
Microbial Degradation of Hydrocarbons—Basic Principles for Bioremediation: A Review

 , ,
, ,  and
and 
Abstract
:1. Introduction
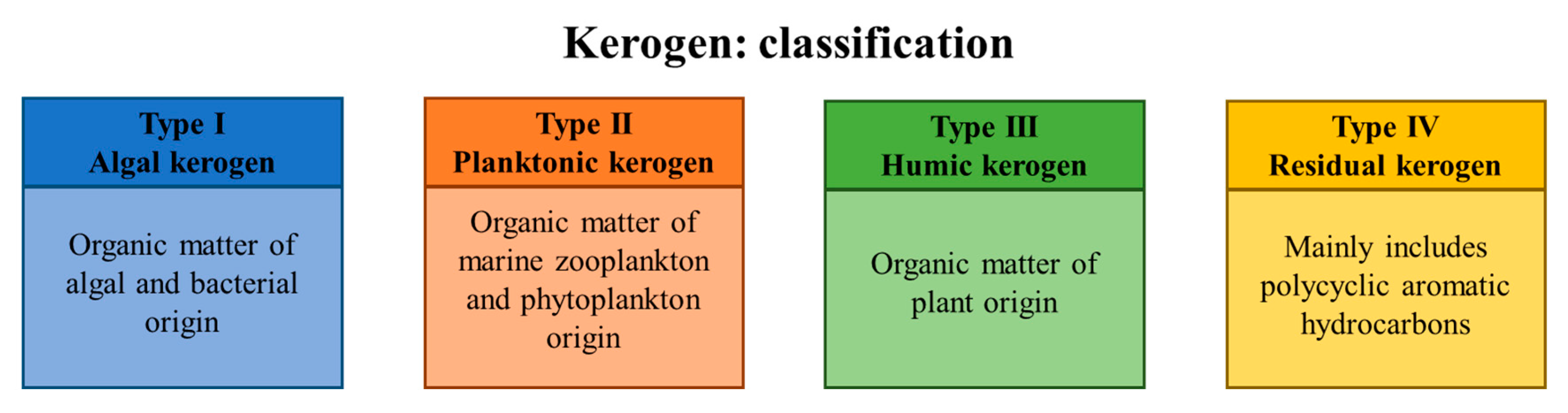
2. The Inseparable Bond between Hydrocarbons and Microbes—When, Where and How?
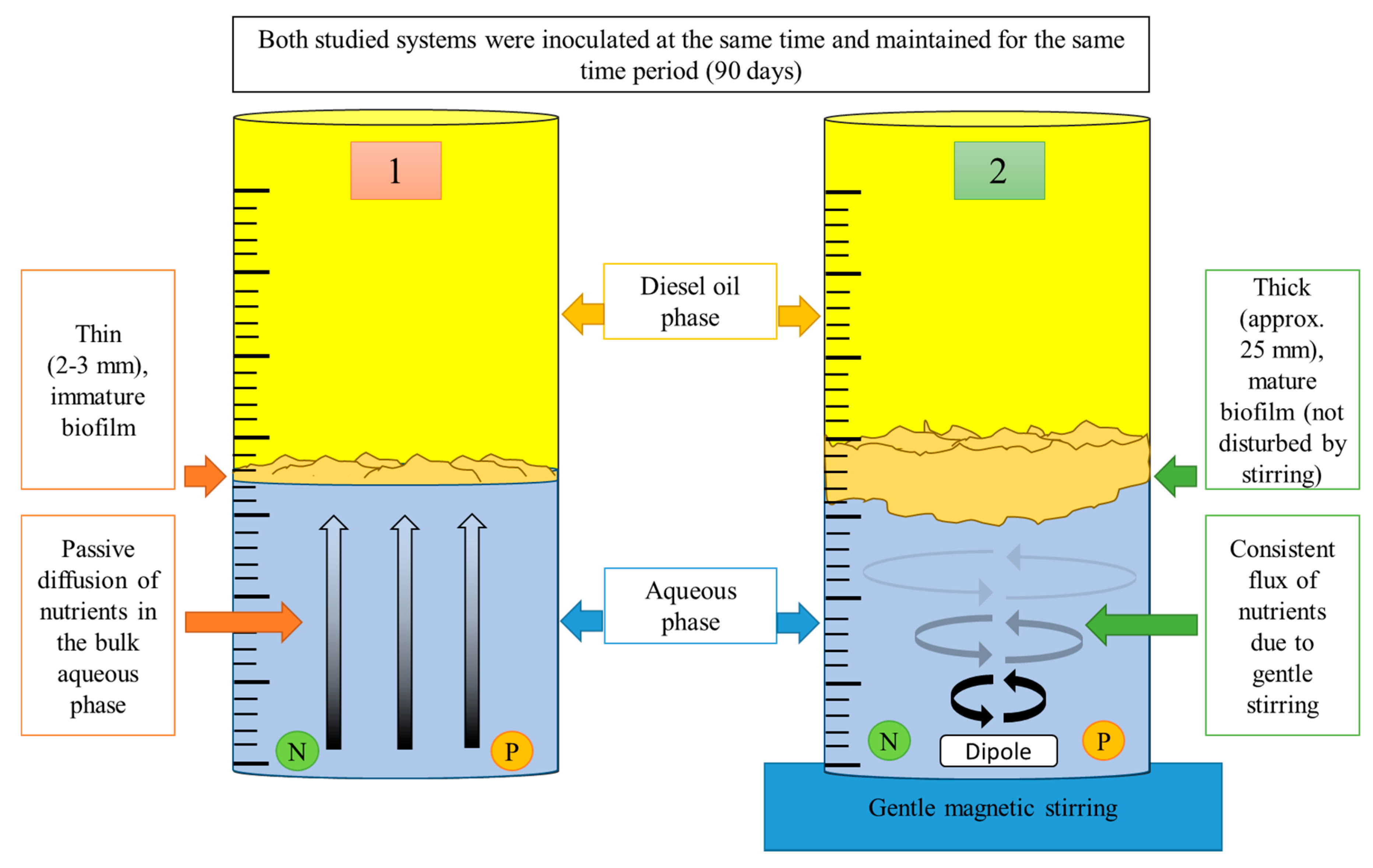
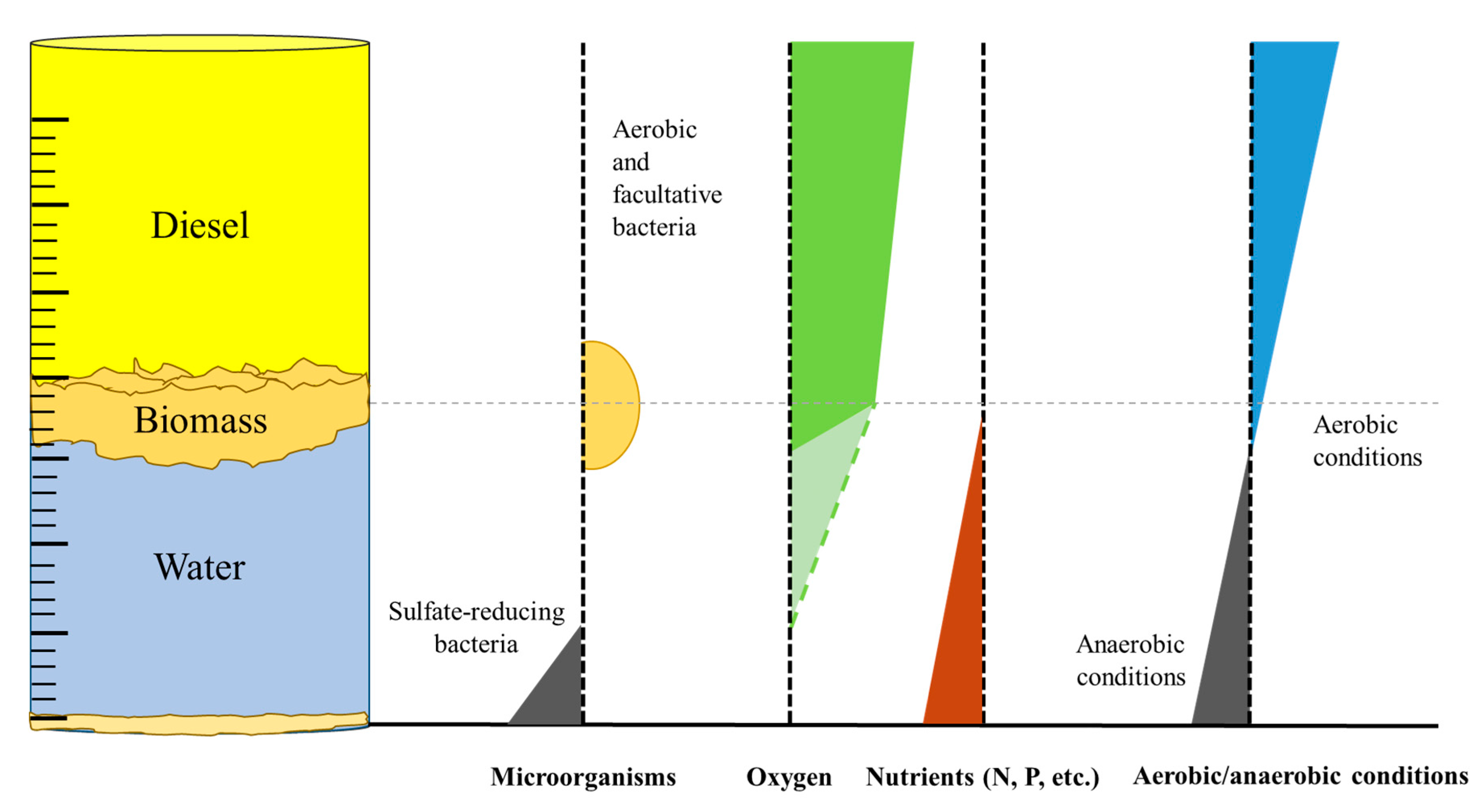
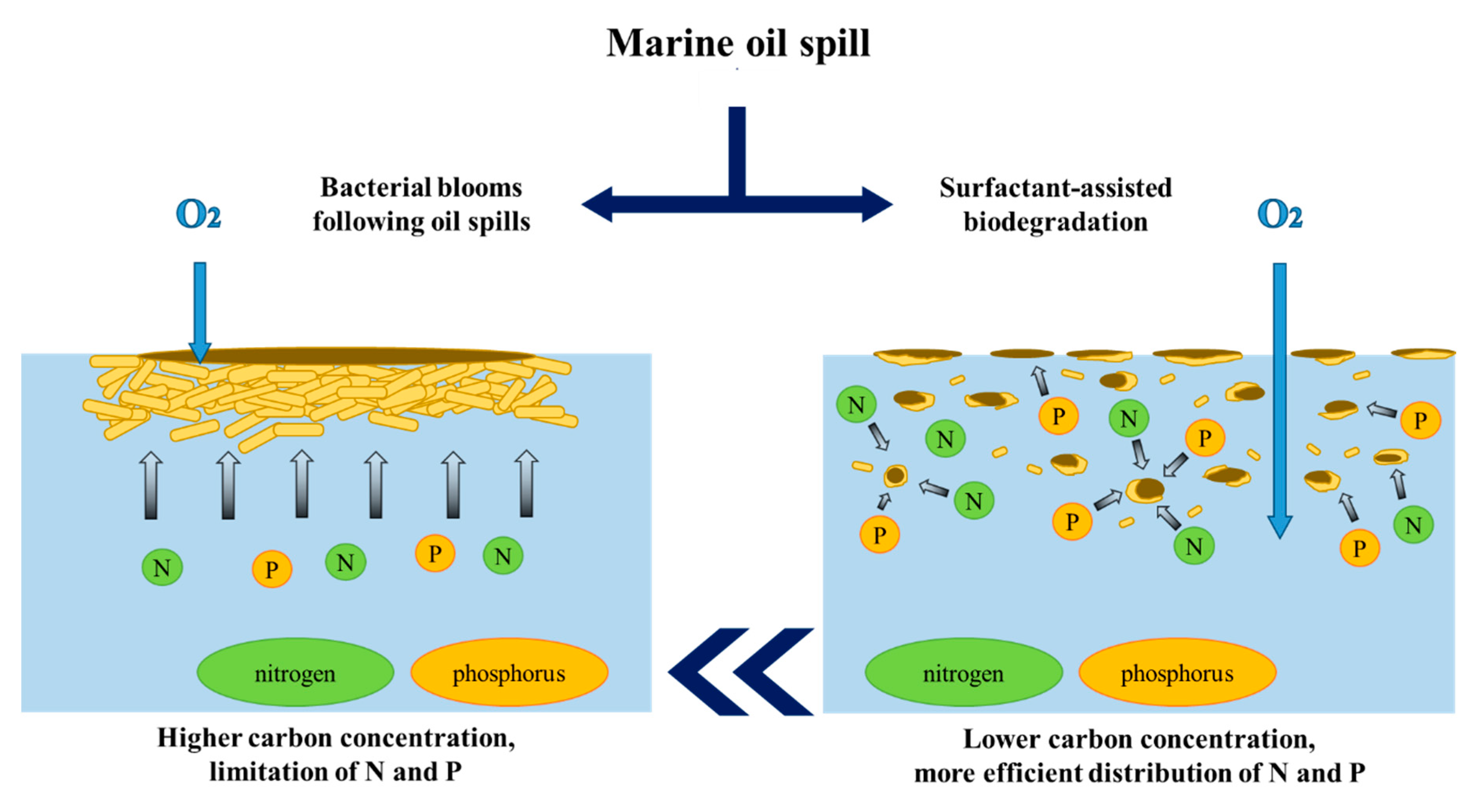
3. To Biodegrade or not to Biodegrade?
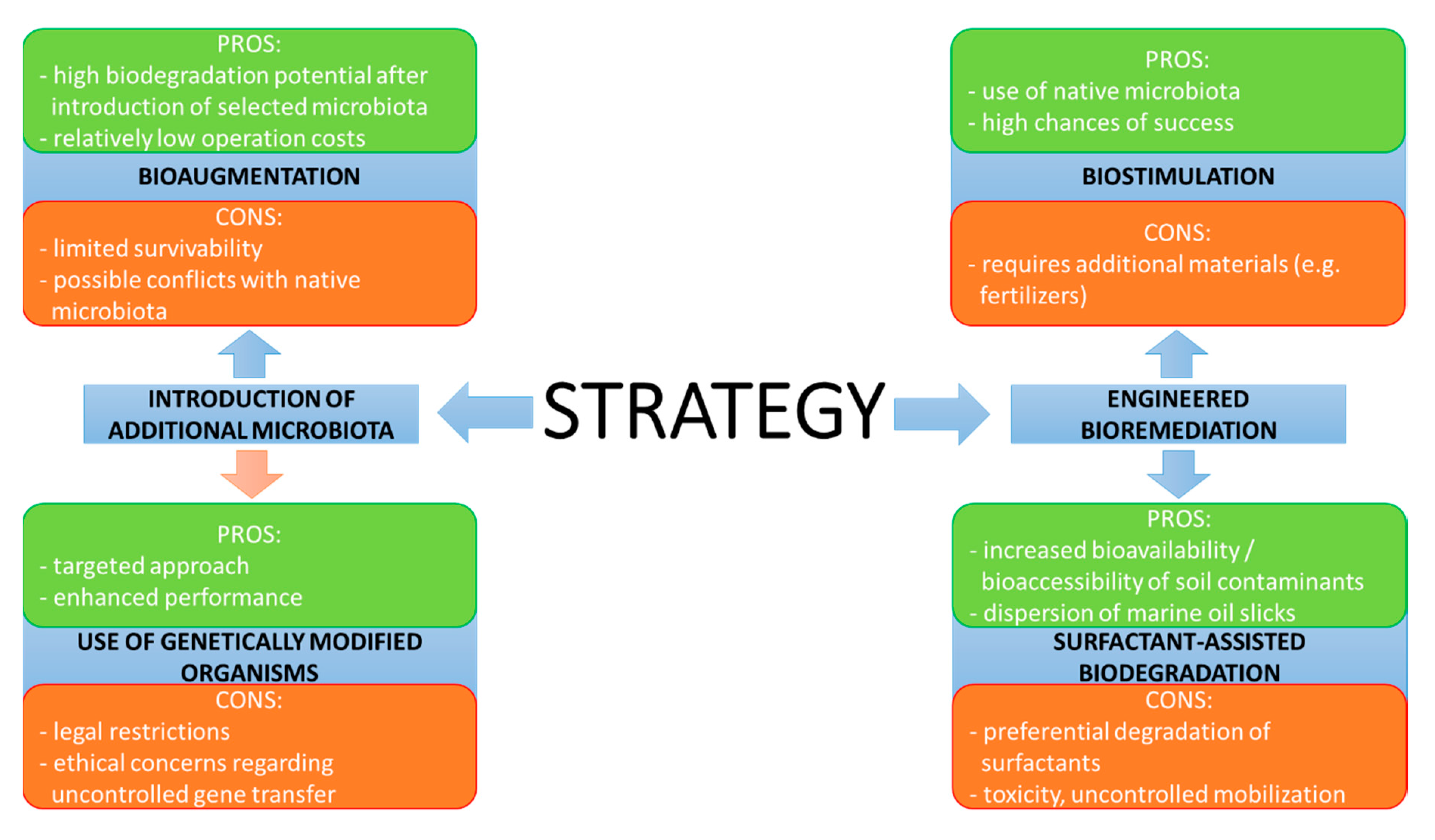
4. Practical Approaches to Bioremediation of Hydrocarbon Contaminations—The Good, the Bad and the Still Developing
5. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Heider, J.; Spormann, A.M.; Beller, H.R.; Widdel, F. Anaerobic bacterial metabolism of hydrocarbons. FEMS Microbiol. Rev. 1998, 22, 459–473. [Google Scholar] [CrossRef]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): A review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [Green Version]
- Farhadian, M.; Vachelard, C.; Duchez, D.; Larroche, C. In situ bioremediation of monoaromatic pollutants in groundwater: A review. Bioresour. Technol. 2008, 99, 5296–5308. [Google Scholar] [CrossRef]
- Chen, M.; Xu, P.; Zeng, G.; Yang, C.; Huang, D.; Zhang, J. Bioremediation of soils contaminated with polycyclic aromatic hydrocarbons, petroleum, pesticides, chlorophenols and heavy metals by composting: Applications, microbes and future research needs. Biotechnol. Adv. 2015, 33, 745–755. [Google Scholar] [CrossRef]
- Van Hamme, J.D.; Singh, A.; Ward, O.P. Recent advances in petroleum microbiology. Microbiol. Mol. Biol. Rev. 2003, 67, 503–549. [Google Scholar] [CrossRef] [Green Version]
- Dalrymple, G.B. The age of the Earth in the twentieth century: A problem (mostly) solved. Geol. Soc. Lond. Spec. Publ. 2001, 190, 205–221. [Google Scholar] [CrossRef]
- Maloof, A.C.; Porter, S.M.; Moore, J.L.; Dudas, F.O.; Bowring, S.A.; Higgins, J.A.; Fike, D.A.; Eddy, M.P. The earliest Cambrian record of animals and ocean geochemical change. Geol. Soc. Am. Bull. 2010, 122, 1731–1774. [Google Scholar] [CrossRef] [Green Version]
- Widdel, F.; Rabus, R. Anaerobic biodegradation of saturated and aromatic hydrocarbons. Curr. Opin. Biotechnol. 2001, 12, 259–276. [Google Scholar] [CrossRef]
- Cornelissen, G.; Gustafsson, Ö.; Bucheli, T.D.; Jonker, M.T.O.; Koelmans, A.A.; Van Noort, P.C.M. Extensive sorption of organic compounds to black carbon, coal, and kerogen in sediments and soils: Mechanisms and consequences for distribution, bioaccumulation, and biodegradation. Environ. Sci. Technol. 2005, 39, 6881–6895. [Google Scholar] [CrossRef]
- Schwark, L. Hydrocarbons in the Pedosphere. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Vandecasteele, J.-P. Petroleum Microbiology: Concepts, Environmental Implications, Industrial Applications; Editions Technip: Paris, France, 2008. [Google Scholar]
- Head, I.M.; Jones, D.M.; Larter, S.R. Biological activity in the deep subsurface and the origin of heavy oil. Nature 2003, 426, 344–352. [Google Scholar] [CrossRef]
- Head, I.M.; Jones, D.M.; Röling, W.F. Marine microorganisms make a meal of oil. Nat. Revi. Microbiol. 2006, 4, 173–182. [Google Scholar] [CrossRef]
- Meckenstock, R.U.; von Netzer, F.; Stumpp, C.; Lueders, T.; Himmelberg, A.M.; Hertkorn, N.; Schmitt-Kopplin, P.; Harir, M.; Hosein, R.; Haque, S.; et al. Oil biodegradation. Water droplets in oil are microhabitats for microbial life. Science 2014, 8, 673–676. [Google Scholar] [CrossRef]
