
Effect of Instant Controlled Pressure-Drop on the Non-Nutritional Compounds of Seeds and Sprouts of Common Black Bean (Phaseolus vulgaris L.)

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Non-Nutritional Compounds in Black Bean without Treatment (BNT), Germinated-Lyophilized (BGL), and Germinated-Dried (BGD)
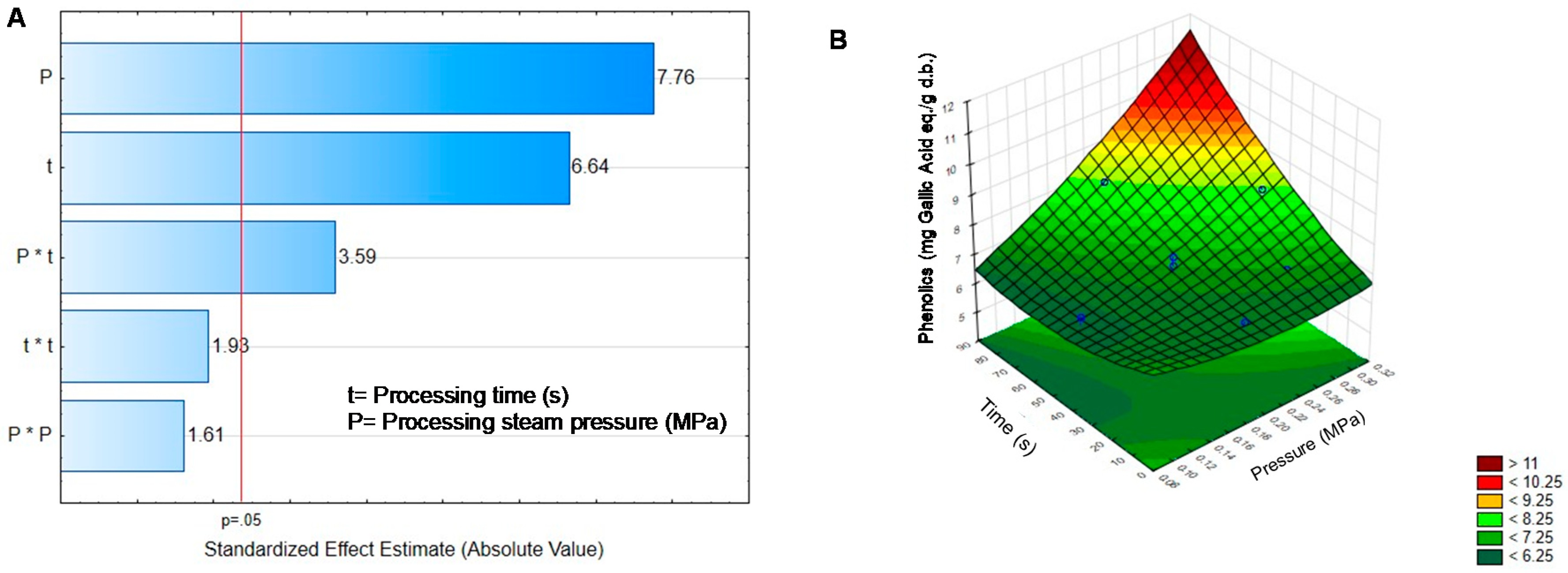
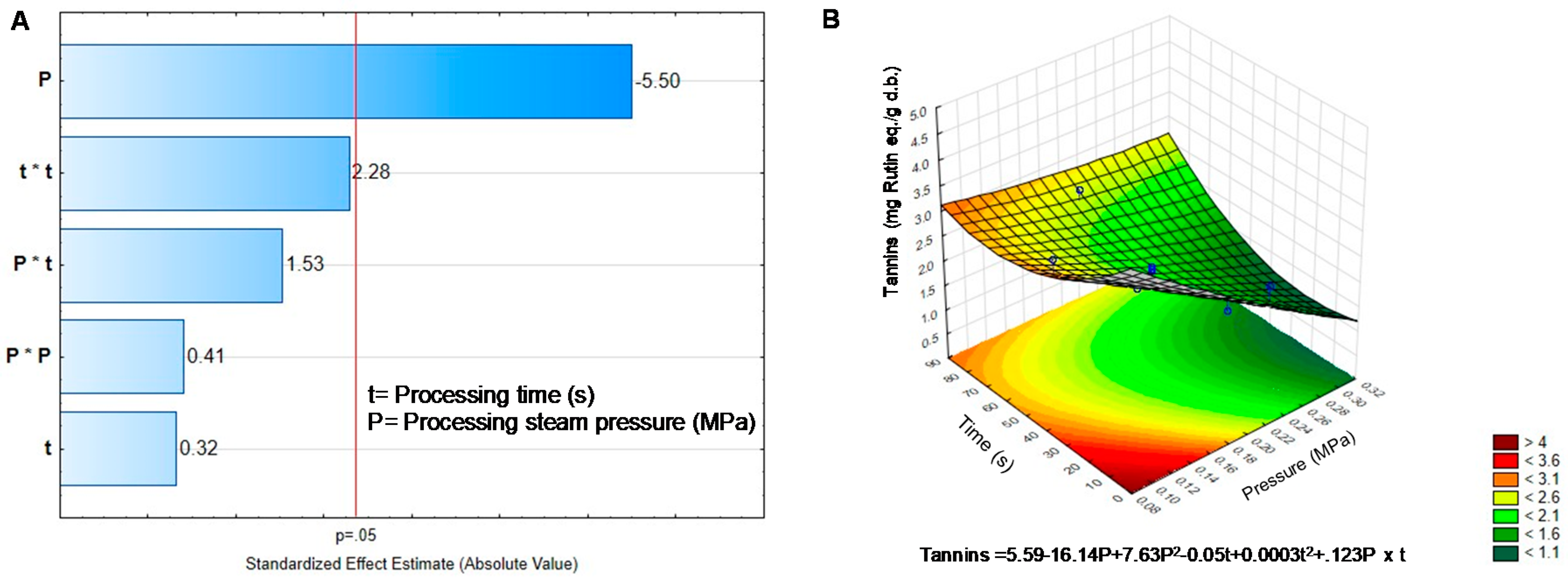
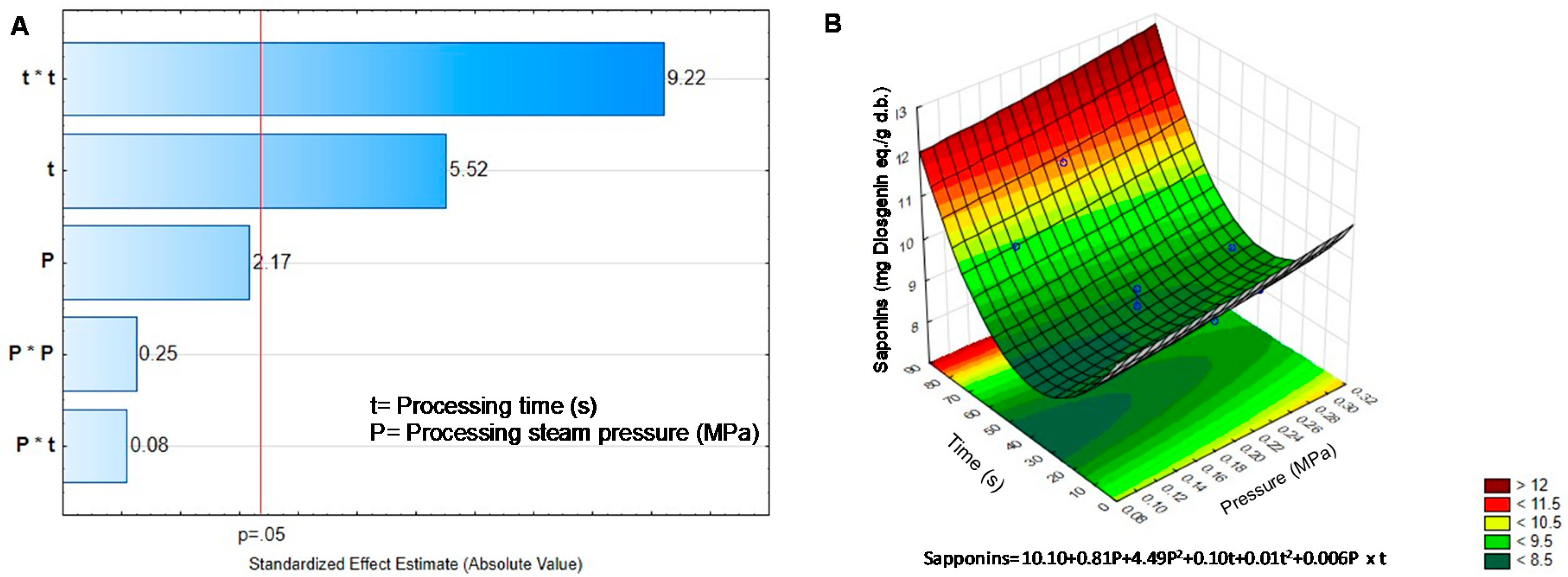
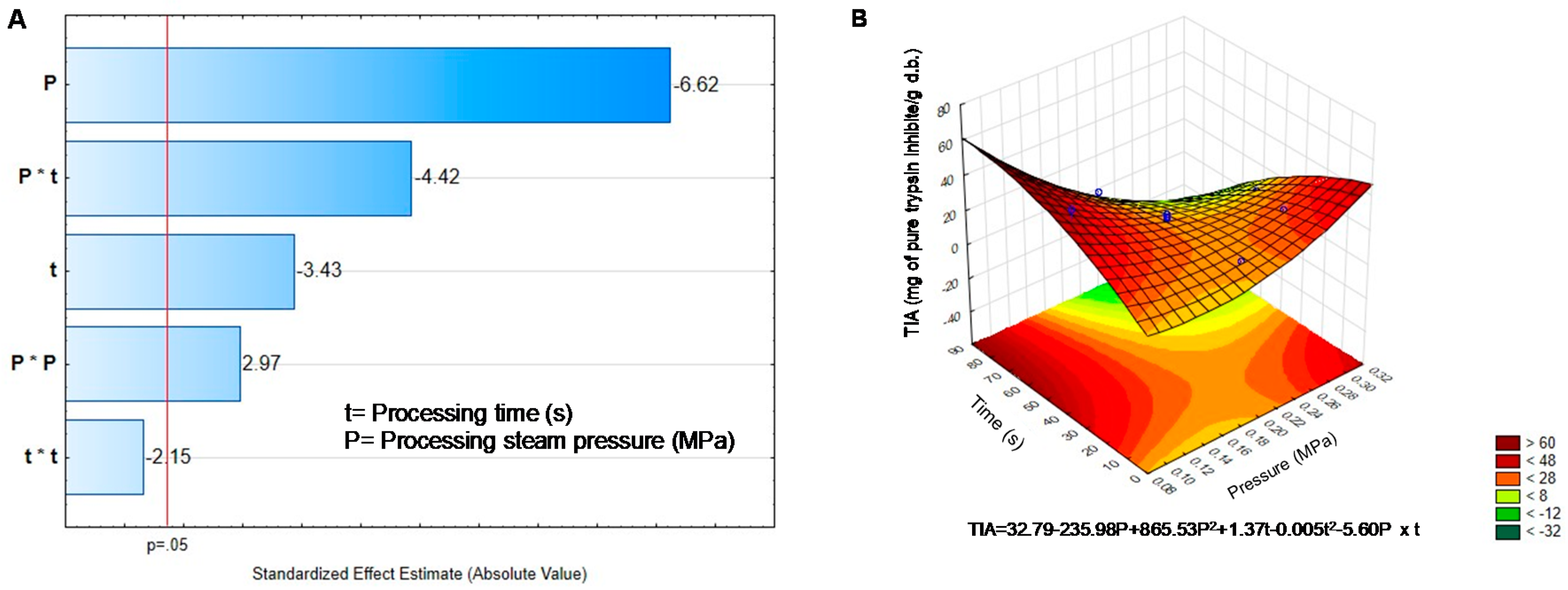
2.2. Effect of DIC on the Non-Nutritional Compounds of Black Bean Sprouts
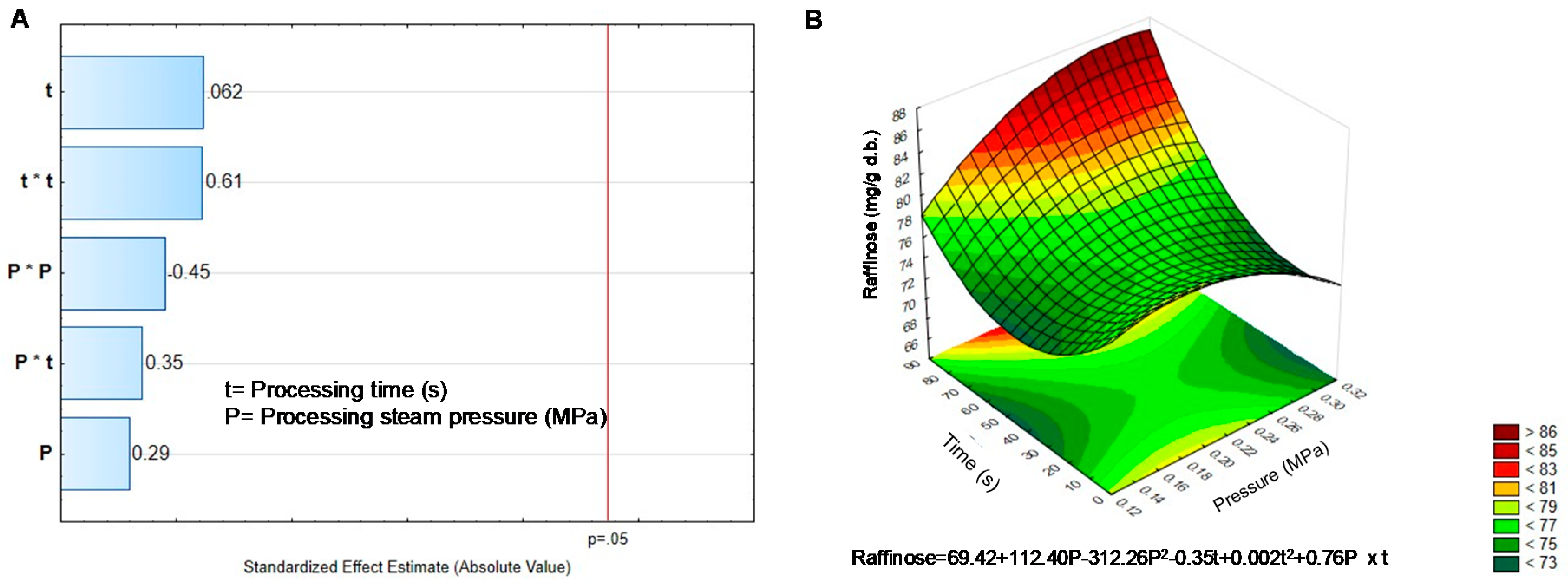
2.3. Determination of α-Galactosides in Black Bean Sprouts Treated with DIC
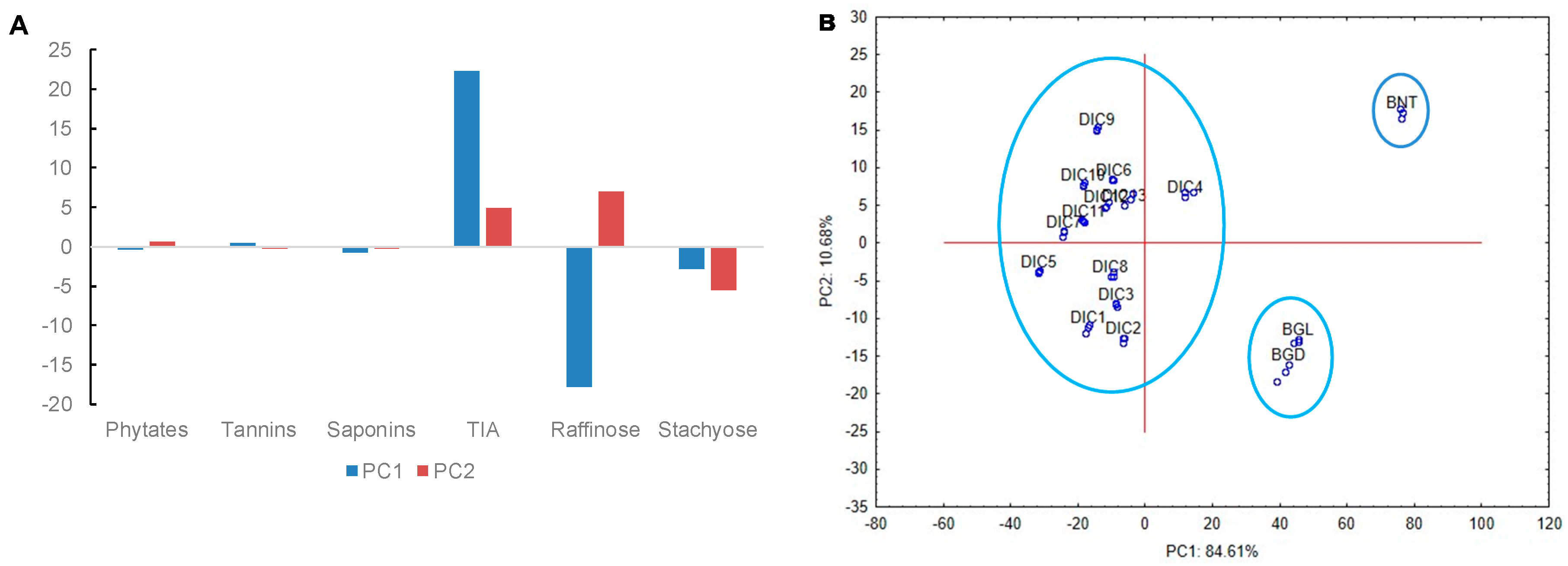
2.4. Principal Component Analysis
3. Materials and Methods
3.1. Germination
3.2. DIC Treatment of Dried Black Bean Sprouts
3.3. Non-Nutritional Compounds Quantification
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Do Evangelho, J.A.; Vanier, N.L.; Pinto, V.Z.; De Berrios, J.J.; Dias, A.R.G.; da Rosa Zavareze, E. Black bean (Phaseolus vulgaris L.) protein hydrolysates: Physicochemical and functional properties. Food Chem. 2017, 214, 460–467. [Google Scholar] [CrossRef]
- Muzquiz, M.; Varela, A.; Burbano, C.; Cuadrado, C.; Guillamón, E.; Pedrosa, M.M. Bioactive compounds in legumes: Pronutritive and antinutritive actions. Implications for nutrition and health. Phytochem. Rev. 2012, 11, 227–244. [Google Scholar] [CrossRef]
- Smith, M.R.; Veneklaas, E.; Polania, J.; Rao, I.M.; Beebe, S.E.; Merchant, A. Field drought conditions impact yield but not nutritional quality of the seed in common bean (Phaseolus vulgaris L.). PLoS ONE 2019, 14, e0217099. [Google Scholar] [CrossRef] [Green Version]
- El-Niely, H.F. Effect of radiation processing on antinutrients, in-vitro protein digestibility and protein efficiency ratio bioassay of legume seeds. Radiat. Phys. Chem. 2007, 76, 1050–1057. [Google Scholar] [CrossRef]
- Kotue, T.; Marlyne Josephine, M.; Wirba, L.; Amalene, S.; Nkenmeni, D.; Fokou, E.; Fokam, D. Nutritional properties and nutrients chemical analysis of common beans seed. MOJ Biol. Med. 2018, 32, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Pedrosa, M.M.; Cuadrado, C.; Burbano, C.; Allaf, K.; Haddad, J.; Gelencsér, E.; Takács, K.; Guillamón, E.; Muzquiz, M. Effect of instant controlled pressure drop on the oligosaccharides, inositol phosphates, trypsin inhibitors and lectins contents of different legumes. Food Chem. 2012, 131, 862–868. [Google Scholar] [CrossRef]
