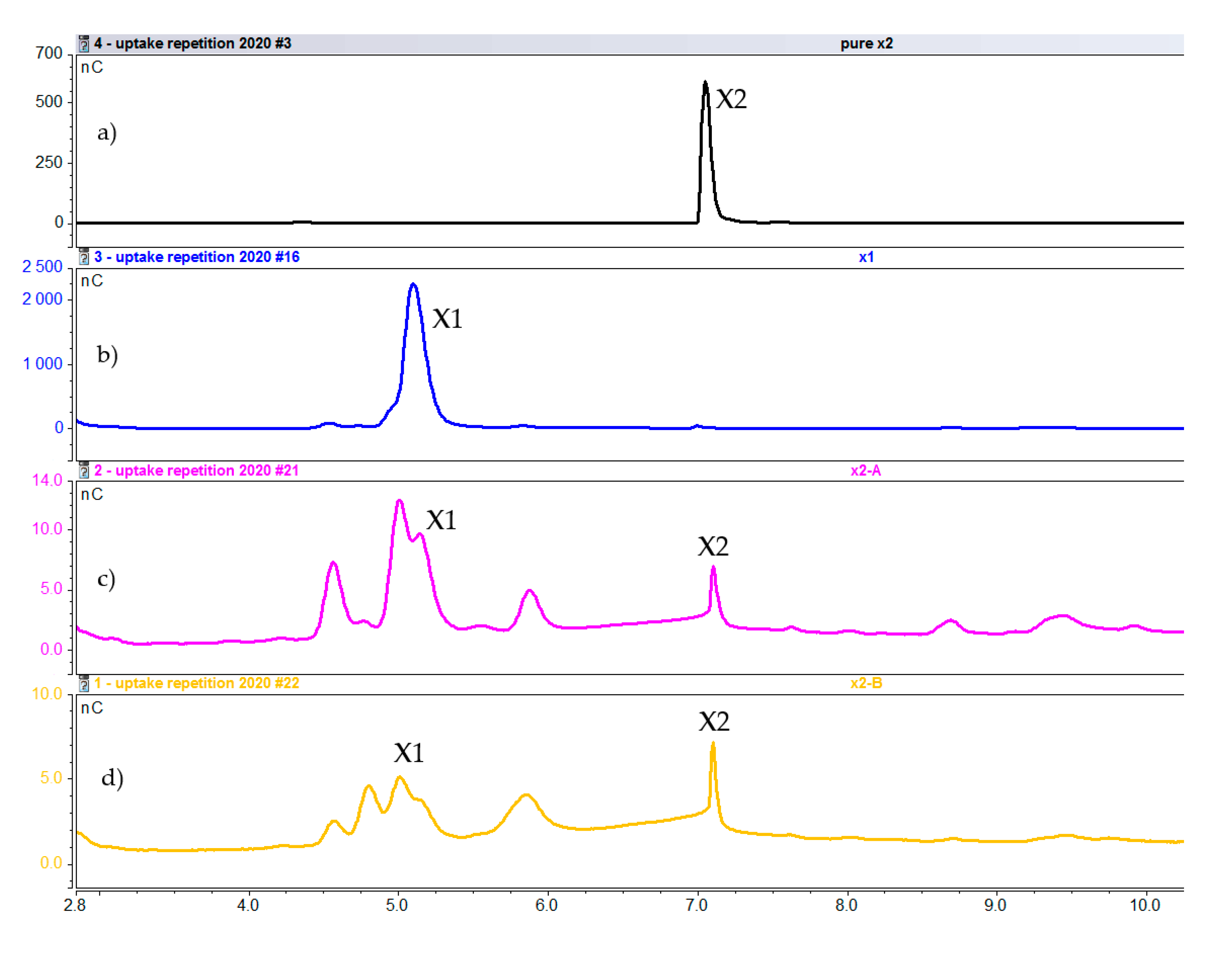
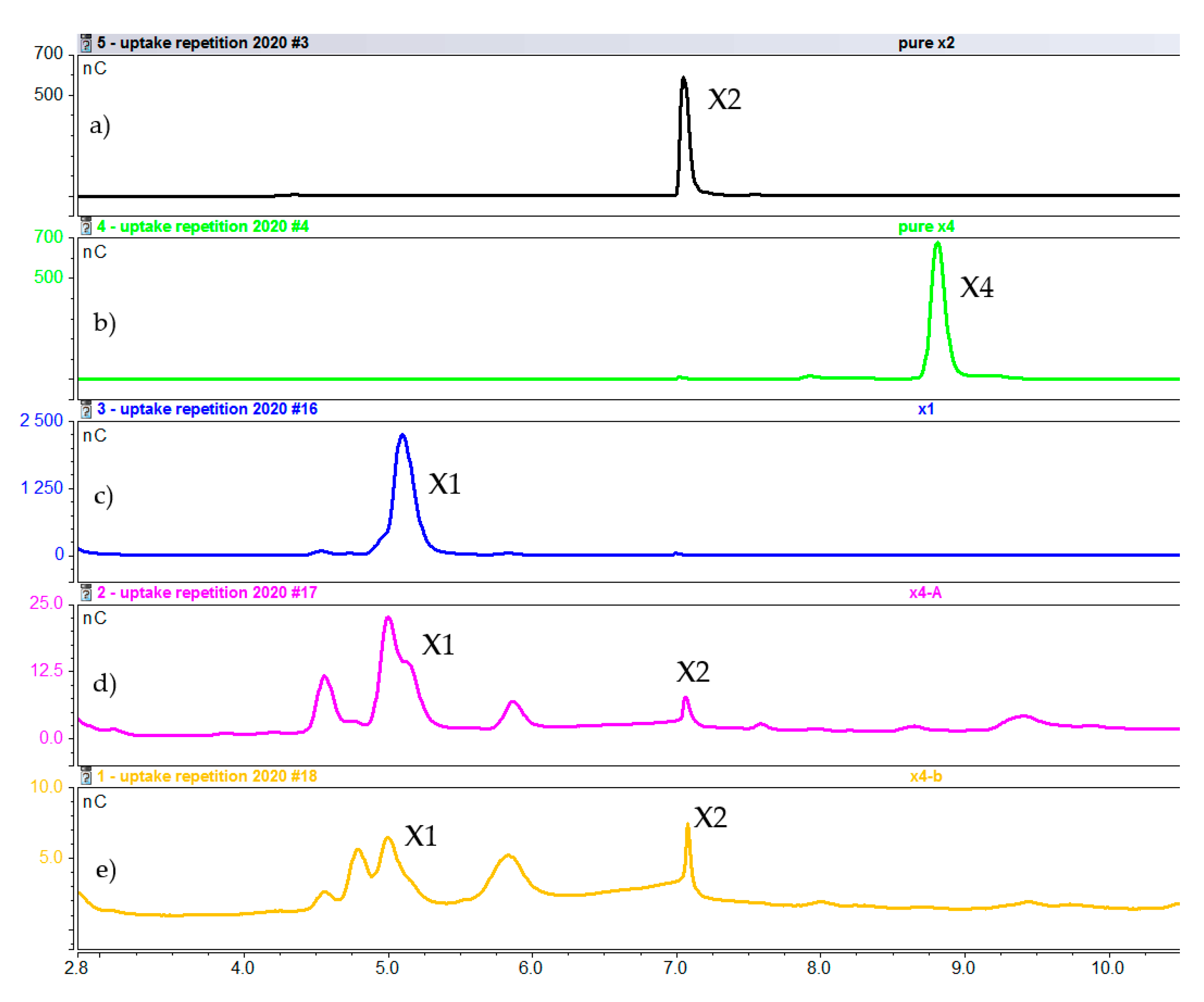
2.2.2. Intracellular Concentration of Xylooligosaccharides
3. Discussion
4. Materials and Methods
4.1. Microorganism and Pre-Cultures
4.2. Enzyme Induction Trials
4.3. Enzyme Activity Assessment
4.4. Xylooligosacharides Uptake Experiment
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prade, R.A. Xylanases: From Biology to BioTechnology. Biotechnol. Genet. Eng. Rev. 1996, 13, 101–132. [Google Scholar] [CrossRef] [PubMed]
- Beg, Q.K.; Kapoor, M.; Mahajan, L.; Hoondal, G.S. Microbial xylanases and their industrial applications: A review. Appl. Microbiol. Biotechnol. 2001, 56, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Wyman, C.; Decker, S.; Himmel, M.; Brady, J.; Skopec, C.; Viikari, L. Hydrolysis of Cellulose and Hemicellulose. Polysaccharides 2004, 1, 1023–1062. [Google Scholar]
- Wong, K.K.; Tan, L.U.; Saddler, J.N. Multiplicity of β-1,4-xylanase in microorganisms: Functions and applications. Microbiol. Rev. 1988, 52, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Madlala, A.M.; Prior, B.A. Thermomyces lanuginosus: Properties of strains and their hemicellulases. FEMS Microbiol. Rev. 2003, 27, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, N.; Shendye, A.; Rao, M. Molecular and biotechnological aspects of xylanases. FEMS Microbiol. Rev. 1999, 23, 411–456. [Google Scholar] [CrossRef]
- Chanda, S.K.; Hirst, E.L.; Jones, J.K.N.; Percival, E.G.V. 262. The constitution of xylan from esparto grass (stipa tenacissima, L.). J. Chem. Soc. 1950, 1289–1297. [Google Scholar] [CrossRef]
- Eda, S.; Ohnishi, A.; Kato, K. Xylan Isolated from the Stalk of Nicotiana tabacum. Agric. Biol. Chem. 1976, 40, 359–364. [Google Scholar] [CrossRef]
- Kenneth, J.H. Average Gestation Period and n π. Nat. Cell Biol. 1940, 146, 620. [Google Scholar] [CrossRef]
- Knob, A.; Terrasan, C.R.F.; Carmona, E.C. β-Xylosidases from filamentous fungi: An overview. World J. Microbiol. Biotechnol. 2010, 26, 389–407. [Google Scholar] [CrossRef]
- Katsimpouras, C.; Dedes, G.; Thomaidis, N.S.; Topakas, E. A novel fungal GH30 xylanase with xylobiohydrolase auxiliary activity. Biotechnol. Biofuels 2019, 12, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gielkens, M.M.C.; Visser, J.; De Graaff, L.H. Arabinoxylan degradation by fungi: Characterization of the arabinoxylan-arabinofuranohydrolase encoding genes from Aspergillus niger and Aspergillus tubingensis. Curr. Genet. 1997, 31, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Poutanen, K.; Sundberg, M.; Korte, H.; Puls, J. Deacetylation of xylans by acetyl esterases of Trichoderma reesei. Appl. Microbiol. Biotechnol. 1990, 33, 506–510. [Google Scholar] [CrossRef]