- Wackett, L.P. Aliphatic Hydrocarbon Producers. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Capuano, E.; Oliviero, T.; Fogliano, V.; Pellegrini, N. Role of the food matrix and digestion on calculation of the actual energy content of food. Nutr. Rev. 2018, 76, 274–289. [Google Scholar] [CrossRef]
- Kharasch, M.S. Heats of combustion of organic compounds. J. Res. Bur Stand. 1929, 2, 359. [Google Scholar] [CrossRef]
- Sydow, M.; Owsianiak, M.; Szczepaniak, Z.; Framski, G.; Smets, B.F.; Ławniczak, Ł.; Lisiecki, P.; Szulc, A.; Cyplik, P.; Chrzanowski, Ł. Evaluating robustness of a diesel-degrading bacterial consortium isolated from contaminated soil. N. Biotechnol. 2016, 33, 852–859. [Google Scholar] [CrossRef] [Green Version]
- Ławniczak, Ł.; Marecik, R. Comparison of metalworking fluids biodegradation efficiency by autochthonous and environmental communities. J. Environ. Manag. 2019, 232, 625–635. [Google Scholar] [CrossRef]
- Baas-Becking, L.G.M. Geobiologie of Inleiding tot de Milieukunde; W.P. Van Stockum & Zoon: The Hague, The Netherlands, 1934. [Google Scholar]
- Kriszt, B.; Táncsics, A.; Cserháti, M.; Tóth, A.; Nagy, I.; Horváth, B.; Nagy, I.; Tamura, T.; Kukolya, J.; Szoboszlay, S. De novo genome project for the aromatic degrader Rhodococcus pyridinivorans strain AK37. J. Bacteriol. 2012, 194, 1247–1248. [Google Scholar] [CrossRef] [Green Version]
- Táncsics, A.; Benedek, T.; Szoboszlay, S.; Veres, P.G.; Farkas, M.; Máthé, I.; Márialigeti, K.; Kukolya, J.; Lányi, S.; Kriszt, B. The detection and phylogenetic analysis of the alkane 1-monooxygenase gene of members of the genus Rhodococcus. Syst. Appl. Microbiol. 2015, 38, 1–7. [Google Scholar] [CrossRef]
- Cases, I.; de Lorenzo, V. Promoters in the environment: Transcriptional regulation in its natural context. Nat. Rev. Microbiol. 2005, 3, 105–118. [Google Scholar] [CrossRef]
- Cases, I.; de Lorenzo, V. The black cat/white cat principle of signal integration in bacterial promoters. EMBO J. 2001, 20, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yakimov, M.M.; Timmis, K.N.; Golyshin, P.N. Obligate oil-degrading marine bacteria. Curr. Opin. Biotechnol. 2007, 18, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Owsianiak, M.; Szulc, A.; Chrzanowski, Ł.; Cyplik, P.; Bogacki, M.; Olejnik-Schmidt, A.K.; Heipieper, H.J. Biodegradation and surfactant-mediated biodegradation of diesel fuel by 218 microbial consortia are not correlated to cell surface hydrophobicity. Appl. Microbiol. Biotechnol. 2009, 84, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Benedek, T.; Táncsics, A.; Szilágyi, N.; Tóth, I.; Farkas, M.; Szoboszlay, S.; Krifaton, C.; Hartman, M.; Kriszt, B. Analysis of biofilm bacterial communities responsible for carbon removal through a reactor cascade treating wastewater. World J. Microbiol. Biotechnol. 2014, 30, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Benedek, T.; Szentgyörgyi, F.; Szabó, I.; Kriszt, B.; Révész, F.; Radó, J.; Maróti, G.; Táncsics, A. Aerobic and oxygen-limited enrichment of BTEX-degrading biofilm bacteria: Dominance of Malikia versus Acidovorax species. Environ. Sci. Pollut. Res. 2018, 25, 32178–32195. [Google Scholar] [CrossRef] [PubMed]
- Révész, F.; Figueroa-Gonzalez, P.A.; Probst, A.J.; Kriszt, B.; Banerjee, S.; Szoboszlay, S.; Maróti, G.; Táncsics, A. Microaerobic conditions caused the overwhelming dominance of Acinetobacter spp. and the marginalization of Rhodococcus spp. in diesel fuel/crude oil mixture-amended enrichment cultures. Arch. Microbiol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Szabó, I.; Szoboszlay, S.; Kriszt, B.; Háhn, J.; Harkai, P.; Baka, E.; Táncsics, A.; Kaszab, E.; Privler, Z.; Kukolya, J. Olivibacter oleidegradans sp. nov., a hydrocarbon degrading bacterium isolated from a biofilter cleanup facility on a hydrocarbon-contaminated site. Int. J. Syst. Evol. Microbiol. 2011, 61, 2861–2865. [Google Scholar] [CrossRef]
