Protective Effect of Diphlorethohydroxycarmalol Isolated from Ishige okamurae Against Particulate Matter-Induced Skin Damage by Regulation of NF-κB, AP-1, and MAPKs Signaling Pathways In Vitro in Human Dermal Fibroblasts

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
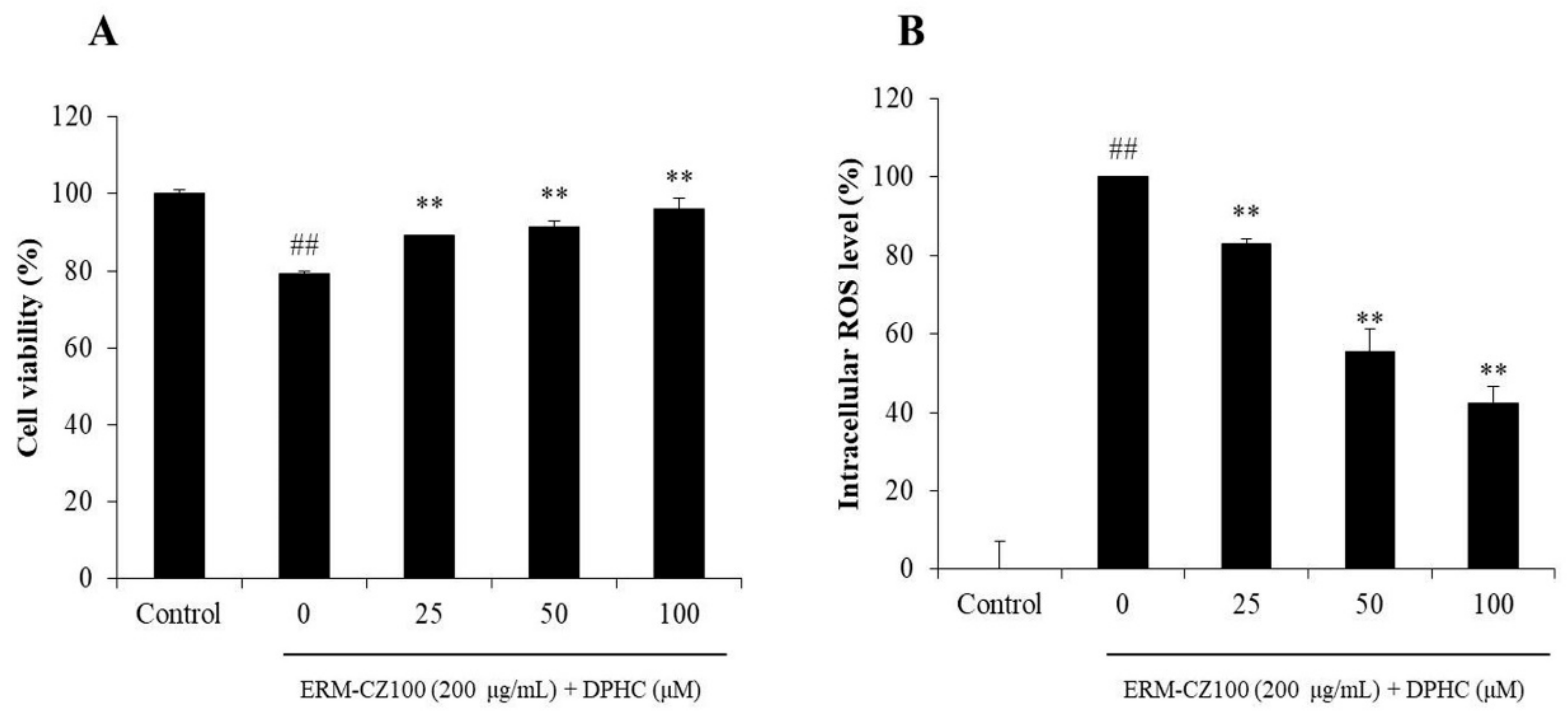
2.1. DPHC Improves PM-Induced Oxidative Damage in HDF Cells
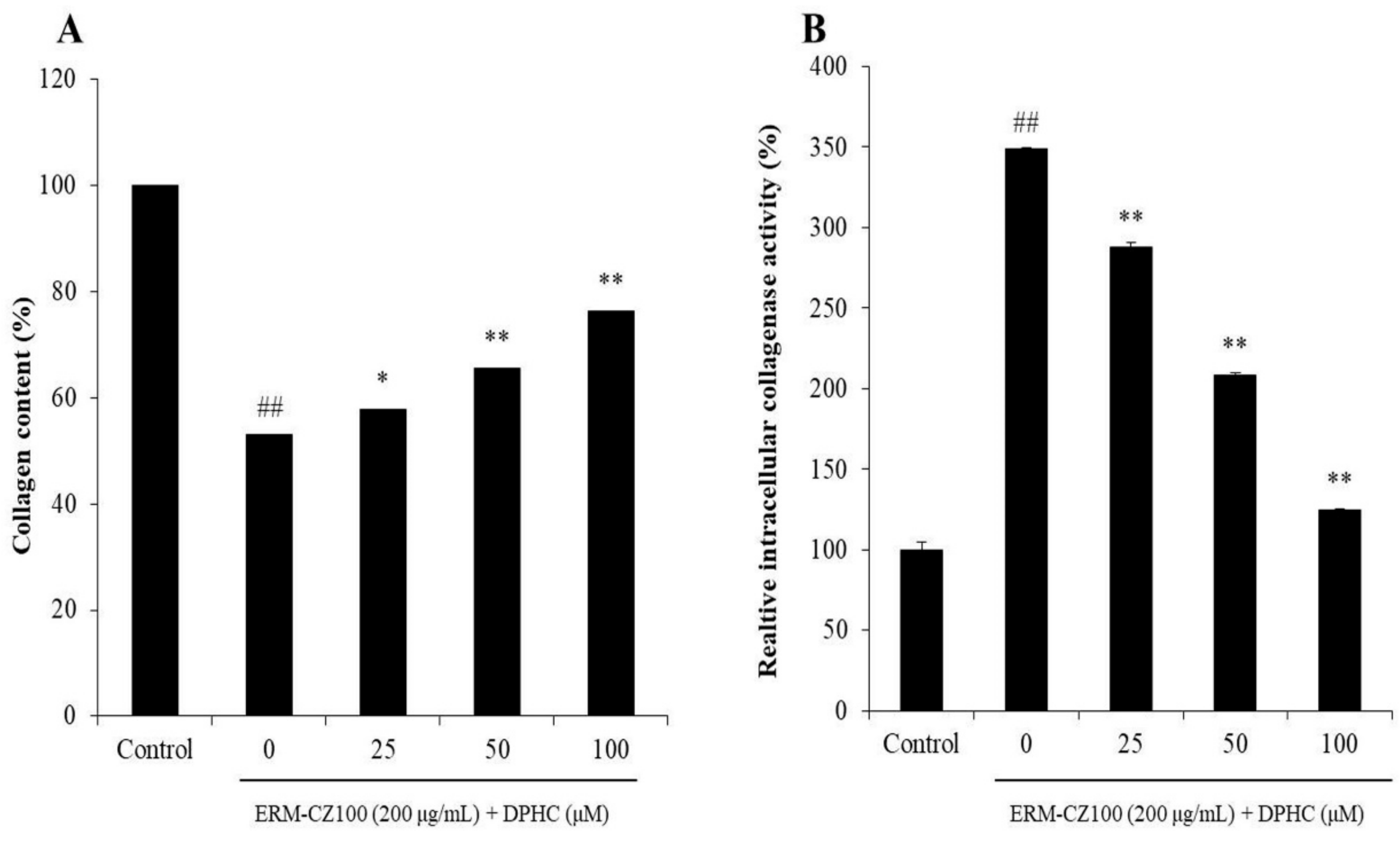
2.2. DPHC Protects Collagen Synthesis and Inhibits Intracellular Collagenase Activity in PM-Stimulated HDF Cells
2.3. DPHC Reduces Matrix Metalloproteinases (MMPs) Expression in PM-Stimulated HDF Cells
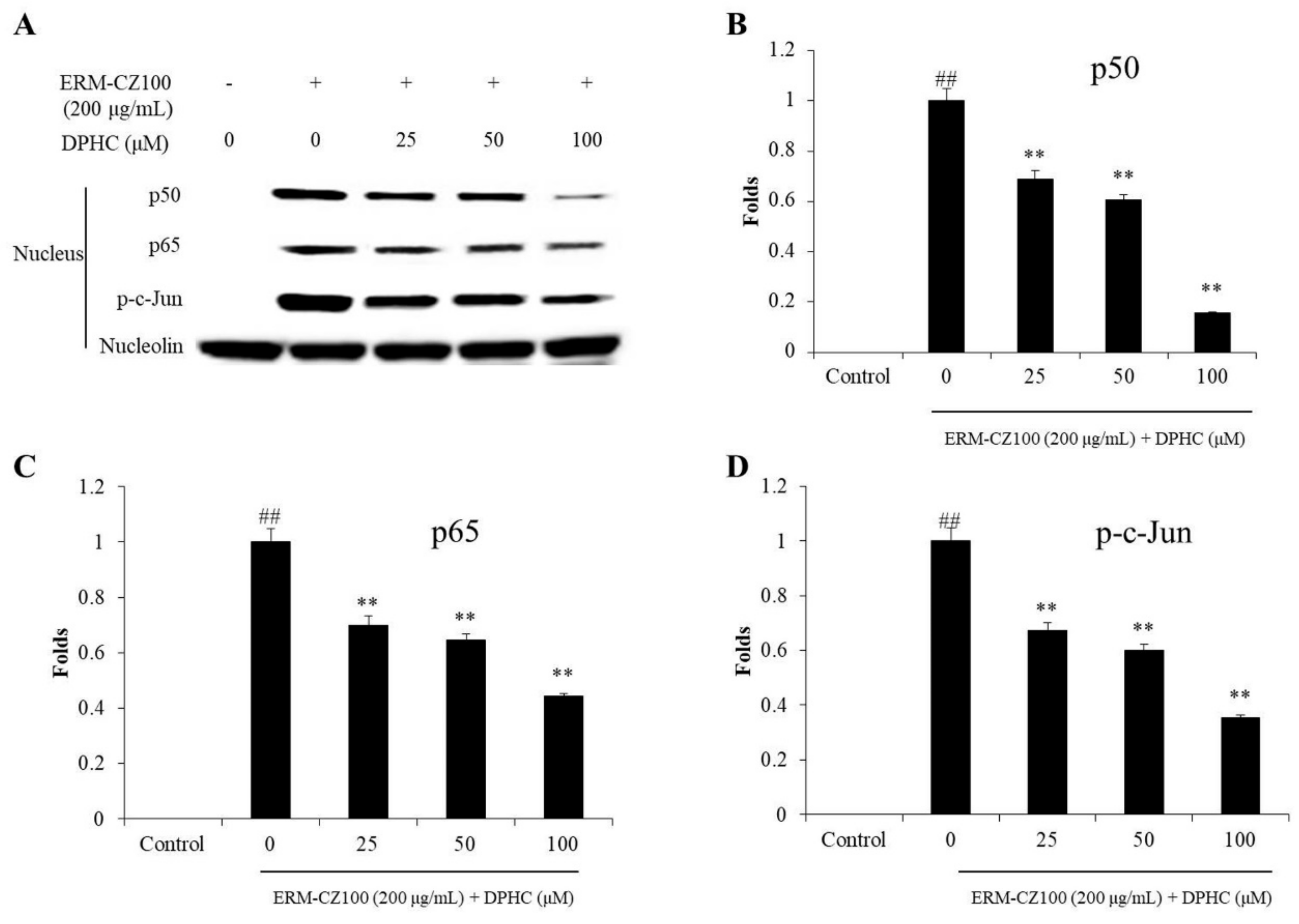
2.4. DPHC Inhibits Nuclear Factor Kappa B (NF-κB) Activation, Reduces Activator Protein (AP-1) Phosphorylation, and Suppresses Mitogen-Activated Protein Kinases (MAPKs) Activation in PM-Stimulated HDF Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of DPHC
4.3. Cell Culture
4.4. Determination of the Effect of DPHC on PM-Induced Cytotoxicity in HDF Cells
4.5. Determination of the Effect of DPHC on Collagen Synthesis, MMP Expression, and Intracellular Collagenase Activity
4.6. Determination of the Effect of DPHC on NF-κB, AP-1, and MAPKs in PM-Stimulated HDF Cells
Author Contributions
Funding
Conflicts of Interest
References
- Burnett, R.; Chen, H.; Szyszkowicz, M.; Fann, N.; Hubbell, B.; Pope, C.A.; Apte, J.S.; Brauer, M.; Cohen, A.; Weichenthal, S.; et al. Global estimates of mortality associated with long-term exposure to outdoor fine particulate matter. Proc. Natl. Acad. Sci. USA 2018, 115, 9592–9597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Ouyang, W.; Shu, Y.; Tian, Y.; Feng, Y.; Zhang, T.; Chen, W. Incorporating bioaccessibility into health risk assessment of heavy metals in particulate matter originated from different sources of atmospheric pollution. Environ. Pollut. 2019, 254, 113113. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-C.; Schwartz, J. Metabolic Syndrome and Inflammatory Responses to Long-Term Particulate Air Pollutants. Environ. Heal. Perspect. 