- Reilly, P.J. Xylanases: Structure and Function. Trends Biol. Ferment. Fuels Chem. 1981, 18, 111–129. [Google Scholar] [CrossRef]
- Li, W.; Wang, J.; Coluccio, L.M.; Matsudaira, P.; Grand, R.J. Brush border myosin I (BBMI): A basally localized transcript in human jejunal enterocytes. J. Histochem. Cytochem. 2000, 48, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajpai, P. Microbial Xylanolytic Systems and Their Properties. Xylanolytic Enzym. 2014, 19–36. [Google Scholar] [CrossRef]
- Poutanen, K.; Sundberg, M. An acetyl esterase of Trichoderma reesei and its role in the hydrolysis of acetyl xylans. Appl. Microbiol. Biotechnol. 1988, 28, 419–424. [Google Scholar] [CrossRef]
- Sørensen, H.R.; Meyer, A.S.; Pedersen, S. Enzymatic hydrolysis of water-soluble wheat arabinoxylan. 1. Synergy between α-l-arabinofuranosidases, endo-1,4-β-xylanases, and β-xylosidase activities. Biotechnol. Bioeng. 2003, 81, 726–731. [Google Scholar] [CrossRef]
- Rasmussen, L.E.; Sørensen, H.R.; Vind, J.; Viksø-Nielsen, A. Mode of action and properties of the β-xylosidases fromTalaromyces emersonii andTrichoderma reesei. Biotechnol. Bioeng. 2006, 94, 869–876. [Google Scholar] [CrossRef]
- Sunna, A.; Antranikian, G. Xylanolytic Enzymes from Fungi and Bacteria. Crit. Rev. Biotechnol. 1997, 17, 39–67. [Google Scholar] [CrossRef]
- Hrmova, M.; Biely, P.; Vršanská, M.; Petráková, E. Induction of cellulose- and xylan-degrading enzyme complex in the yeast Trichosporon cutaneum. Arch. Microbiol. 1984, 138, 371–376. [Google Scholar] [CrossRef]
- Liu, W.; Zhu, W.; Lu, Y.; Kong, J.; Ma, G. Production, partial purification and characterization of xylanase from Trichosporon cutaneum SL409. Process. Biochem. 1998, 33, 331–336. [Google Scholar] [CrossRef]
- Gilbert, J.; Hazlewood, P. Review Article Bacterial cellulases and xylanases. J. Gen. Microbiol. 1993, 1, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Lakshmi, G.S.; Bhargavi, P.L.; Prakasham, R.S. Sustainable Bioprocess Evaluation for Xylanase Production by Isolated Aspergillus terreus and Aspergillus fumigatus Under Solid—State Fermentation Using Oil Palm Empty Fruit Bunch Fiber. Curr. Trends Biotechnol. Pharm. 2011, 5, 1434–1444. [Google Scholar]
- Pathak, P.; Bhardwaj, N.K.; Singh, A.K. Production of Crude Cellulase and Xylanase From Trichoderma harzianum PPDDN10 NFCCI-2925 and Its Application in Photocopier Waste Paper Recycling. Appl. Biochem. Biotechnol. 2014, 172, 3776–3797. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G.N. Chapter Fourteen—Plant Diseases Caused by Viruses. In Agrios GNBT-PP, 5th ed.; Academic Press: San Diego, CA, USA, 2005; pp. 723–824. [Google Scholar] [CrossRef]