- Da Fonseca, M.M.B.; Minnicelli, C.F.; Silva-Portela, R.D.C.B.; de Farias, M.F.; dos Santos, P.R.S.; Fernandes, G.J.T.; Agnez-Lima, L.F. Unlocking and functional profiling of the bacterial communities in diesel tanks upon additive treatment. Fuel 2019, 236, 1311–1320. [Google Scholar] [CrossRef]
- Benedek, G.T.; Máthé, I.; Táncsics, A.; Márialigeti, K.; Albert, B.; Lányi, S. Intrinsic bioremediability of petroleum hydrocarbon contaminated sites in Romania: Diversity of bacterial community, catechol dioxygenase and alkane-monooxygenase. UPB Sci. Bull. B Chem. Mat. Sci. 2011, 73, 51–62. [Google Scholar]
- Navia, R.; Heipieper, H.J. The role of energy efficient biotechnological processes in the waste management industry. Waste Manag. Res. 2011, 29, 563–564. [Google Scholar] [CrossRef] [Green Version]
- Guerra, A.B.; Oliveira, J.S.; Silva-Portela, R.C.B.; Araújo, W.; Carlos, A.C.; Vasconcelos, A.T.R.; Freitas, A.T.; Domingos, Y.S.; de Farias, M.F.; Fernandes, G.J.T.; et al. Metagenome enrichment approach used for selection of oil-degrading bacteria consortia for drill cutting residue bioremediation. Environ. Pollut. 2018, 235, 869–880. [Google Scholar] [CrossRef]
- Li, J.; Luo, C.; Zhang, D.; Song, M.; Cai, X.; Jiang, L.; Zhang, G. Autochthonous bioaugmentation-modified bacterial diversity of phenanthrene degraders in PAH-contaminated wastewater as revealed by DNA-stable isotope probing. Environ. Sci. Technol. 2018, 52, 2934–2944. [Google Scholar] [CrossRef] [PubMed]
- Koolivand, A.; Abtahi, H.; Parhamfar, M.; Didehdar, M.; Saeedi, R.; Fahimirad, S. Biodegradation of high concentrations of petroleum compounds by using indigenous bacteria isolated from petroleum hydrocarbons-rich sludge: Effective scale-up from liquid medium to composting process. J. Environ. Manag. 2019, 248, 109228. [Google Scholar] [CrossRef] [PubMed]
- Pacwa-Płociniczak, M.; Czapla, J.; Płociniczak, T.; Piotrowska-Seget, Z. The effect of bioaugmentation of petroleum-contaminated soil with Rhodococcus erythropolis strains on removal of petroleum from soil. Ecotox. Environ. Saf. 2019, 169, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Woźniak-Karczewska, M.; Lisiecki, P.; Białas, W.; Owsianiak, M.; Piotrowska-Cyplik, A.; Wolko, Ł.; Ławniczak, Ł.; Heipieper, H.J.; Gutierrez, T.; Chrzanowski, Ł. Effect of bioaugmentation on long-term biodegradation of diesel/biodiesel blends in soil microcosms. Sci. Total Environ. 2019, 671, 948–958. [Google Scholar] [CrossRef]
- Wolf, D.C.; Gan, J. Influence of rhamnolipid biosurfactant and Brij-35 synthetic surfactant on 14C-pyrene mineralization in soil. Environ. Pollut. 2018, 243, 1846–1853. [Google Scholar] [CrossRef]
- Haleyur, N.; Shahsavari, E.; Jain, S.S.; Koshlaf, E.; Ravindran, V.B.; Morrison, P.D.; Osborn, A.M.; Ball, A.S. Influence of bioaugmentation and biostimulation on PAH degradation in aged contaminated soils: Response and dynamics of the bacterial community. J. Environ. Manag. 2019, 238, 49–58. [Google Scholar] [CrossRef]
- Tao, K.; Zhang, X.; Chen, X.; Liu, X.; Hu, X.; Yuan, X. Response of soil bacterial community to bioaugmentation with a plant residue-immobilized bacterial consortium for crude oil removal. Chemosphere 2019, 222, 831–838. [Google Scholar] [CrossRef]