2008, 116, 612–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubowsky, S.D.; Suh, H.H.; Schwartz, J.; Coull, B.A.; Gold, D.R. Diabetes, Obesity, and Hypertension May Enhance Associations between Air Pollution and Markers of Systemic Inflammation. Environ. Heal. Perspect. 2006, 114, 992–998. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Kim, H.-S.; Sanjeewa, K.K.A.; Oh, J.-Y.; Jeon, Y.-J.; Lee, W.W. Inhibition of inflammatory responses elicited by urban fine dust particles in keratinocytes and macrophages by diphlorethohydroxycarmalol isolated from a brown alga Ishige okamurae. ALGAE 2017, 32, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, S.-Y.; Lee, H.G.; Je, J.-G.; Jee, Y.; Jeon, Y.-J. Sargassum horneri (Turner) inhibit urban particulate matter-induced inflammation in MH-S lung macrophages via blocking TLRs mediated NF-κB and MAPK activation. J. Ethnopharmacol. 2019, 249, 112363. [Google Scholar] [CrossRef]
- Bowe, B.; Xie, Y.; Li, T.; Yan, Y.; Xian, H.; Al-Aly, Z. Particulate Matter Air Pollution and the Risk of Incident CKD and Progression to ESRD. J. Am. Soc. Nephrol. 2017, 29, 218–230. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Lee, W.; Cui, Y.R.; Ahn, G.; Jeon, Y.-J. Protective effect of green tea catechin against urban fine dust particle-induced skin aging by regulation of NF-κB, AP-1, and MAPKs signaling pathways. Environ. Pollut. 2019, 252, 1318–1324. [Google Scholar] [CrossRef]
- Piao, M.J.; Ahn, M.J.; Kang, K.A.; Ryu, Y.S.; Hyun, Y.J.; Shilnikova, K.; Zhen, A.X.; Jeong, J.W.; Choi, Y.H.; Kang, H.K.; et al. Particulate matter 2.5 damages skin cells by inducing oxidative stress, subcellular organelle dysfunction, and apoptosis. Arch. Toxicol. 2018, 92, 2077–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Wang, P.; Pan, X.; Wang, Y.; Ren, S. Effects of short-term exposure to particulate matters on heart rate variability: A systematic review and meta-analysis based on controlled animal studies. Environ. Pollut. 2019, 256, 113306. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Hann, T.; Lee, S.J. Effect of flow and humidity on indoor deposition of particulate matter. Environ. Pollut. 2019, 255, 113263. [Google Scholar] [CrossRef] [PubMed]