- López, A.; El-Naggar, T.; Dueñas, M.; Ortega, T.; Estrella, I.; Hernández, T.; Gómez-Serranillos, M.P.; Palomino, O.M.; Carretero, M.E. Effect of cooking and germination on phenolic composition and biological properties of dark beans (Phaseolus vulgaris L.). Food Chem. 2013, 138, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Haddad, J.; Greiner, R.; Allaf, K. Effect of instantaneous controlled pressure drop on the phytate content of lupin. LWT Food Sci. Technol. 2007, 40, 448–453. [Google Scholar] [CrossRef]
- Haddad, J.; Allaf, K. A study of the impact of instantaneous controlled pressure drop on the trypsin inhibitors of soybean. J. Food Eng. 2007, 79, 353–357. [Google Scholar] [CrossRef]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef]
- Zhang, G.; Xu, Z.; Gao, Y.; Huang, X.; Zou, Y.; Yang, T. Effects of Germination on the Nutritional Properties, Phenolic Profiles, and Antioxidant Activities of Buckwheat. J. Food Sci. 2015, 80, H1111–H1119. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Bi, J.F.; Peng, J.; Allaf, T.; Allaf, K. Advances of Instant Controlled Pressure Drop (DIC) Technique in Agro-product Processing. Mod. Food Sci. Technol. 2017, 33, 311–318. [Google Scholar] [CrossRef]
- Hernandez-Aguirre, A.I.; Tellez-Perez, C.; San Martin-Azocar, A.; Cardador-Martinez, A. Effect of Instant Controlled Pressure-Drop (DIC), Cooking and Germination on Non-Nutritional Factors of Common Vetch (Vicia sativa spp.). Molecules 2019, 25, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Bi, J.; Jin, X.; Wu, X.; Lyu, J.; Li, X.; Hu, L. Effect of the moisture equilibrium process on the expansion behavior of instant controlled pressure drop (DIC) drying of dried apple cubes. J. Sci. Food Agric. 2019. [Google Scholar] [CrossRef]
- Cuadrado, C.; Cabanillas, B.; Pedrosa, M.M.; Muzquiz, M.; Haddad, J.; Allaf, K.; Rodriguez, J.; Crespo, J.F.; Burbano, C. Effect of instant controlled pressure drop on IgE antibody reactivity to peanut, lentil, chickpea and soybean proteins. Int. Arch. Allergy Immunol. 2011, 156, 397–404. [Google Scholar] [CrossRef]
- Cardador-Martínez, A.; Loarca-Piña, G.; Oomah, B.D. Antioxidant activity in common beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2002, 50, 6975–6980. [Google Scholar] [CrossRef]