- Wu, M.; Wu, J.; Zhang, X.; Ye, X. Effect of bioaugmentation and biostimulation on hydrocarbon degradation and microbial community composition in petroleum-contaminated loessal soil. Chemosphere 2019, 237, 124456. [Google Scholar] [CrossRef]
- Oualha, M.; Al-Kaabi, N.; Al-Ghouti, M.; Zouari, N. Identification and overcome of limitations of weathered oil hydrocarbons bioremediation by an adapted Bacillus sorensis strain. J. Environ. Manag. 2019, 250, 109455. [Google Scholar] [CrossRef]
- Ortega, M.F.; Guerrero, D.E.; García-Martínez, M.J.; Bolonio, D.; Llamas, J.F.; Canoira, L.; Gallego, J.L.R. Optimization of landfarming amendments based on soil texture and crude oil concentration. Water Air Soil Pollut. 2018, 229, 234. [Google Scholar] [CrossRef]
- Crampon, M.; Cébron, A.; Portet-Koltalo, F.; Uroz, S.; Le Derf, F.; Bodilis, J. Low effect of phenanthrene bioaccessibility on its biodegradation in diffusely contaminated soil. Environ. Pollut. 2017, 225, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zeng, G.; Zhong, H.; Wang, Z.; Liu, Z.; Cheng, M.; Liu, G.; Yang, X.; Liu, S. Effect of rhamnolipid solubilization on hexadecane bioavailability: Enhancement or reduction? J. Hazard. Mater. 2017, 322, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Lee, H.; Kwon, B.-O.; Khim, J.S.; Yim, U.H.; Kim, B.S.; Kim, J.-J. Biosurfactant-assisted bioremediation of crude oil by indigenous bacteria isolated from Taean beach sediment. Environ. Pollut. 2018, 241, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, J.; Dick, R.P.; Li, H.; Shen, D.; Gao, Y.; Waigi, M.G.; Ling, W. Rhamnolipid influences biosorption and biodegradation of phenanthrene by phenanthrene-degrading strain Pseudomonas sp. Ph6. Environ. Pollut. 2018, 240, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhang, J.; Peng, C. Shift of Soil Polycyclic Aromatic Hydrocarbons (PAHs) dissipation pattern and microbial community composition due to rhamnolipid supplementation. Water Air Soil Pollut. 2019, 230, 107. [Google Scholar] [CrossRef]
- Reddy, P.V.; Karegoudar, T.B.; Nayak, A.S. Enhanced utilization of fluorene by Paenibacillus sp. PRNK-6: Effect of rhamnolipid biosurfactant and synthetic surfactants. Ecotox. Environ. Saf. 2018, 151, 206–211. [Google Scholar] [CrossRef]
- Staninska-Pięta, J.; Piotrowska-Cyplik, A.; Juzwa, W.; Zgoła-Grześkowiak, A.; Wolko, Ł.; Sydow, Z.; Kaczorowski, Ł.; Powierska-Czarny, J.; Cyplik, P. The impact of natural and synthetic surfactants on bacterial community during hydrocarbon biodegradation. Int. Biodeterior. Biodegrad. 2019, 142, 191–199. [Google Scholar] [CrossRef]
- Czarny, J.; Staninska-Pięta, J.; Piotrowska-Cyplik, A.; Wolko, Ł.; Staninski, K.; Hornik, B.; Cyplik, P. Assessment of soil potential to natural attenuation and autochthonous bioaugmentation using microarray and functional predictions from metagenome profiling. Ann. Microbiol. 2019, 69, 945–955. [Google Scholar] [CrossRef]
- Ramadass, K.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Bioavailability of weathered hydrocarbons in engine oil-contaminated soil: Impact of bioaugmentation mediated by Pseudomonas spp. on bioremediation. Sci. Total Environ. 2018, 636, 968–974. [Google Scholar] [CrossRef]
- Roy, A.; Dutta, A.; Pal, S.; Gupta, A.; Sarkar, J.; Chatterjee, A.; Saha, A.; Sarkar, P.; Sar, P.; Kazy, S.K. Biostimulation and bioaugmentation of native microbial community accelerated bioremediation of oil refinery sludge. Bioresour. Technol. 2018, 253, 22–32. [Google Scholar] [CrossRef]