- Oh, I.; Lee, J.; Ahn, K.; Kim, J.; Kim, Y.-M.; Sim, C.S.; Kim, Y. Association between particulate matter concentration and symptoms of atopic dermatitis in children living in an industrial urban area of South Korea. Environ. Res. 2018, 160, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Zhang, Q. Oxidative damage of dust storm fine particles instillation on lungs, hearts and livers of rats. Environ. Toxicol. Pharmacol. 2006, 22, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Moretti, S.; Smets, W.; Hofman, J.; Mubiana, K.V.; Oerlemans, E.; Vandenheuvel, D.; Samson, R.; Blust, R.; Lebeer, S. Human inflammatory response of endotoxin affected by particulate matter-bound transition metals. Environ. Pollut. 2019, 244, 118–126. [Google Scholar] [CrossRef]
- White, A.; Keller, J.; Zhao, S.; Kaufman, J.; Sandler, D. Air Pollution, Clustering of Particulate Matter Components and Breast Cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 624–625. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ruan, Z.; Wang, X.; Yang, Y.; Mason, T.G.; Lin, H.; Tian, L. Short-term and long-term exposures to fine particulate matter constituents and health: A systematic review and meta-analysis. Environ. Pollut. 2019, 247, 874–882. [Google Scholar] [CrossRef]
- Kim, H.-S.; Shin, B.-O.; Kim, S.; Wang, L.; Lee, W.; Kim, Y.T.; Rho, S.; Cho, M.; Jeon, Y.-J. Antioxidant Activity of Pepsin Hydrolysate Derived from Edible Hippocampus abdominalis in vitro and in Zebrafish Models. Korean J. Fish. Aquat. Sci. 2016, 49, 445–453. [Google Scholar]
- Wang, L.; Oh, J.Y.; Kim, H.S.; Lee, W.; Cui, Y.; Lee, H.G.; Kim, Y.-T.; Jeon, Y.-J.; Ko, J.Y. Protective effect of polysaccharides from Celluclast-assisted extract of Hizikia fusiforme against hydrogen peroxide-induced oxidative stress in vitro in Vero cells and in vivo in zebrafish. Int. J. Boil. Macromol. 2018, 112, 483–489. [Google Scholar] [CrossRef]
- Wang, L.; Park, Y.-J.; Jeon, Y.-J.; Ryu, B. Bioactivities of the edible brown seaweed, Undaria pinnatifida: A review. Aquaculture 2018, 495, 873–880. [Google Scholar] [CrossRef]
- Wright, A.D.; König, G.M.; Angerhofer, C.K.; Greenidge, P.; Linden, A.; Desqueyroux-Faúndez, R. Antimalarial Activity: The Search for Marine-Derived Natural Products with Selective Antimalarial Activity. J. Nat. Prod. 1996, 59, 710–716. [Google Scholar] [CrossRef]