- Oomah, B.D.; Caspar, F.; Malcolmson, L.J.; Bellido, A.-S. Phenolics and antioxidant activity of lentil and pea hulls. Food Res. Int. 2011, 44, 436–441. [Google Scholar] [CrossRef]
- Pająk, P.; Socha, R.; Gałkowska, D.; Rożnowski, J.; Fortuna, T. Phenolic profile and antioxidant activity in selected seeds and sprouts. Food Chem. 2014, 143, 300–306. [Google Scholar] [CrossRef]
- Xu, M.; Jin, Z.; Ohm, J.B.; Schwarz, P.; Rao, J.; Chen, B. Improvement of the Antioxidative Activity of Soluble Phenolic Compounds in Chickpea by Germination. J. Agric. Food Chem. 2018, 66, 6179–6187. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Frias, J.; Zielinski, H.; Piskula, M.K.; Kozlowska, H.; Vidal-Valverde, C. Kinetic study of the antioxidant compounds and antioxidant capacity during germination of Vigna radiata cv. emmerald, Glycine max cv. jutro and Glycine max cv. merit. Food Chem. 2008, 111, 622–630. [Google Scholar] [CrossRef]
- Randhir, R.; Lin, Y.-T.; Shetty, K. Stimulation of phenolics, antioxidant and antimicrobial activities in dark germinated mung bean sprouts in response to peptide and phytochemical elicitors. Process Biochem. 2004, 39, 637–646. [Google Scholar] [CrossRef]
- Yeo, J.; Shahidi, F. Critical Evaluation of Changes in the Ratio of Insoluble Bound to Soluble Phenolics on Antioxidant Activity of Lentils during Germination. J. Agric. Food Chem. 2015, 63, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Gangoliya, S.S.; Singh, N.K. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 2015, 52, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Fouad, A.A.; Rehab, F. Effect of germination time on proximate analysis, bioactive compounds and antioxidant activity of lentil (Lens culinaris Medik.) sprouts. Acta Sci. Polonorum Technol. Aliment. 2015, 14, 233–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frontela, C.; Ros, G.; Martínez, C. Empleo de fitasas como ingrediente funcional en alimentos. Use of phytases as a functional ingredient in food. Mundo Aliment. 2010, 58, 6–11. [Google Scholar]
- Khattak, A.B.; Zeb, A.; Bibi, N.; Khalil, S.A.; Khattak, M.S. Influence of germination techniques on phytic acid and polyphenols content of chickpea (Cicer arietinum L.) sprouts. Food Chem. 2007, 104, 1074–1079. [Google Scholar] [CrossRef]