- Bertonha, L.C.; Neto, M.L.; Garcia, J.A.A.; Vieira, T.F.; Castoldi, R.; Bracht, A.; Peralta, R.M. Screening of Fusarium sp. for xylan and cellulose hydrolyzing enzymes and perspectives for the saccharification of delignified sugarcane bagasse. Biocatal. Agric. Biotechnol. 2018, 16, 385–389. [Google Scholar] [CrossRef]
- Panagiotou, G.; Kekos, D.; Macris, B.J.; Christakopoulos, P. Production of cellulolytic and xylanolytic enzymes by Fusarium oxysporum grown on corn stover in solid state fermentation. Ind. Crop. Prod. 2003, 18, 37–45. [Google Scholar] [CrossRef]
- Calero-Nieto, F.; Di Pietro, A.; Roncero, M.I.G.; Hera, C. Role of the Transcriptional Activator XlnR of Fusarium oxysporum in Regulation of Xylanase Genes and Virulence. Mol. Plant Microbe Interact. 2007, 20, 977–985. [Google Scholar] [CrossRef] [Green Version]
- Linko, M.; Viikari, L.; Suihko, M.-L. Hydrolysis of xylan and fermentation of xylose to ethanol. Biotechnol. Adv. 1984, 2, 233–252. [Google Scholar] [CrossRef]
- Christakopoulos, P.; Macris, B.; Kekos, D. Direct fermentation of cellulose to ethanol by Fusarium oxysporum. Enzym. Microb. Technol. 1989, 11, 236–239. [Google Scholar] [CrossRef]
- Dinolfo, M.I.; Eliana, C.; Stenglein, S.A. Fusarium–plant interaction: State of the art—A review. Plant Prot. Sci. 2017, 53, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Paccanaro, M.C.; Sella, L.; Castiglioni, C.; Giacomello, F.; Martínez-Rocha, A.L.; D’Ovidio, R.; Schã Fer, W.; Favaron, F. Synergistic Effect of Different Plant Cell Wall–Degrading Enzymes Is Important for Virulence of Fusarium graminearum. Mol. Plant Microbe Interact. 2017, 30, 886–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, R.P.; Visser, J.; De Graaff, L.H. CreA modulates the XlnR-induced expression on xylose of Aspergillus niger genes involved in xylan degradation. Res. Microbiol. 1999, 150, 281–285. [Google Scholar] [CrossRef]
- Kuhad, R.C.; Manchanda, M.; Singh, A. Optimization of xylanase production by a hyperxylanolytic mutant strain of Fusarium oxysporum. Process. Biochem. 1998, 33, 641–647. [Google Scholar] [CrossRef]
- Christakopoulos, P.; Mamma, D.; Nerinckx, W.; Kekos, D.; Macris, B.; Claeyssens, M. Production and partial characterization of xylanase from Fusarium oxysporum. Bioresour. Technol. 1996, 58, 115–119. [Google Scholar] [CrossRef]
- Fontes, C.M.G.A.; Gilbert, H.J.; Hazlewood, G.P.; Clarke, J.; Prates, J.A.M.; McKie, V.A.; Nagy, T.; Fernandes, T.H.; Ferreira, L.M.A. A novel Cellvibrio mixtus family 10 xylanase that is both intracellular and expressed under non-inducing conditions The GenBank accession numbers for the sequences described in this paper are AF049493 and AF168359 for xynC and xynG, respectively. Microbiology 2000, 146, 1959–1967. [Google Scholar] [CrossRef] [Green Version]
