Rifamycin O, An Alternative Anti-Mycobacterium abscessus Agent


 ,
,
Abstract
:1. Introduction
2. Results
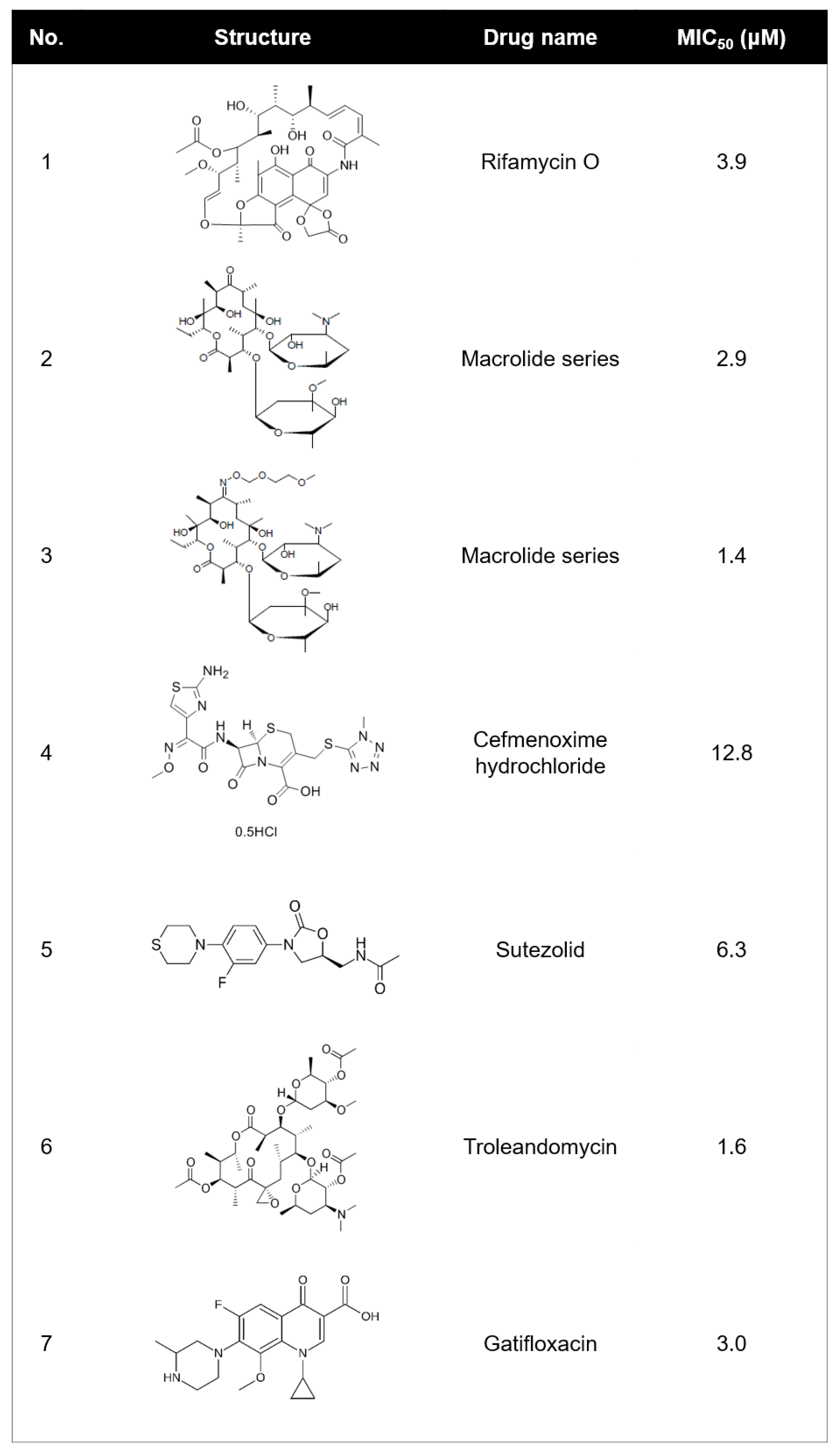
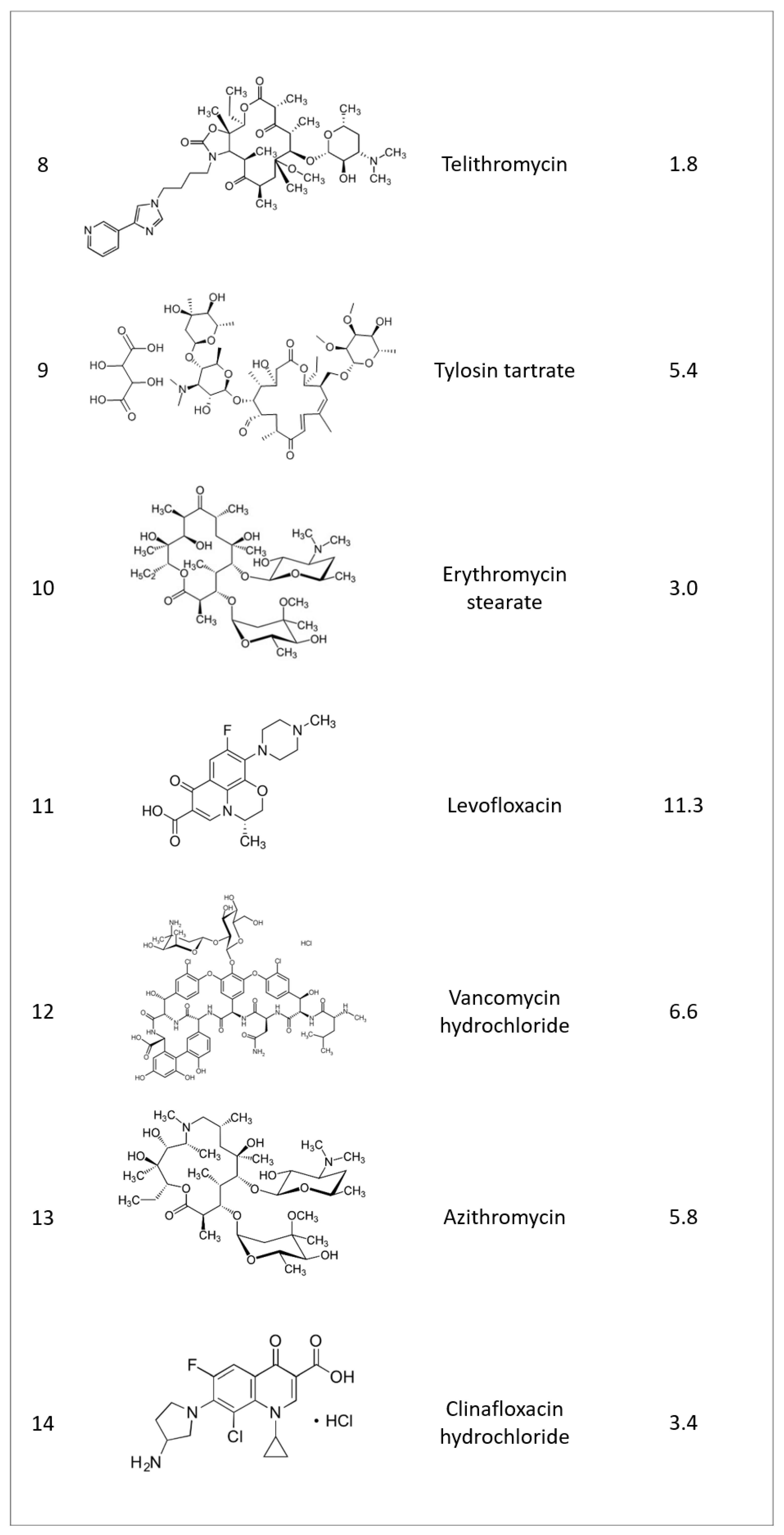
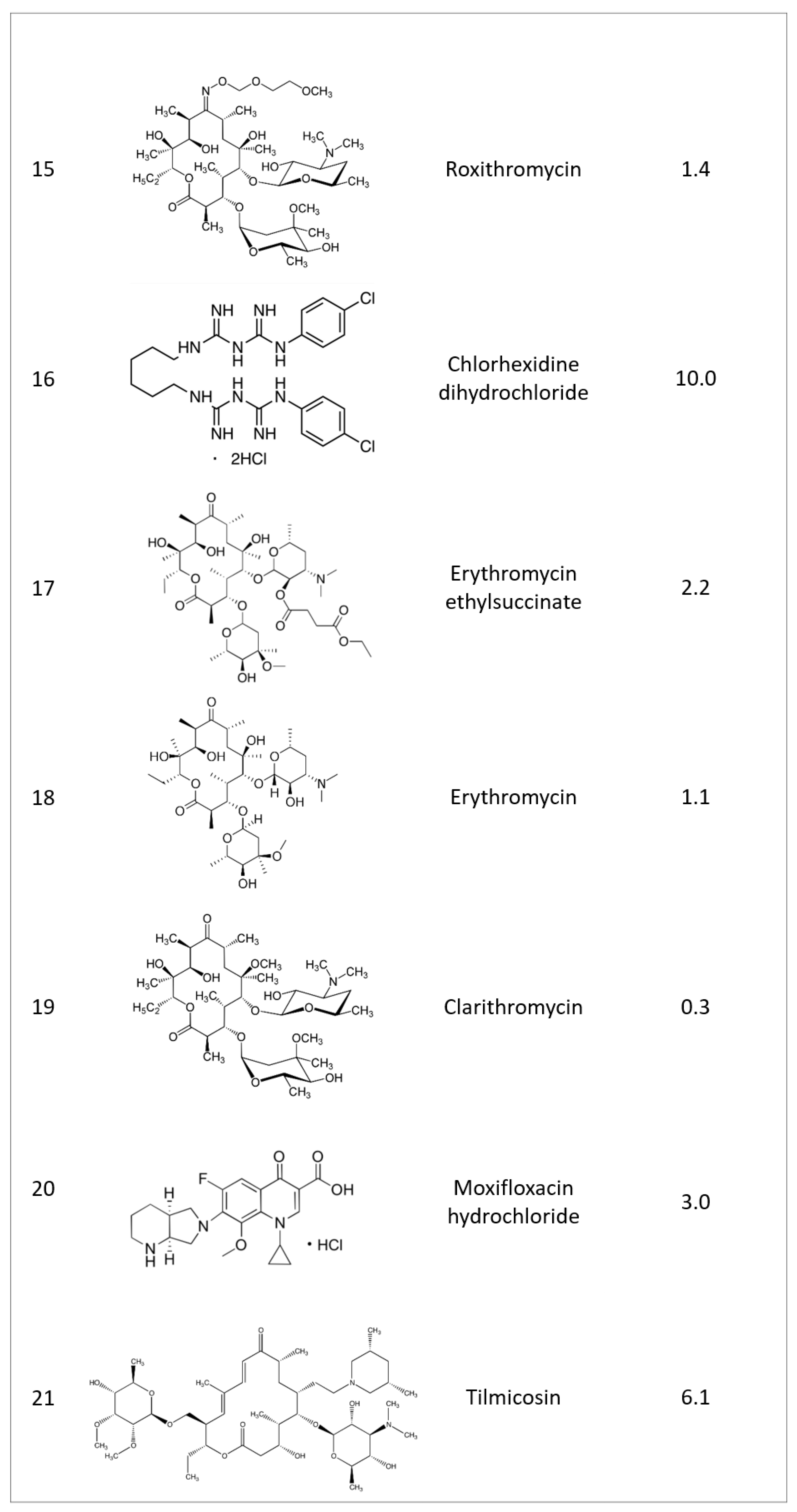
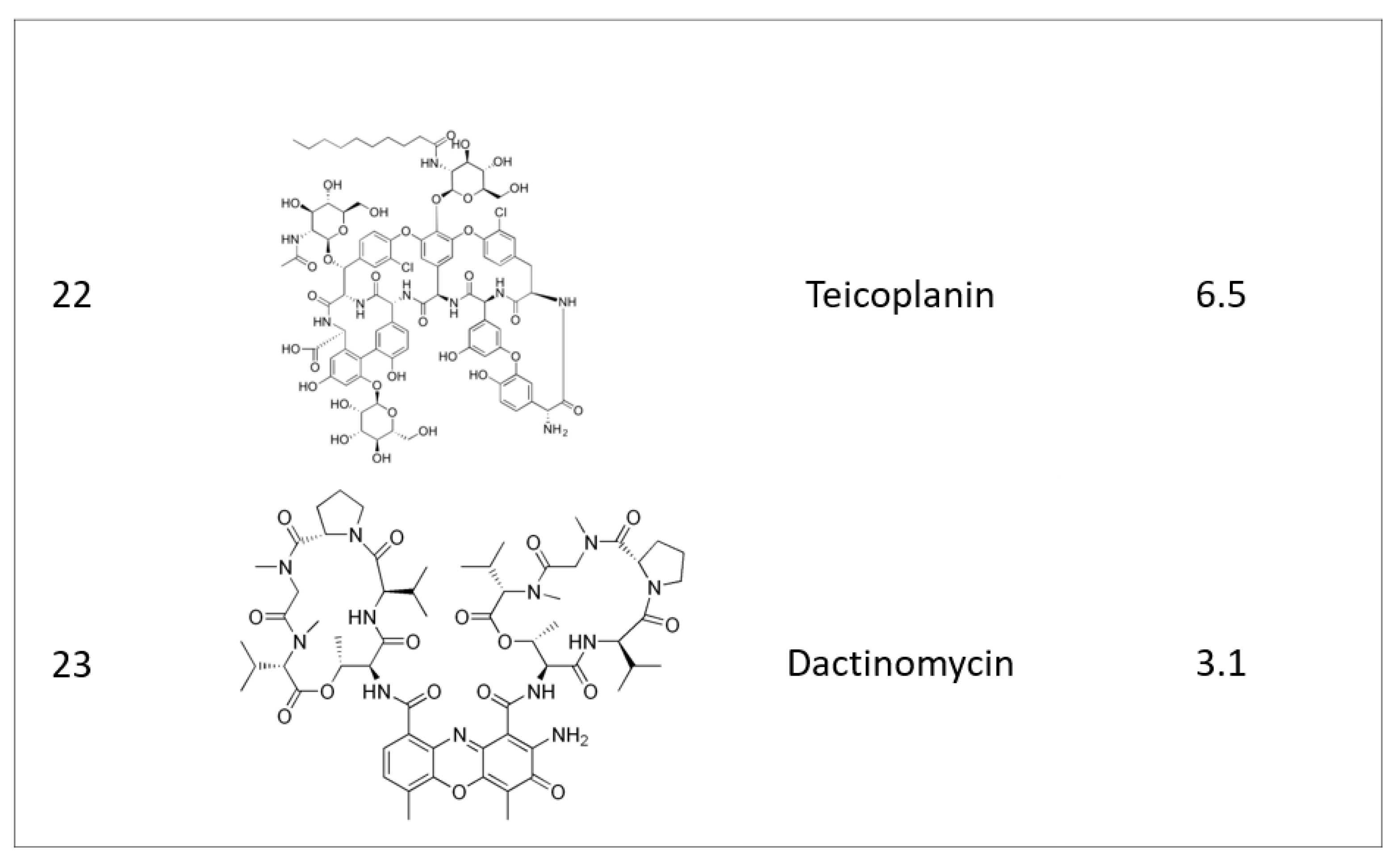
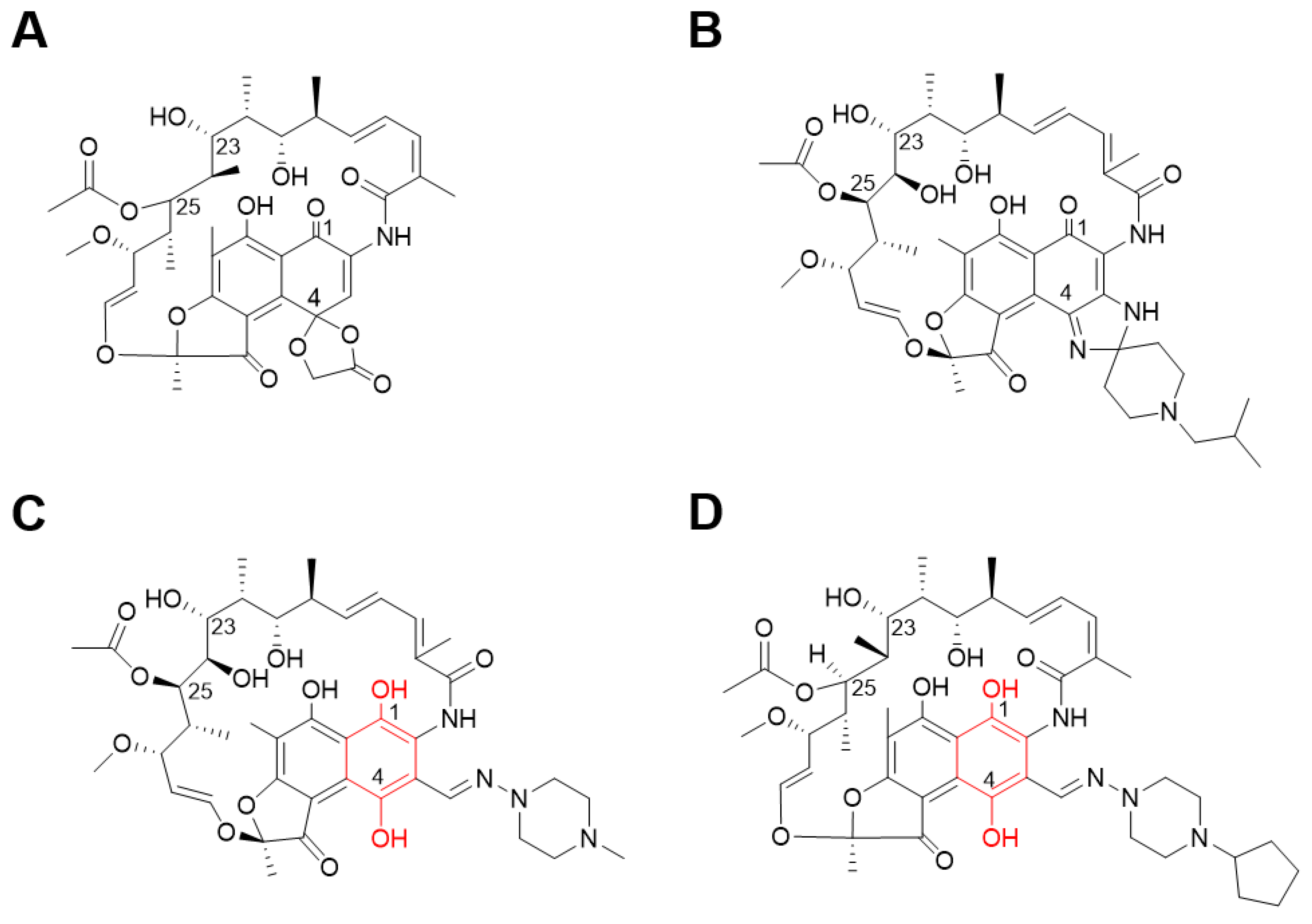
2.1. Identifying Active Compounds against Bioluminescent M. abscessus from a KCB Library
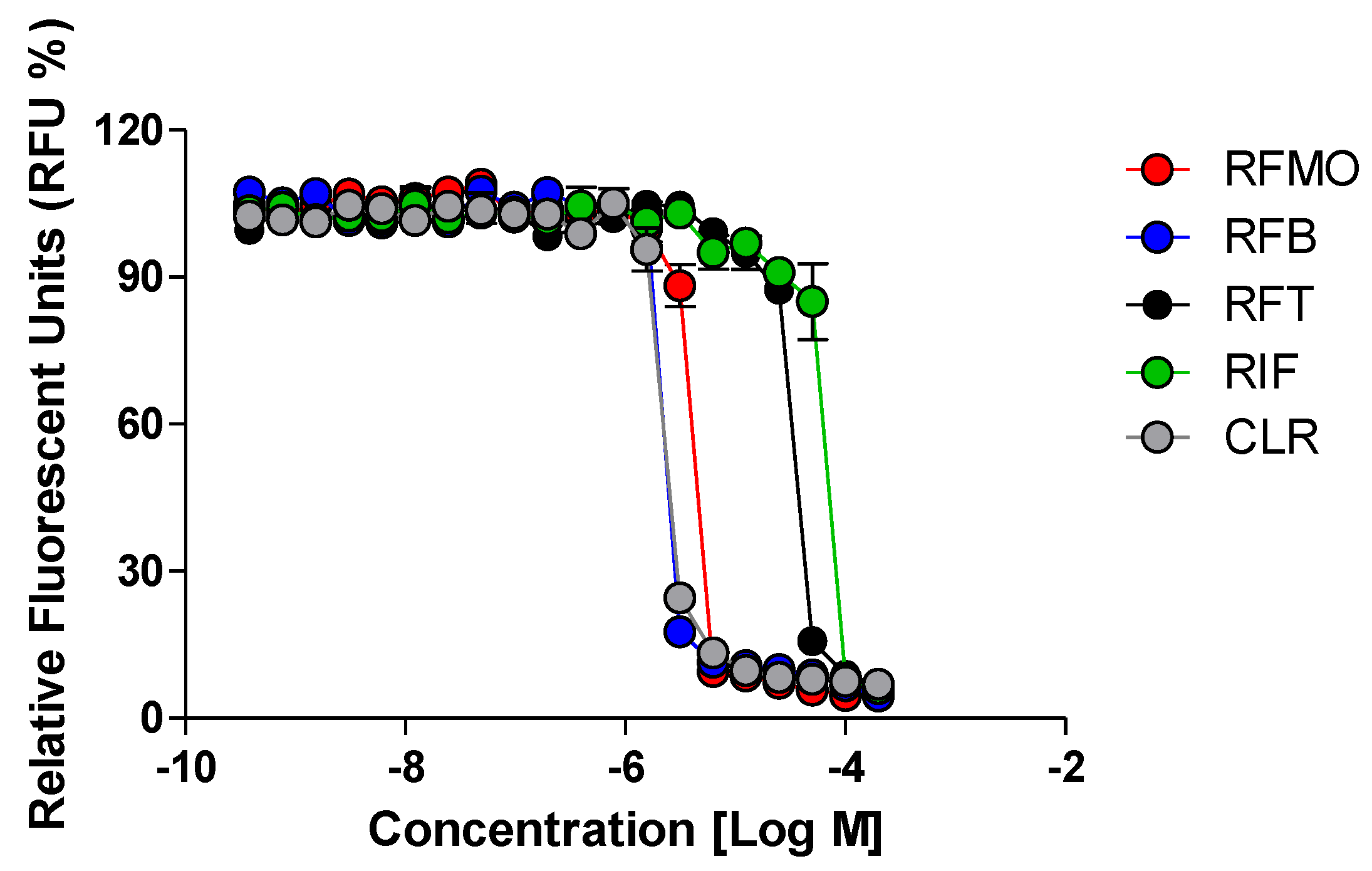
2.2. Rifamycin O is Non-toxic to Cells and Inhibits In Vitro Growth of M. abscessus CIP 104536 T
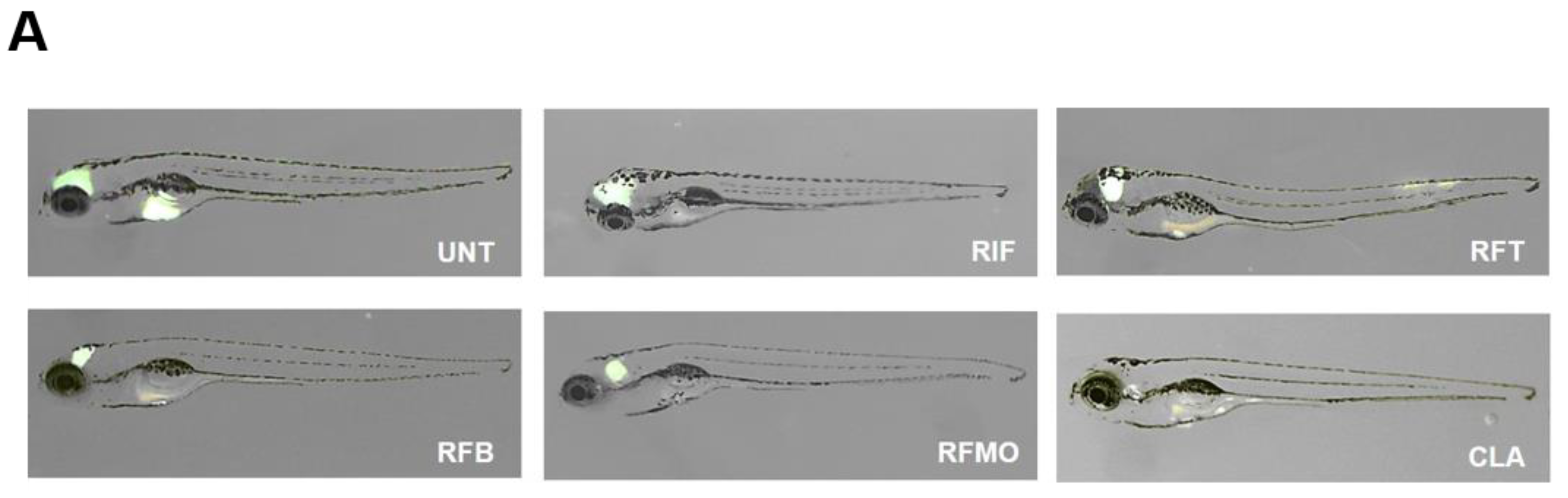
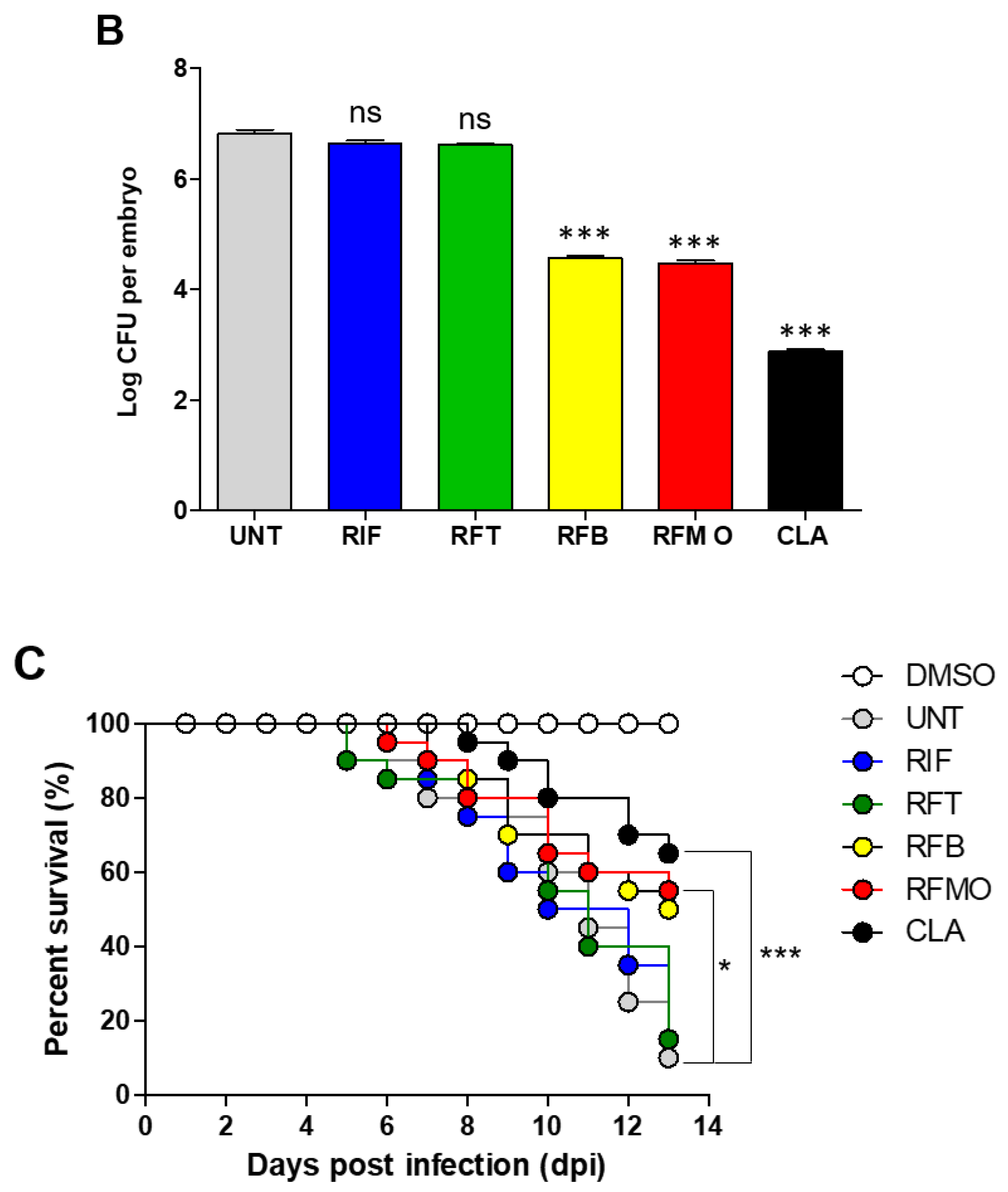
2.3. In Vivo Rifamycin O Efficacy Assessment Using ZF Embryo
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Bacterial Strains and Growing Conditions
4.3. Bioluminescent Reporter-Based Screening and Determination of Minimum Inhibitory Concentrations
4.4. Chemical Analysis
4.5. Cytotoxicity Assay