- Shimelis, E.A.; Rakshit, S.K. Effect of processing on antinutrients and in vitro protein digestibility of kidney bean (Phaseolus vulgaris L.) varieties grown in East Africa. Food Chem. 2007, 103, 161–172. [Google Scholar] [CrossRef]
- Rusydi, M.R.; Noraliza, C.W.; Azlan, A.; Amom, Z. Nutritional changes in germinated legumes and rice varieties. Int. Food Res. J. 2011, 18, 705–713. [Google Scholar]
- Paucar-Menacho, L.M.; Berhow, M.A.; Mandarino, J.M.G.; Chang, Y.K.; Mejia, E.G.d. Effect of time and temperature on bioactive compounds in germinated Brazilian soybean cultivar BRS 258. Food Res. Int. 2010, 43, 1856–1865. [Google Scholar] [CrossRef] [Green Version]
- Trugo, L.C.; Donangelo, C.M.; Trugo, N.M.F.; Bach Knudsen, K.E. Effect of Heat Treatment on Nutritional Quality of Germinated Legume Seeds. J. Agric. Food Chem. 2000, 48, 2082–2086. [Google Scholar] [CrossRef]
- Sangronis, E.; Machado, C.J. Influence of germination on the nutritional quality of Phaseolus vulgaris and Cajanus cajan. LWT Food Sci. Technol. 2007, 40, 116–120. [Google Scholar] [CrossRef]
- Alajaji, S.A.; El-Adawy, T.A. Nutritional composition of chickpea (Cicer arietinum L.) as affected by microwave cooking and other traditional cooking methods. J. Food Compos. Anal. 2006, 19, 806–812. [Google Scholar] [CrossRef]
- Sánchez-Chino, X.M.; Jiménez-Martínez, C.; Vásquez-Garzón, V.R.; Álvarez-González, I.; Villa-Treviño, S.; Madriga-Blujaidar, E.; Dávila-Ortiz, G.; Baltiérrez-Hoyos, R. Cooked chickpea consumption inhibits colon carcinogenesis in mice induced with azoxymethane and dextran sulfate sodium. J. Am. Coll. Nutr. 2017, 36, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Alonzo-Macías, M.; Cardador-Martínez, A.; Mounir, S.; Montejano-Gaitán, G.; Allaf, K. Comparative study of the effects of drying methods on antioxidant activity of dried strawberry (Fragaria Var. Camarosa). J. Food Res. 2013, 2, 92–107. [Google Scholar] [CrossRef]
- Gahler, S.; Otto, K.; Böhm, V. Alterations of Vitamin C, Total Phenolics, and Antioxidant Capacity as Affected by Processing Tomatoes to Different Products. J. Agric. Food Chem. 2003, 51, 7962–7968. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.J.; Bozonnet, S.; Mullen, W.; Jenkins, G.I.; Lean, M.E.J.; Crozier, A. Occurrence of Flavonols in Tomatoes and Tomato-Based Products. J. Agric. Food Chem. 2000, 48, 2663–2669. [Google Scholar] [CrossRef]