- Safdari, M.-S.; Kariminia, H.-R.; Rahmati, M.; Fazlollahi, F.; Polasko, A.; Mahendra, S.; Wilding, W.V.; Fletcher, T.H. Development of bioreactors for comparative study of natural attenuation, biostimulation, and bioaugmentation of petroleum-hydrocarbon contaminated soil. J. Hazard. Mater. 2018, 342, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.; Upasani, V.N. Influence of abiotic factors, natural attenuation, bioaugmentation and nutrient supplementation on bioremediation of petroleum crude contaminated agricultural soil. J. Environ. Manag. 2019, 245, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Cecotti, M.; Coppotelli, B.M.; Mora, V.C.; Viera, M.; Morelli, I.S. Efficiency of surfactant-enhanced bioremediation of aged polycyclic aromatic hydrocarbon-contaminated soil: Link with bioavailability and the dynamics of the bacterial community. Sci. Total Environ. 2018, 634, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Thompson, I.P.; Van Der Gast, C.J.; Ciric, L.; Singer, A.C. Bioaugmentation for bioremediation: The challenge of strain selection. Environ. Microbiol. 2005, 7, 909–915. [Google Scholar] [CrossRef]
- Atashgahi, S.; Sánchez-Andrea, I.; Heipieper, H.J.; van der Meer, J.R.; Stams, A.J.M.; Smidt, H. Prospects for harnessing biocide resistance for bioremediation and detoxification. Science 2018, 360, 743–746. [Google Scholar] [CrossRef] [Green Version]
- Bradford, L.M.; Vestergaard, G.; Táncsics, A.; Zhu, B.; Schloter, M.; Lueders, T. Transcriptome-Stable isotope probing provides targeted functional and taxonomic insights into microaerobic pollutant-degrading aquifer microbiota. Front. Microbiol. 2018, 9, 02696. [Google Scholar] [CrossRef]
- Semple, K.T.; Doick, K.J.; Jones, K.C.; Burauel, P.; Craven, A.; Harms, H. Defining bioavailability and bioaccessibility of contaminated soil and sediment is complicated. Environ. Sci. Technol. 2004, 38, 228A–231A. [Google Scholar] [CrossRef] [Green Version]
- Perminova, I.V.; Grechishcheva, N.Y.; Kovalevskii, D.V.; Kudryavtsev, A.V.; Petrosyan, V.S.; Matorin, D.N. Quantification and prediction of the detoxifying properties of humic substances related to their chemical binding to polycyclic aromatic hydrocarbons. Environ. Sci. Technol. 2001, 35, 3841–3848. [Google Scholar] [CrossRef]
- Paria, S. Surfactant-enhanced remediation of organic contaminated soil and water. Adv. Colloid Interface Sci. 2008, 138, 24–58. [Google Scholar] [CrossRef]
- Wang, X.; Sun, L.; Wang, H.; Wu, H.; Chen, S.; Zheng, X. Surfactant-enhanced bioremediation of DDTs and PAHs in contaminated farmland soil. Environ. Technol. 2018, 39, 1733–1744. [Google Scholar] [CrossRef]
- Peziak, D.; Piotrowska, A.; Marecik, R.; Lisiecki, P.; Woźniak, M.; Szulc, A.; Ławniczak, Ł.; Chrzanowski, Ł. Bioavailability of hydrocarbons to bacterial consortia during Triton X-100 mediated biodegradation in aqueous media. Acta Bioch. Pol. 2013, 60, 789–793. [Google Scholar] [CrossRef] [Green Version]
- Cierniak, D.; Woźniak-Karczewska, M.; Parus, A.; Wyrwas, B.; Loibner, A.P.; Heipieper, H.J.; Ławniczak, Ł.; Chrzanowski, Ł. How to accurately assess surfactant biodegradation-impact of sorption on the validity of results. Appl. Microbiol. Biotechnol. 2020, 104, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marecik, R.; Wojtera-Kwiczor, J.; Ławniczak, Ł.; Cyplik, P.; Szulc, A.; Piotrowska-Cyplik, A.; Chrzanowski, Ł. Rhamnolipids increase the phytotoxicity of diesel oil towards four common plant species in a terrestrial environment. Water Air Soil Pollut. 2012, 223, 4275–4282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzionek, A.; Wojcieszyńska, D.; Hupert-Kocurek, K.; Adamczyk-Habrajska, M.; Guzik, U. Immobilization of Planococcus sp. S5 strain on the loofah sponge and its application in naproxen removal. Catalysts 2018, 8, 176. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioremediation Strategy | Contaminants | Test System | Removal Efficiency | Process Duration | Conclusions/Comments | Reference |