- Teplitsky, A.; Shulami, S.; Moryles, S.; Shoham, Y.; Shoham, G. Crystallization and preliminary X-ray analysis of an intracellular xylanase from Bacillus stearothermophilus T-6. Acta Crystallogr. Sect. D Biol. Crystallogr. 2000, 56, 181–184. [Google Scholar] [CrossRef]
- Kamoldeen, A.A.; Leh, C.P.; Abdullah, W.N.W.; Lee, C.K. Assessment of the Effect of Easily-metabolised Carbon Supplements on Xylanase Production by Newly Isolated Trichoderma asperellum USM SD4 Cultivated on Oil Palm Empty Fruit Bunches. Bioresources 2016, 11, 9611–9627. [Google Scholar] [CrossRef] [Green Version]
- Subramaniyan, S.; Prema, P. Biotechnology of Microbial Xylanases: Enzymology, Molecular Biology, and Application. Crit. Rev. Biotechnol. 2002, 22, 33–64. [Google Scholar] [CrossRef]
- Dobrev, G.T.; Pishtiyski, I.G.; Stanchev, V.S.; Mircheva, R. Optimization of nutrient medium containing agricultural wastes for xylanase production by Aspergillus niger B03 using optimal composite experimental design. Bioresour. Technol. 2007, 98, 2671–2678. [Google Scholar] [CrossRef]
- De Vries, R.; van de Vondervoort, P.; Hendriks, L.; Van de Belt, M.; Visser, J. Regulation of the α-glucuronidase-encoding gene (aguA) from Aspergillus niger. Mol. Genet. Genom. 2002, 268, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Martiín-Urdiíroz, M.C.; Di Pietro, A.; Huertas-González, M.D.; Roncero, M.I. Two xylanase genes of the vascular wilt pathogen Fusarium oxysporum are differentially expressed during infection of tomato plants. Mol. Genet. Genom. 1999, 261, 530–536. [Google Scholar] [CrossRef]
- Royer, J.; Nakas, J. Xylanase production by Trichoderma longibrachiatum. Enzym. Microb. Technol. 1989, 11, 405–410. [Google Scholar] [CrossRef]
- Raj, K.C.; Chandra, T.S. A cellulase-free xylanase from alkali-tolerantAspergillus fischeri Fxn1. Biotechnol. Lett. 1995, 17, 309–314. [Google Scholar] [CrossRef]
- Christakopoulos, P.; Nerinckx, W.; Kekos, D.; Macris, B.; Claeyssens, M. Purification and characterization of two low molecular mass alkaline xylanases from Fusarium oxysporum F3. J. Biotechnol. 1996, 51, 181–189. [Google Scholar] [CrossRef]
- Gaffney, M.; Doyle, S.; Murphy, R. Optimization of Xylanase Production byThermomyces lanuginosusin Solid State Fermentation. Biosci. Biotechnol. Biochem. 2009, 73, 2640–2644. [Google Scholar] [CrossRef] [Green Version]
- Khosravi, C.; Kowalczyk, J.E.; Chroumpi, T.; Battaglia, E.; Pontes, M.-V.A.; Peng, M.; Wiebenga, A.; Ng, V.; Lipzen, A.; He, G.; et al. Transcriptome analysis of Aspergillus niger xlnR and xkiA mutants grown on corn Stover and soybean hulls reveals a highly complex regulatory network. BMC Genom. 2019, 20, 1–16. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, H.; Lv, B.; Li, C. Regulating Strategies for Producing Carbohydrate Active Enzymes by Filamentous Fungal Cell Factories. Front. Bioeng. Biotechnol. 2020, 8, 691. [Google Scholar] [CrossRef]