- Świeca, M.; Gawlik-Dziki, U. Effects of sprouting and postharvest storage under cool temperature conditions on starch content and antioxidant capacity of green pea, lentil and young mung bean sprouts. Food Chem. 2015, 185, 99–105. [Google Scholar] [CrossRef]
- Mubarak, A.E. Nutritional composition and antinutritional factors of mung bean seeds (Phaseolus aureus) as affected by some home traditional processes. Food Chem. 2005, 89, 489–495. [Google Scholar] [CrossRef]
- Khandelwal, S.; Udipi, S.A.; Ghugre, P. Polyphenols and tannins in Indian pulses: Effect of soaking, germination and pressure cooking. Food Res. Int. 2010, 43, 526–530. [Google Scholar] [CrossRef]
- Sánchez, M.N. Efecto de la Cocción y de los Compuestos Fenólicos Sobre el Valor Biológico de la Proteína de frijol. In Effect of Cooking and Phenolic Compounds on the Biological Value of Bean Protein; Instituto Politécnico Nacional: Mexico City, Mexico , 2012. [Google Scholar]
- Guajardo-Flores, D.; García-Patiño, M.; Serna-Guerrero, D.; Gutiérrez-Uribe, J.; Serna-Saldívar, S. Characterization and quantification of saponins and flavonoids in sprouts, seed coats and cotyledons of germinated black beans. Food Chem. 2012, 134, 1312–1319. [Google Scholar] [CrossRef]
- Reim, V.; Rohn, S. Characterization of saponins in peas (Pisum sativum L.) by HPTLC coupled to mass spectrometry and a hemolysis assay. Food Res. Int. 2015, 76, 3–10. [Google Scholar] [CrossRef]
- Avilés-Gaxiola, S.; Chuck-Hernández, C.; Serna Saldívar, S.O. Inactivation Methods of Trypsin Inhibitor in Legumes: A Review. J. Food Sci. 2018, 83, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Amor, B.B.; Lamy, C.; Andre, P.; Allaf, K. Effect of instant controlled pressure drop treatments on the oligosaccharides extractability and microstructure of Tephrosia purpurea seeds. J. Chromatogr. A 2008, 1213, 118–124. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Frías, J.; Gómez, R.; Vidal-Valverde, C. Influence of addition of raffinose family oligosaccharides on probiotic survival in fermented milk during refrigerated storage. Int. Dairy J. 2006, 16, 768–774. [Google Scholar] [CrossRef]
- De Cortes Sánchez, M.A.; Altares, P.; Pedrosa, M.M.; Burbano, C.; Cuadrado, C.; Goyoaga, C.; Muzquiz, M.; Jiménez-Martínez, C.; Dávila-Ortiz, G. Alkaloid variation during germination in different lupin species. Food Chem. 2005, 90, 347–355. [Google Scholar] [CrossRef]