|---|---|---|---|---|---|---|
| Autochthonous bioaugmentation | Alkanes(initial concentration not specified) | Drill cuttings microcosms | 35–66% | 12 weeks | Consortia were isolated from drill cuttings, enriched and reintroduced. The consortia exhibited a high biodegradation potential towards several hydrocarbon substrates and the ability to produce biosurfactants. Enrichment of Firmicutes was observed. | Guerra et al. 2018 [34] |
| Autochthonous bioaugmentation | Phenanthrene (10 mg/L) | Bottle tests | >90% | 6 days | Autochthonous bioaugmentation allowed to improve the biodegradation efficiency. The re-introduced autochthonous isolate did not directly participate in the biodegradation process, and the improvement was attributed to altered diversity of PAH degraders. | Li et al. 2018 [35] |
| Autochthonous bioaugmentation | Crude oil (10–50 g/kg) | Composting | 60–91% | 12 weeks | Re-introduction of two autochthonous isolates into the population allowed for successful bioaugmentation and improvement of the biodegradation process. | Koolivand et al. 2019 [36] |
| Bioaugmentation | Crude oil (TPH at 12 g/kg) | Soil microcosms | 30–38% | 182 days | The bioaugmentation initially improved the biodegradation efficiency; however, after 91 days, a significant decrease of soil respiration was observed with changes of the bacterial community composition. | Pacwa-Płociniczak et al. 2019 [37] |
| Bioaugmentation | Diesel oil and diesel/biodiesel blends (1% v/w) | Soil microcosms | 88–97% | 64.5 weeks | Bioaugmentation initially improved the biodegradation kinetics; however, there was no significant effect in the long term. Furthermore, the ratio of aliphatic to aromatic fractions remained unchanged regardless of the treatment used. | Woźniak-Karczewska et al. 2019 [38] |
| Bioaugmentation + biosurfactant/surfactant-assisted biodegradation | Pyrene (10 mg/kg) | Soil microcosms | 60% | 10 days | Bioaugmentation was successful. High biodegradation efficiency was observed in the case of unamended and surfactant (Brij-35)-amended soil samples. Supplementation with rhamnolipids inhibited the biodegradation process due to their utilization as a preferential carbon source. | Wolf and Gan 2018 [39] |
| Bioaugmentation + biostimulation | PAHs (1.5 g/kg) | Soil mesocosms | 99% | 56 days | Biostimulation successfully improved the biodegradation efficiency, whereas bioaugmentation did not significantly contribute to the process. Enrichment of the community in PAH-degrading species was observed. | Haleyur et al. 2019 [40] |
| Bioaugmentation + biostimulation | Crude oil (TPH at 20 g/kg) | Soil microcosms | 36–51% | 30 days | The highest biodegradation efficiency was achieved when bioaugmentation was carried out using an immobilized bacterial consortium, with Eichhornia crassipes dried straw acting as both a carried and additional source of C and N. | Tao et al. 2019 [41] |
| Bioaugmentation + biostimulation | Crude oil (TPH at 19.8 g/kg) | Soil microcosms | 28% (biostimulation) and 14% (bioaugmentation) | 12 weeks | Biostimulation allowed to achieve superior efficiency compared to bioaugmentation. Application of bioaugmentation resulted in notably decreased biodiversity of the soil community. | Wu et al. 2019 [42] |