- Dowzer, C.E.A.; Kelly, J.M. Cloning of the creA gene from Aspergillus nidulans: A gene involved in carbon catabolite repression. Curr. Genet. 1989, 15, 457–459. [Google Scholar] [CrossRef]
- Strauss, J.; Mach, R.L.; Zeilinger, S.; Hartler, G.; Stöffler, G.; Wolschek, M.; Kubicek, C.P. Crel, the carbon catabolite repressor protein from Trichoderma reesei. FEBS Lett. 1995, 376, 103–107. [Google Scholar] [CrossRef] [Green Version]
- De La Serna, I.; Ng, D.; Tyler, B.M. Carbon Regulation of Ribosomal Genes in Neurospora crassa Occurs by a Mechanism Which Does Not Require Cre-1, the Homologue of the Aspergillus Carbon Catabolite Repressor, CreA. Fungal Genet. Biol. 1999, 26, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Todd, R.B.; Lockington, R.A.; Kelly, J.M. The Aspergillus nidulans creC gene involved in carbon catabolite repression encodes a WD40 repeat protein. Mol. Genet. Genom. 2000, 263, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Lockington, R.A.; Kelly, J.M. The WD40-repeat protein CreC interacts with and stabilizes the deubiquitinating enzyme CreB in vivo in Aspergillus nidulans. Mol. Microbiol. 2002, 43, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, L.N.A.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse Regulation of the CreA Carbon Catabolite Repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef] [Green Version]
- Christakopoulos, P.; Nerinckx, W.; Kekos, D.; Macris, B.; Claeyssens, M. The alkaline xylanase III from Fusarium oxysporum F3 belongs to family F/10. Carbohydr. Res. 1997, 302, 191–195. [Google Scholar] [CrossRef]
- Chaudhuri, B.; Sahai, V. Comparison of growth and maintenance parameters for cellulase biosynthesis by Trichoderma reesei-C5 with some published data. Enzym. Microb. Technol. 1994, 16, 1079–1083. [Google Scholar] [CrossRef]
- Xu, J.; Nogawa, M.; Okada, H.; Morikawa, Y. Regulation of xyn3 gene expression in Trichoderma reesei PC-3-7. Appl. Microbiol. Biotechnol. 2000, 54, 370–375. [Google Scholar] [CrossRef]
- Xiong, H.; Turunen, O.; Pastinen, O.; Leisola, M.; Von Weymarn, N. Improved xylanase production by Trichoderma reesei grown on l-arabinose and lactose or d-glucose mixtures. Appl. Microbiol. Biotechnol. 2004, 64, 353–358. [Google Scholar] [CrossRef]
- Kang, S.-W.; Kim, S.W.; Lee, J.-S. Production of cellulase and xylanase in a bubble column using immobilizedAspergillus Niger KKS. Appl. Biochem. Biotechnol. 1995, 53, 101–106. [Google Scholar] [CrossRef]
- Wang, P.; Mason, J.C.; Broda, P. Xylanases from Streptomyces cyaneus: Their production, purification and characterization. J. Gen. Microbiol. 1993, 139, 1987–1993. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chany, C.J.; Sims, P.F.; Sinnott, M.L. Definition of the substrate specificity of the ‘sensing’ xylanase of Streptomyces cyaneus using xylooligosaccharide and cellooligosaccharide glycosides of 3,4-dinitrophenol. J. Biotechnol. 1997, 57, 181–190. [Google Scholar] [CrossRef]