- Guillamon, E.; Burbano, C.; Cuadrado, C.; Muzquiz, M.; Pedrosa, M.M.; Sanchez, M.; Cabanillas, B.; Crespo, J.F.; Rodriguez, J.; Haddad, J.; et al. Effect of an instantaneous controlled pressure drop on in vitro allergenicity to lupins (Lupinus albus var Multolupa). Int. Arch. Allergy Immunol. 2008, 145, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Hiai, S.; Oura, H.; Nakajima, T. Color reaction of some sapogenins and saponins with vanillin and sulfuric acid. Planta Med. 1976, 29, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Vaintraub, I.A.; Lapteva, N.A. Colorimetric determination of phytate in unpurified extracts of seeds and the products of their processing. Anal. Biochem. 1988, 175, 227–230. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Abdel-Aal, E.S.; Hucl, P. A rapid method for quantifying total anthocyanins in blue aleurone and purple pericarp wheats. Cereal Chem. 1999, 76, 350–354. [Google Scholar] [CrossRef]
- Smith, C.; van Megen, W.; Twaalfhoven, L.; Hitchcock, C. The determination of trypsin inhibitor levels in foodstuffs. J. Sci. Food Agric. 1980, 31, 341–350. [Google Scholar] [CrossRef]
- Liu, K.; Markakis, P. Trypsin inhibition assay as related to limited hydrolysis of inhibitors. Anal. Biochem. 1989, 178, 159–165. [Google Scholar] [CrossRef]
- Deshpande, S.S.; Cheryan, M. Evaluation of Vanillin Assay for Tannin Analysis of Dry Beans. J. Food Sci. 1985, 50, 905–910. [Google Scholar] [CrossRef]
- Muzquiz, M.; Rey, C.; Cuadrado, C.; Fenwick, G.R. Effect of germination on the oligosaccharide content of lupin species. J. Chromatogr. A 1992, 607, 349–352. [Google Scholar] [CrossRef]
- Johansen, H.N.; Glitsø, V.; Bach Knudsen, K.E. Influence of extraction solvent and temperature on the quantitative determination of oligosaccharides from plant materials by high-performance liquid chromatography. J. Agric. Food Chem. 1996, 44, 1470–1474. [Google Scholar] [CrossRef]
Sample Availability: Samples of the black beans germinated and DIC treated are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | BNT | BGL | BGD |
|---|---|---|---|
| Phenolics 1 | 4.11 ± 0.07 a | 8.20 ± 0.32 b | 8.35 ± 0.38 b |
| Phytic acid 2 | 17.33 ± 0.20 a | 12.80 ± 0.71 b | 13.44 ± 1.10 b |
| Tannins 3 | 2.62 ± 0.06 a | 4.55 ± 0.20 b | 4.47 ± 0.25 b |
| Saponins 4 | 7.05 ± 0.12 a | 9.37 ± 0.19 b | 9.47 ± 0.31 b |
| TIA 5 | 99.65 ± 0.43 a | 58.11 ± 0.80 b | 53.07 ± 1.15 b |