| Bioaugmentation + biostimulation + surfactant-assisted biodegradation | Diesel oil hydrocarbons (3 g/kg) + PAHs (400 µg/kg) | Weathered oily-soil biopiles | 39% for diesel oil hydrocarbons and 32% for PAHs | 160 days | Combined bioaugmentation, biostimulation and surfactant supplementation (Tween 80) improved the biodegradation efficiency. In case of biostimulation, ammonium nitrate facilitated the process, whereas the use of urea inhibited the biodegradation efficiency. | Oualha et al. 2019 [43] |
| Biostimulation + surfactant-assisted biodegradation | Crude oil (either 20 g/kg or 50 g/kg) | Field study in soil | 49–62% | 486 days | Biostimulation improved the biodegradation efficiency in all experimental variants. Surfactant supplementation (Bioversal) improved the biodegradation process in cases of higher concentration of crude oil, whereas in cases of lower concentrations, it did not significantly affect the process. | Ortega et al. 2018 [44] |
| Biosurfactant-assisted biodegradation | Phenanthrene (0.1–1.0 mg/L) | Sorption reactors with soil | >90% | Up to 50 days | Supplementation of the biosurfactant (rhamnolipids) influenced the sorption kinetics of phenanthrene; however, it had no effect on its biodegradation kinetics. No significant influence of the biosurfactant on the main phenanthrene degraders was observed. | Crampon et al. 2017 [45] |
| Biosurfactant-assisted biodegradation | Hexadecane (2% v/v) | Flask studies | 20–100% | 180 h | The biosurfactant (rhamnolipids) increased the availability of hexadecane in the case of Pseudomonas aeruginosa (which was capable of producing rhamnolipids) and decreased the availability in the case of Pseudomonas putida (which was unable to produce rhamnolipids). The decrease occurred due to a blocking effect by rhamnolipids. Dissipation of rhamnolipids was also observed. | Liu et al. 2017 [46] |
| Biosurfactant-assisted biodegradation | Crude oil (1% v/v) | Flask studies | >85% | 14 days | Isolates from beach sediments exhibited the ability to efficiently degrade hydrocarbons and produce biosurfactants. The biosurfactants increased the emulsification of crude oil and facilitated the biodegradation process. | Lee et al. 2018 [47] |
| Biosurfactant-assisted biodegradation | Phenanthrene (0.2–1.0 mg/L) | Flask studies | 60–100% | 14 days | The biosurfactant (rhamnolipids) was supplemented in order to improve the biodegradation efficiency. At a concentration of up to 100 mg/L of rhamnolipids, an enhancement of phenanthrene biodegradation was observed. At concentrations higher than 200 mg/L of rhamnolipids, the biodegradation efficiency was decreased due to the hindered biosorption of phenanthrene. | Ma et al. 2018 [48] |
| Biosurfactant-assisted biodegradation | PAHs: phenanthrene, fluoranthene, and pyrene (6 mg/kg) | Soil microcosms | 72% for phenanthrene, 64% for fluoranthene, and 58% for pyrene at day 7 | up to 35 days | Supplementation with the biosurfactant (rhamnolipids) initially increased the biodegradation of the studied PAHs (at day 7); however, no effect or even lower efficiency were observed in the latter stages (up to 35 days). | Lu et al. 2019 [49] |
| Biosurfactant/surfactant-assisted biodegradation | Fluorene (280 or 320 mg/L) | Flask studies | 75–97% | 24 h | Supplementation with the biosurfactant (rhamnolipids) allowed to achieve higher biodegradation efficiency compared to synthetic surfactants (Tween-80, Tween-60, Tween-40, Tween-20 and Triton X-100). | Reddy et al. 2018 [50] |