4.6. Drug Efficacy Assessment in M. Abscessus-infected ZF and the Use of Ziehl-Neelsen Staining
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, M.R.; Sheng, W.H.; Hung, C.C.; Yu, C.J.; Lee, L.N.; Hsueh, P.R. Mycobacterium abscessus complex infections in humans. Emerg. Infect. Dis. 2015, 21, 1638–1646. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.; Byrd, T.F. Mycobacterium abscessus: Shapeshifter of the Mycobacterial World. Front. Microbiol. 2018, 9, 2642. [Google Scholar] [CrossRef] [PubMed]
- Mougari, F.; Bouziane, F.; Crockett, F.; Nessar, R.; Chau, F.; Veziris, N.; Sapriel, G.; Raskine, L.; Cambau, E. Selection of Resistance to Clarithromycin in Mycobacterium abscessus Subspecies. Antimicrob. Agents Chemother. 2017, 61, e00943-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nessar, R.; Cambau, E.; Reyrat, J.M.; Murray, A.; Gicquel, B. Mycobacterium abscessus: A new antibiotic nightmare. J. Antimicrob. Chemother. 2012, 67, 810–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, J.M.; Grogono, D.M.; Greaves, D.; Foweraker, J.; Roddick, I.; Inns, T.; Reacher, M.; Haworth, C.S.; Curran, M.D.; Harris, S.R.; et al. Whole-genome sequencing to identify transmission of Mycobacterium abscessus between patients with cystic fibrosis: A retrospective cohort study. Lancet 2013, 381, 1551–1560. [Google Scholar] [CrossRef] [Green Version]
- Brown-Elliott, B.A.; Vasireddy, S.; Vasireddy, R.; Iakhiaeva, E.; Howard, S.T.; Nash, K.; Parodi, N.; Strong, A.; Gee, M.; Smith, T.; et al. Utility of Sequencing the erm (41) Gene in Isolates of Mycobacterium abscessus subsp. abscessus with Low and Intermediate Clarithromycin MICs. J. Clin. Microbiol. 2015, 53, 1211–1215. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.T. Rifamycin resistance in mycobacteria. Res. Microbiol. 1996, 147, 48–52. [Google Scholar] [CrossRef]