| Total oligosaccharides 6 | 44.43 ± 0.18 a | 48.50 ± 0.19 b | 47.01 ± 0.01 b |
| Raffinose | 6.26 ± 0.06 a | 28.95 ± 0.44 b | 27.37 ± 0.19 b |
| Stachyose | 33.11 ± 0.67 a | 19.55 ± 0.06 b | 19.64 ± 0.14 b |
| Verbascose | 5.06 ± 0.60 | ND | ND |
| Treatment | Pressure | Time | Phenolics | Phytates | Tannins | Saponins | TIA |
|---|---|---|---|---|---|---|---|
| BGD | - | - | 7.74 ± 0309 | 12.46 ± 0.20 | 4.15 ± 0.25 | 8.78 ± 0.39 | 53.07 ± 1.15 |
| DIC1 | 0.30 | 45 | 8.20 ± 0.10 * | 14.99 ± 0.13 * | 1.14 ± 0.11 * | 9.02 ± 0.10 | 15.30 ± 0.52 * |
| DIC2 | 0.20 | 45 | 6.95 ± 0.25 * | 13.73 ± 0.32 * | 2.05 ± 0.09 * | 8.67 ± 0.09 | 25.59 ± 0.18 * |
| DIC3 | 0.20 | 45 | 6.98 ± 0.09 * | 12.72 ± 0.16 | 1.96 ± 0.07 * | 8.32 ± 0.04 * | 24.40 ± 0.13 * |
| DIC4 | 0.10 | 45 | 6.63 ± 0.04 * | 14.19 ± 0.17 * | 3.10 ± 0.15 * | 8.22 ± 0.07 * | 49.62 ± 0.1.2 |
| DIC5 | 0.27 | 70 | 8.29 ± 0.07 * | 15.99 ± 0.22 * | 2.11 ± 0.09 * | 10.99 ± 013 * | 2.20 ± 0.07 * |
| DIC6 | 0.27 | 20 | 6.98 ± 0.13 * | 15.81 ± 0.15 * | 1.79 ± 0.12 * | 9.15 ± 0.07 * | 28.72 ± 0.24 * |
| DIC7 | 0.20 | 80 | 8.27 ± 0.07 * | 15.39 ± 0.51 * | 2.72 ± 0.11 * | 10.95 ± 0.06 * | 10.63 ± 0.61 * |
| DIC8 | 0.20 | 10 | 6.67 ± 0.06 * | 13.85 ± 0.11 * | 2.21 ± 0.16 * | 9.52 ± 0.12 * | 23.18 ± 0.24 * |
| DIC9 | 0.20 | 45 | 6.81 ± 0.12 * | 13.72 ± 0.17 * | 2.11 ± 0.11 * | 9.07 ± 0.05 | 21.72 ± 0.13 * |
| DIC10 | 0.20 | 45 | 6.70 ± 0.06 * | 13.62 ± 0.15 * | 2.11 ± 0.24 * | 8.99 ± 0.05 | 21.11 ± 0.31 * |
| DIC11 | 0.20 | 45 | 7.25 ± 0.05 * | 13.85 ± 0.13 * | 2.00 ± 0.06 * | 8.57 ± 0.28 | 19.16 ± 0.13 * |
| DIC12 | 0.13 | 20 | 6.46 ± 0.05 * | 13.31 ± 0.24 | 2.97 ± 0.11 * | 8.86 ± 0.16 | 25.58 ± 0.31 * |
| DIC13 | 0.13 | 70 | 6.59 ± 0.28 * | 14.97 ± 0.17 * | 2.29 ± 0.15 * | 9.98 ± 0.09 * | 31.38 ± 0.55 * |
| Treatment | Pressure | Time | Raffinose | Stachyose | Total Oligosaccharides |
|---|---|---|---|---|---|
| BGD | - | - | 27.37 ± 0.19 | 19.64 ± 014 | 47.01 ± 0.01 |
| DIC1 | 0.30 | 45 | 71.61 ± 0.18 * | 31.84 ± 0.81 * | 103.45 ± 0.93 * |
| DIC2 | 0.20 | 45 | 66.04 ± 0.25 * | 36.10 ± 0.28 * | 102.14 ± 0.15 * |
| DIC3 | 0.20 | 45 | 68.70 ± 0.77 * | 29.50 ± 0.39 * | 98.20 ± 0.21 * |
| DIC4 | 0.10 | 45 | 69.53 ± 0.17 * | 27.74 ± 0.13 * | 97.27 ± 0.90 * |
| DIC5 | 0.27 | 70 | 82.21 ± 0.39 * | 21.22 ± 0.43 * | 103.43 ± 0.60 * |
| DIC6 | 0.27 | 20 | 80.32 ± 0.15 * | 19.67 ± 0.45 | 99.99 ± 0.81 * |