| Biosurfactant/surfactant-assisted biodegradation | Diesel oil (1% v/v) | Flaks studies | 20–99% | 7 days | Surfactant supplementation (Tween-80) enhanced the biodegradation of diesel oil hydrocarbons. Supplementation with the biosurfactant (rhamnolipids) inhibited the biodegradation process due to their utilization as a preferential substrate. | Staninska-Pięta et al. 2019 [51] |
| Natural attenuation + autochthonous bioaugmentation | Diesel oil (1% v/v) | Flask tests | 20–40% | 7 days | Efficiency of biodegradation processes with autochthonous bioaugmentation depended on the previous exposure of soils to pollution. In the majority of tested systems, the autochthonous bioaugmentation resulted in a significant enrichment of Proteobacteria. | Czarny et al. 2019 [52] |
| Natural attenuation + bioaugmentation + biostimulation | Engine oil (39–41 g/kg TPH) | Soil microcosms | 31–75% | 210 days | The combined bioaugmentation and biostimulation approach resulted in the inhibition of biodegradation processes in comparison to natural attenuation. | Ramadass et al. 2018 [53] |
| Natural attenuation + bioaugmentation + biostimulation | Petroleum refinery waste (TPH at 144 g/kg) | Vial microcosms | 57–75% | 120 days | Combined bioaugmentation-biostimulation approach allowed to achieve the best biodegradation efficiency. Biostimulation was the major driving force for the enhancement. | Roy et al. 2018 [54] |
| Natural attenuation + bioaugmentation + biostimulation | Crude oil (20 g/kg) | Bioreactors with soil | 51–90% | 60 days | The combined bioaugmentation and biostimulation approach allowed to achieve the highest biodegradation rate. Among individual treatments, the efficiency of biostimulation was superior (82% of TPH removal) compared to bioaugmentation (63% of TPH removal). | Safdari et al. 2018 [55] |
| Natural attenuation + bioaugmentation + biostimulation | Crude oil (3% w/v) | Soil microcosms | 94% | 45 days | Combined bioaugmentation and biostimulation allowed to achieve the most rapid and efficient biodegradation process. | Varjani and Upasani 2019 [56] |
| Surfactant-assisted biodegradation | PAHs (574 mg/kg) | Soil microcosms | 72–77% | 84 days | Enhanced biodegradation was observed at sub-CMC concentrations of the surfactant (Triton X-100), whereas decreased efficiency was observed at CMC concentrations. The negative effect may be attributed to the preferential degradation of surfactant at CMC concentrations. | Cecotti et al. 2018 [57] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ławniczak, Ł.; Woźniak-Karczewska, M.; Loibner, A.P.; Heipieper, H.J.; Chrzanowski, Ł. Microbial Degradation of Hydrocarbons—Basic Principles for Bioremediation: A Review. Molecules 2020, 25, 856. https://doi.org/10.3390/molecules25040856
Ławniczak Ł, Woźniak-Karczewska M, Loibner AP, Heipieper HJ, Chrzanowski Ł. Microbial Degradation of Hydrocarbons—Basic Principles for Bioremediation: A Review. Molecules. 2020; 25(4):856. https://doi.org/10.3390/molecules25040856
Chicago/Turabian StyleŁawniczak, Łukasz, Marta Woźniak-Karczewska, Andreas P. Loibner, Hermann J. Heipieper, and Łukasz Chrzanowski. 2020. "Microbial Degradation of Hydrocarbons—Basic Principles for Bioremediation: A Review" Molecules 25, no. 4: 856. https://doi.org/10.3390/molecules25040856
APA StyleŁawniczak, Ł., Woźniak-Karczewska, M., Loibner, A. P., Heipieper, H. J., & Chrzanowski, Ł. (2020). Microbial Degradation of Hydrocarbons—Basic Principles for Bioremediation: A Review. Molecules, 25(4), 856. https://doi.org/10.3390/molecules25040856