- Biely, P. Microbial xylanolytic systems. Trends Biotechnol. 1985, 3, 286–290. [Google Scholar] [CrossRef]
- Gomes, D.; Gomes, J.; Steiner, W. Factors influencing the induction of endo-xylanase by Thermoascus aurantiacus. J. Biotechnol. 1994, 33, 87–94. [Google Scholar] [CrossRef]
- Rapp, P.; Wagner, F. Production and Properties of Xylan-Degrading Enzymes from Cellulomonas uda. Appl. Environ. Microbiol. 1986, 51, 746–752. [Google Scholar] [CrossRef] [Green Version]
- La Grange, D.C.; Claeyssens, M.; Pretorius, I.S.; Van Zyl, W.H. Coexpression of the Bacillus pumilus β-xylosidase ( xynB ) gene with the Trichoderma reesei β-xylanase 2 ( xyn2 ) gene in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2000, 54, 195–200. [Google Scholar] [CrossRef]
- De Paula, R.G.; Antoniêto, A.C.C.; Ribeiro, L.F.C.; Carraro, C.B.; Nogueira, K.M.V.; Lopes, D.C.B.; Silva, A.C.; Zerbini, M.T.; Pedersoli, W.R.; Costa, M.D.N.; et al. New Genomic Approaches to Enhance Biomass Degradation by the Industrial Fungus Trichoderma reesei. Int. J. Genom. 2018, 2018, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Gu, R.; Wang, B.; Li, J.; Wan, L.; Tian, C.; Ma, Y. Evidence of a Critical Role for Cellodextrin Transporte 2 (CDT-2) in Both Cellulose and Hemicellulose Degradation and Utilization in Neurospora crassa. PLoS ONE 2014, 9, e89330. [Google Scholar] [CrossRef]
- Porciuncula, J.D.O.; Furukawa, T.; Shida, Y.; Mori, K.; Kuhara, S.; Morikawa, Y.; Ogasawara, W. Identification of Major Facilitator Transporters Involved in Cellulase Production during Lactose Culture ofTrichoderma reeseiPC-3-7. Biosci. Biotechnol. Biochem. 2013, 77, 1014–1022. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, C.; Bååth, J.A.; Seiboth, B.; Kubicek, C.P. Systems Analysis of Lactose Metabolism in Trichoderma reesei Identifies a Lactose Permease That Is Essential for Cellulase Induction. PLoS ONE 2013, 8, e62631. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, N.; Kumari, I.; Sandhu, P.; Ahmed, M.; Akhter, Y. Proteome scale census of major facilitator superfamily transporters in Trichoderma reesei using protein sequence and structure based classification enhanced ranking. Gene 2016, 585, 166–176. [Google Scholar] [CrossRef]
- Schmoll, M.; Dattenböck, C.; Carreras-Villaseñor, N.; Mendoza-Mendoza, A.; Tisch, D.; Alemán, M.I.; Baker, S.E.; Brown, C.; Cervantes-Badillo, M.G.; Cetz-Chel, J.; et al. The Genomes of Three Uneven Siblings: Footprints of the Lifestyles of Three Trichoderma Species. Microbiol. Mol. Biol. Rev. 2016, 80, 205–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, K.M.V.; Mendes, V.; Carraro, C.B.; Taveira, I.C.; Oshiquiri, L.H.; Gupta, V.K.; Silva, R.N. Sugar transporters from industrial fungi: Key to improving second-generation ethanol production. Renew. Sustain. Energy Rev. 2020, 131, 109991. [Google Scholar] [CrossRef]