- Suzuki, M.; Kawamoto, T.; Vairappan, C.S.; Ishii, T.; Abe, T.; Masuda, M. Halogenated metabolites from Japanese Laurencia spp. Phytochemistry 2005, 66, 2787–2793. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, W.A.J.P.; Kim, E.-A.; Kang, M.-C.; Lee, W.-W.; Lee, H.-S.; Vairappan, C.S.; Jeon, Y.-J. Assessment of anti-inflammatory effect of 5β-hydroxypalisadin B isolated from red seaweed Laurencia snackeyi in zebrafish embryo in vivo model. Environ. Toxicol. Pharmacol. 2014, 37, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Vairappan, C.S. Antioxidant, antibacterial and cytotoxic activities of essential oils and ethanol extracts of selected South East Asian herbs. J. Med. Plants Res. 2011, 5, 5284–5290. [Google Scholar]
- Kamada, T.; Vairappan, C.S. A New Bromoallene-Producing Chemical Type of the Red Alga Laurencia nangii Masuda. Molecules 2012, 17, 2119–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagappan, T.; Segaran, T.C.; Wahid, M.E.A.; Ramasamy, P.; Vairappan, C.S. Efficacy of Carbazole Alkaloids, Essential Oil and Extract of Murraya koenigii in Enhancing Subcutaneous Wound Healing in Rats. Molecules 2012, 17, 14449–14463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.-Y.; Kang, N.; Lee, J.-H.; Kim, J.-S.; Kim, W.-S.; Park, S.-J.; Kim, Y.-T.; Jeon, Y.-J. Angiotensin I-converting enzyme inhibitory peptides from an enzymatic hydrolysate of flounder fish (Paralichthys olivaceus) muscle as a potent anti-hypertensive agent. Process. Biochem. 2016, 51, 535–541. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.; Hwang, J.; Ko, J.; Jeon, Y.-J.; Ryu, B. In Vitro and In Vivo Antioxidant Activities of Polysaccharides Isolated from Celluclast-Assisted Extract of an Edible Brown Seaweed, Sargassum fulvellum. Antioxidants 2019, 8, 493. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.; Lin, X.; Wang, P.; Zhou, X.; Yang, B.; Wang, J.; Liu, Y. Perylenequione Derivatives with Anticancer Activities Isolated from the Marine Sponge-Derived Fungus, Alternaria sp. SCSIO41014. Mar. Drugs 2018, 16, 280. [Google Scholar] [CrossRef] [Green Version]