- Rominski, A.; Roditscheff, A.; Selchow, P.; Böttger, E.C.; Sander, P. Intrinsic rifamycin resistance of Mycobacterium abscessus is mediated by ADP-ribosyltransferase MAB_0591. J. Antimicrob. Chemother. 2017, 72, 376–384. [Google Scholar] [CrossRef] [Green Version]
- Dick, T.; Shin, S.J.; Koh, W.J.; Dartois, V.; Gengenbacher, M. Rifabutin is active against mycobacterium abscessus in mice. Antimicrob. Agents Chemother. 2020, 64, 2019–2021. [Google Scholar] [CrossRef] [Green Version]
- Aziz, D.B.; Low, J.L.; Wu, M.-L.; Gengenbacher, M.; Teo, J.W.P.; Dartois, V.; Dick, T. Rifabutin Is Active against Mycobacterium abscessus Complex. Antimicrob. Agents Chemother. 2017, 61, e00155-17. [Google Scholar] [CrossRef] [Green Version]
- Ganapathy, U.S.; Dartois, V.; Dick, T. Repositioning rifamycins for Mycobacterium abscessus lung disease. Expert Opin. Drug Discov. 2019, 14, 867–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernut, A.; Herrmann, J.-L.; Ordway, D.; Kremer, L. The Diverse Cellular and Animal Models to Decipher the Physiopathological Traits of Mycobacterium abscessus Infection. Front. Cell. Infect. Microbiol. 2017, 7, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacchi, A.; Pelizzi, G.; Nebuloni, M.; Ferrari, P. Comprehensive Study on Structure–Activity Relationships of Rifamycins: Discussion of Molecular and Crystal Structure and Spectroscopic and Thermochemical Properties of Rifamycin O. J. Med. Chem. 1998, 41, 2319–2332. [Google Scholar] [CrossRef]
- Combrink, K.D.; Ramos, A.R.; Spring, S.; Schmidl, S.; Elizondo, K.; Morin, P.; De Jesus, B.; Maurer, F.P. Rifamycin derivatives active against pathogenic rapidly-growing mycobacteria. Bioorg. Med. Chem. Lett. 2019, 29, 2112–2115. [Google Scholar] [CrossRef] [PubMed]
- De Groote, M.A.; Johnson, L.; Podell, B.; Brooks, E.; Basaraba, R.; Gonzalez-Juarrero, M. GM-CSF knockout mice for preclinical testing of agents with antimicrobial activity against Mycobacterium abscessus. J. Antimicrob. Chemother. 