- Fravel, D.; Moravec, B.; Jones, R.W.; Costanzo, S. Characterization of two ABC transporters from biocontrol and phytopathogenic Fusarium oxysporum. Physiol. Mol. Plant Pathol. 2008, 73, 2–8. [Google Scholar] [CrossRef]
- Atanasova, L.; Le Crom, S.; Gruber, S.; Coulpier, F.; Seidl-Seiboth, V.; Kubicek, C.P.; Druzhinina, I.S. Comparative transcriptomics reveals different strategies of Trichoderma mycoparasitism. BMC Genom. 2013, 14, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Hwang, J.-U.; Lee, M.; Kim, Y.-Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef] [Green Version]
- Henderson, R.K.; Fendler, K.; Poolman, B. Coupling efficiency of secondary active transporters. Curr. Opin. Biotechnol. 2019, 58, 62–71. [Google Scholar] [CrossRef]
- Baral, B. Chapter Four—Evolutionary Trajectories of Entomopathogenic Fungi ABC Transporters. In Goodwin SFBT-A in G; Friedmann, T., Dunlap, J.C., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 117–154. [Google Scholar] [CrossRef]
- Vongsangnak, W.; Salazar, M.; Hansen, K.; Nielsen, J. Genome-wide analysis of maltose utilization and regulation in aspergilli. Microbiology 2009, 155, 3893–3902. [Google Scholar] [CrossRef] [Green Version]
- Galagan, J.E.; Calvo, S.E.; Borkovich, K.A.; Selker, E.U.; Read, N.D.; Jaffe, D.; Fitzhugh, W.; Ma, L.-J.; Smirnov, S.; Purcell, S.; et al. The genome sequence of the filamentous fungus Neurospora crassa. Nature 2003, 422, 859–868. [Google Scholar] [CrossRef]
- Saier, M.H.; Beatty, J.T.; Goffeau, A.; Harley, K.T.; Heijne, W.H.M.; Huang, S.C.; Jack, D.L.; Jähn, P.S.; Lew, K.; Liu, J.; et al. The major facilitator superfamily. J. Mol. Microbiol. Biotechnol. 1999, 1, 257–279. [Google Scholar]
- Law, C.J.; Maloney, P.C.; Wang, D. Ins and Outs of Major Facilitator Superfamily Antiporters. Annu. Rev. Microbiol. 2008, 62, 289–305. [Google Scholar] [CrossRef] [Green Version]
- Yan, N. Structural advances for the major facilitator superfamily (MFS) transporters. Trends Biochem. Sci. 2013, 38, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Davejan, P.; Hendargo, K.J.; Javadi-Razaz, I.; Chou, A.; Yee, D.C.; Ghazi, F.; Lam, K.J.K.; Conn, A.M.; Madrigal, A.; et al. Expansion of the Major Facilitator Superfamily (MFS) to include novel transporters as well as transmembrane-acting enzymes. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183277. [Google Scholar] [CrossRef] [PubMed]