- Sanjeewa, K.K.A.; Lee, W.W.; Kim, J.-I.; Jeon, Y.-J. Exploiting biological activities of brown seaweed Ishige okamurae Yendo for potential industrial applications: A review. Environ. Boil. Fishes 2017, 29, 3109–3119. [Google Scholar] [CrossRef]
- Ryu, B.; Jiang, Y.; Kim, H.-S.; Hyun, J.-M.; Lim, S.-B.; Li, Y.; Jeon, Y.-J. Ishophloroglucin A, a Novel Phlorotannin for Standardizing the Anti-α-Glucosidase Activity of Ishige okamurae. Mar. Drugs 2018, 16, 436. [Google Scholar] [CrossRef] [Green Version]
- Yoon, N.Y.; Lee, S.-H.; Li, Y.; Kim, S.-K. Phlorotannins from Ishige okamurae and their acetyl- and butyrylcholinesterase inhibitory effects. J. Funct. Foods 2009, 1, 331–335. [Google Scholar] [CrossRef]
- Zou, Y.; Qian, Z.-J.; Li, Y.; Kim, M.-M.; Lee, S.-H.; Kim, S.-K. Antioxidant Effects of Phlorotannins Isolated from Ishige okamurae in Free Radical Mediated Oxidative Systems. J. Agric. Food Chem. 2008, 56, 7001–7009. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Wang, L.; Im, S.; Hwang, O.; Kim, H.-S.; Kang, M.-C.; Lee, S.-H. Anti-Obesity Effect of Diphlorethohydroxycarmalol Isolated from Brown Alga Ishige okamurae in High-Fat Diet-Induced Obese Mice. Mar. Drugs 2019, 17, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Kim, H.S.; Oh, J.Y.; Je, J.G.; Jeon, Y.-J.; Ryu, B. Protective effect of diphlorethohydroxycarmalol isolated from Ishige okamurae against UVB-induced damage in vitro in human dermal fibroblasts and in vivo in zebrafish. Food Chem. Toxicol. 2020, 136, 110963. [Google Scholar] [CrossRef]
- Heo, S.-J.; Ko, S.-C.; Kang, S.-M.; Cha, S.-H.; Lee, S.-H.; Kang, -H.; Jung, W.-K.; Affan, A.; Oh, C.; Jeon, Y.-J. Inhibitory effect of diphlorethohydroxycarmalol on melanogenesis and its protective effect against UV-B radiation-induced cell damage. Food Chem. Toxicol. 2010, 48, 1355–1361. [Google Scholar] [CrossRef]
- Piao, M.J.; Kang, K.A.; Kim, K.C.; Chae, S.; Kim, G.O.; Shin, T.; Kim, H.S.; Hyun, J.W. Diphlorethohydroxycarmalol attenuated cell damage against UVB radiation via enhancing antioxidant effects and absorbing UVB ray in human HaCaT keratinocytes. Environ. Toxicol. Pharmacol. 2013, 36, 680–688. [Google Scholar] [CrossRef]