| DIC7 | 0.20 | 80 | 81.51 ± 0.16 * | 18.12 ± 0.56 * | 99.63 ± 0.43 * |
| DIC8 | 0.20 | 10 | 72.23 ± 0.02 * | 27.72 ± 0.09 * | 99.95 ± 0.20 * |
| DIC9 | 0.20 | 45 | 81.80 ± 0.02 * | 2.69 ± 0.33 * | 84.49 ± 0.35 * |
| DIC10 | 0.20 | 45 | 84.80 ± 0.34 * | 19.61 ± 037 | 104.41 ± 0.38 * |
| DIC11 | 0.20 | 45 | 81.89 ± 0.04 * | 23.24 ± 0.07 * | 105.13 ± 0.27 * |
| DIC12 | 0.13 | 20 | 79.12 ± 0.14 * | 21.51 ± 0.26 * | 100.63 ± 0.30 * |
| DIC13 | 0.13 | 70 | 75.68 ± 0.17 * | 20.78 ± 0.04 * | 96.46 ± 0.18 * |
| Treatment | Pressure (MPa) | Time (s) |
|---|---|---|
| DIC1 | 0.30 | 45 |
| DIC2 | 0.20 | 45 |
| DIC3 | 0.20 | 45 |
| DIC4 | 0.10 | 45 |
| DIC5 | 0.27 | 70 |
| DIC6 | 0.27 | 20 |
| DIC7 | 0.20 | 80 |
| DIC8 | 0.20 | 10 |
| DIC9 | 0.20 | 45 |
| DIC10 | 0.20 | 45 |
| DIC11 | 0.20 | 45 |
| DIC12 | 0.13 | 20 |
| DIC13 | 0.13 | 70 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardador-Martínez, A.; Martínez-Tequitlalpan, Y.; Gallardo-Velazquez, T.; Sánchez-Chino, X.M.; Martínez-Herrera, J.; Corzo-Ríos, L.J.; Jiménez-Martínez, C. Effect of Instant Controlled Pressure-Drop on the Non-Nutritional Compounds of Seeds and Sprouts of Common Black Bean (Phaseolus vulgaris L.). Molecules 2020, 25, 1464. https://doi.org/10.3390/molecules25061464
Cardador-Martínez A, Martínez-Tequitlalpan Y, Gallardo-Velazquez T, Sánchez-Chino XM, Martínez-Herrera J, Corzo-Ríos LJ, Jiménez-Martínez C. Effect of Instant Controlled Pressure-Drop on the Non-Nutritional Compounds of Seeds and Sprouts of Common Black Bean (Phaseolus vulgaris L.). Molecules. 2020; 25(6):1464. https://doi.org/10.3390/molecules25061464
Chicago/Turabian StyleCardador-Martínez, Anaberta, Yara Martínez-Tequitlalpan, Tzayhri Gallardo-Velazquez, Xariss M. Sánchez-Chino, Jorge Martínez-Herrera, Luis Jorge Corzo-Ríos, and Cristian Jiménez-Martínez. 2020. "Effect of Instant Controlled Pressure-Drop on the Non-Nutritional Compounds of Seeds and Sprouts of Common Black Bean (Phaseolus vulgaris L.)" Molecules 25, no. 6: 1464. https://doi.org/10.3390/molecules25061464
APA StyleCardador-Martínez, A., Martínez-Tequitlalpan, Y., Gallardo-Velazquez, T., Sánchez-Chino, X. M., Martínez-Herrera, J., Corzo-Ríos, L. J., & Jiménez-Martínez, C. (2020). Effect of Instant Controlled Pressure-Drop on the Non-Nutritional Compounds of Seeds and Sprouts of Common Black Bean (Phaseolus vulgaris L.). Molecules, 25(6), 1464. https://doi.org/10.3390/molecules25061464