- Castro, S.; Pedersoli, W.R.; Cristina, A.; Antoniêto, C.; Steindorff, A.S.; Silva-rocha, R.; Martinez-Rossi, N.M.; Rossi, A.; Brown, N.A.; Goldman, G.H.; et al. Comparative metabolism of cellulose, sophorose and glucose in Trichoderma reesei using high-throughput genomic and proteomic analyses. Biotechnol. Biofuels 2014, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniêto, A.C.C.; De Paula, R.G.; Castro, L.D.S.; Silva-Rocha, R.; Persinoti, G.F.; Silva, R.N. Trichoderma reesei CRE1-mediated Carbon Catabolite Repression in Response to Sophorose Through RNA Sequencing Analysis. Curr. Genom. 2016, 17, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galazka, J.M.; Tian, C.; Beeson, W.T.; Martinez, B.; Glass, N.L.; Cate, J.H. Cellodextrin transport in yeast for improved biofuel production. Science 2010, 330, 84–86. [Google Scholar] [CrossRef]
- Li, X.; Yu, V.Y.; Lin, Y.; Chomvong, K.; Estrela, R.; Park, A.; Liang, J.M.; Znameroski, E.; Feehan, J.; Kim, S.R.; et al. Expanding xylose metabolism in yeast for plant cell wall conversion to biofuels. eLife 2015, 4, 1–55. [Google Scholar] [CrossRef]
- Miao, Y.; Li, J.; Xiao, Z.; Shen, Q.; Zhang, R. Characterization and identification of the xylanolytic enzymes from Aspergillus fumigatus Z5. BMC Microbiol. 2015, 15, 126. [Google Scholar] [CrossRef] [Green Version]
- Znameroski, E.A.; Coradetti, S.T.; Roche, C.M.; Tsai, J.C.; Iavarone, A.T.; Cate, J.H.D.; Glass, N.L. Induction of lignocellulose-degrading enzymes in Neurospora crassa by cellodextrins. Proc. Natl. Acad. Sci. USA 2012, 109, 6012–6017. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]









Sample Availability: Samples of the compounds are not available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Najjarzadeh, N.; Matsakas, L.; Rova, U.; Christakopoulos, P. Effect of Oligosaccharide Degree of Polymerization on the Induction of Xylan-Degrading Enzymes by Fusarium oxysporum f. sp. Lycopersici. Molecules 2020, 25, 5849. https://doi.org/10.3390/molecules25245849
Najjarzadeh N, Matsakas L, Rova U, Christakopoulos P. Effect of Oligosaccharide Degree of Polymerization on the Induction of Xylan-Degrading Enzymes by Fusarium oxysporum f. sp. Lycopersici. Molecules. 2020; 25(24):5849. https://doi.org/10.3390/molecules25245849
Chicago/Turabian StyleNajjarzadeh, Nasim, Leonidas Matsakas, Ulrika Rova, and Paul Christakopoulos. 2020. "Effect of Oligosaccharide Degree of Polymerization on the Induction of Xylan-Degrading Enzymes by Fusarium oxysporum f. sp. Lycopersici" Molecules 25, no. 24: 5849. https://doi.org/10.3390/molecules25245849
APA StyleNajjarzadeh, N., Matsakas, L., Rova, U., & Christakopoulos, P. (2020). Effect of Oligosaccharide Degree of Polymerization on the Induction of Xylan-Degrading Enzymes by Fusarium oxysporum f. sp. Lycopersici. Molecules, 25(24), 5849. https://doi.org/10.3390/molecules25245849