- Zhen, A.X.; Piao, M.J.; Hyun, Y.J.; Kang, K.A.; Fernando, P.D.S.M.; Cho, S.J.; Ahn, M.J.; Hyun, J.W. Diphlorethohydroxycarmalol Attenuates Fine Particulate Matter-Induced Subcellular Skin Dysfunction. Mar. Drugs 2019, 17, 95. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.-E.; Choi, H.; Shin, D.W.; Na, H.-W.; Park, N.Y.; Kim, J.B.; Jo, D.S.; Cho, M.J.; Lyu, J.H.; Chang, J.H.; et al. Fine particulate matter (PM2.5) inhibits ciliogenesis by increasing SPRR3 expression via c-Jun activation in RPE cells and skin keratinocytes. Sci. Rep. 2019, 9, 3994. [Google Scholar] [CrossRef]
- Ryu, Y.S.; Kang, K.A.; Piao, M.J.; Ahn, M.J.; Yi, J.M.; Hyun, Y.-M.; Kim, S.H.; Ko, M.K.; Park, C.O.; Hyun, J.W. Particulate matter induces inflammatory cytokine production via activation of NFκB by TLR5-NOX4-ROS signaling in human skin keratinocyte and mouse skin. Redox Boil. 2019, 21, 101080. [Google Scholar] [CrossRef]
- Zhang, M.; Hwang, E.; Lin, P.; Gao, W.; Ngo, H.T.; Yi, T.-H. Prunella vulgaris L. Exerts a Protective Effect against Extrinsic Aging Through NF-κB, MAPKs, AP-1, and TGF-β/Smad Signaling Pathways in UVB-Aged Normal Human Dermal Fibroblasts. Rejuvenation Res. 2018, 21, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S.; Bowden, G. Ultraviolet B Regulation of Transcription Factor Families: Roles of Nuclear Factor-kappa B (NF-κB) and Activator Protein-1 (AP-1) in UVB-Induced Skin Carcinogenesis. Curr. Cancer Drug Targets 2007, 7, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Fernando, K.H.N.; Yang, H.-W.; Jiang, Y.; Jeon, Y.-J.; Ryu, B. Diphlorethohydroxycarmalol Isolated from Ishige okamurae Represses High Glucose-Induced Angiogenesis In Vitro and In Vivo. Mar. Drugs 2018, 16, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Oh, J.Y.; Fernando, S.; Sanjeewa, K.K.A.; Kim, E.-A.; Lee, W.; Jeon, Y.-J. Soft corals collected from Jeju Island; a potential source of anti-inflammatory phytochemicals. J. Chitin Chitosan 2016, 21, 247–254. [Google Scholar] [CrossRef]
- Dong, H.; Zheng, L.; Yu, P.; Jiang, Q.; Wu, Y.; Huang, C.; Yin, B. Characterization and Application of Lignin–Carbohydrate Complexes from Lignocellulosic Materials as Antioxidants for Scavenging In Vitro and In Vivo Reactive Oxygen Species. ACS Sustain. Chem. Eng. 2019, 8, 256–266. [Google Scholar] [CrossRef]
- Huang, C.; Wang, X.; Liang, C.; Jiang, X.; Yang, G.; Xu, J.; Yong, Q. A sustainable process for procuring biologically active fractions of high-purity xylooligosaccharides and water-soluble lignin from Moso bamboo prehydrolyzate. Biotechnol. Biofuels 2019, 12, 189. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Pei, W.; Tang, S.; Yan, F.; Peng, Z.; Huang, C.; Yang, J.; Yong, Q. Procuring biologically active galactomannans from spent coffee ground (SCG) by autohydrolysis and enzymatic hydrolysis. Int. J. Boil. Macromol. 2020, 149, 572–580. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Kim, H.S.; Je, J.-G.; Oh, J.Y.; Kim, Y.-S.; Cha, S.-H.; Jeon, Y.-J. Protective Effect of Diphlorethohydroxycarmalol Isolated from Ishige okamurae Against Particulate Matter-Induced Skin Damage by Regulation of NF-κB, AP-1, and MAPKs Signaling Pathways In Vitro in Human Dermal Fibroblasts. Molecules 2020, 25, 1055. https://doi.org/10.3390/molecules25051055
Wang L, Kim HS, Je J-G, Oh JY, Kim Y-S, Cha S-H, Jeon Y-J. Protective Effect of Diphlorethohydroxycarmalol Isolated from Ishige okamurae Against Particulate Matter-Induced Skin Damage by Regulation of NF-κB, AP-1, and MAPKs Signaling Pathways In Vitro in Human Dermal Fibroblasts. Molecules. 2020; 25(5):1055. https://doi.org/10.3390/molecules25051055
Chicago/Turabian StyleWang, Lei, Hyun Soo Kim, Jun-Geon Je, Jae Young Oh, Young-Sang Kim, Seon-Heui Cha, and You-Jin Jeon. 2020. "Protective Effect of Diphlorethohydroxycarmalol Isolated from Ishige okamurae Against Particulate Matter-Induced Skin Damage by Regulation of NF-κB, AP-1, and MAPKs Signaling Pathways In Vitro in Human Dermal Fibroblasts" Molecules 25, no. 5: 1055. https://doi.org/10.3390/molecules25051055
APA StyleWang, L., Kim, H. S., Je, J. -G., Oh, J. Y., Kim, Y. -S., Cha, S. -H., & Jeon, Y. -J. (2020). Protective Effect of Diphlorethohydroxycarmalol Isolated from Ishige okamurae Against Particulate Matter-Induced Skin Damage by Regulation of NF-κB, AP-1, and MAPKs Signaling Pathways In Vitro in Human Dermal Fibroblasts. Molecules, 25(5), 1055. https://doi.org/10.3390/molecules25051055