2014, 69, 1057–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combrink, K.D.; Denton, D.A.; Harran, S.; Ma, Z.; Chapo, K.; Yan, D.; Bonventre, E.; Roche, E.D.; Doyle, T.B.; Robertson, G.T.; et al. New C25 carbamate rifamycin derivatives are resistant to inactivation by ADP-ribosyl transferases. Bioorg. Med. Chem. Lett. 2007, 17, 522–526. [Google Scholar] [CrossRef]
- Obregón-Henao, A.; Arnett, K.A.; Henao-Tamayo, M.; Massoudi, L.; Creissen, E.; Andries, K.; Lenaerts, A.J.; Ordway, D.J. Susceptibility of Mycobacterium abscessus to Antimycobacterial Drugs in Preclinical Models. Antimicrob. Agents Chemother. 2015, 59, 6904–6912. [Google Scholar] [CrossRef] [Green Version]
- Meeker, N.D.; Trede, N.S. Immunology and zebrafish: Spawning new models of human disease. Dev. Comp. Immunol. 2008. [CrossRef]
- Bernut, A.; Lutfalla, G.; Kremer, L. Regard à travers le danio pour mieux comprendre les interactions hôte/pathogène. Medecine/Sciences 2015, 31, 638–646. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Netherton, M.; Byrd, T.F.; Rohde, K.H. Reporter-Based Assays for High-Throughput Drug Screening against Mycobacterium abscessus. Front. Microbiol. 2017, 8, 2204. [Google Scholar] [CrossRef]
- Kim, T.H.; Hanh, B.T.B.; Kim, G.; Lee, D.G.; Park, J.W.; Lee, S.E.; Kim, J.S.; Kim, B.S.; Ryoo, S.; Jo, E.K.; et al. Thiostrepton: A Novel Therapeutic Drug Candidate for Mycobacterium abscessus Infection. Molecules 2019, 24, 4511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.; Kim, G.; Moon, C.; Kim, H.J.; Kim, T.H.; Jang, J. Pathogen Box screening for hit identification against Mycobacterium abscessus. PLoS ONE 2018, 13, e0195595. [Google Scholar] [CrossRef] [PubMed]
- Ramis, I.B.; Cnockaert, M.; von Groll, A.; Nogueira, C.L.; Leão, S.C.; Andre, E.; Simon, A.; Palomino, J.C.; da Silva, P.E.A.; Vandamme, P.; et al. Antimicrobial susceptibility of rapidly growing mycobacteria using the rapid colorimetric method. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1403–1413. [Google Scholar] [CrossRef]
- Takaki, K.; Davis, J.M.; Winglee, K.; Ramakrishnan, L. Evaluation of the pathogenesis and treatment of Mycobacterium marinum infection in zebrafish. Nat. Protoc. 2013, 8, 1114–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC90 (μM) | ||||
|---|---|---|---|---|---|
| RFM O | RFB | RIF | RFT | CLR | |
| M. abscessus subsp. abscessus CIP104536T | 6.2 | 4.0 | >50 | >50 | 1.4 |
| M. abscessus subsp. bolletii CIP108541T | 4.0 | 7.1 | >50 | 46.6 | 1.5 |
| M. abscessus subsp. massiliense CIP108297T | 5.6 | 4.5 | 47.3 | >50 | 0.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanh, B.T.B.; Park, J.-W.; Kim, T.H.; Kim, J.-S.; Yang, C.-S.; Jang, K.; Cui, J.; Oh, D.-C.; Jang, J. Rifamycin O, An Alternative Anti-Mycobacterium abscessus Agent. Molecules 2020, 25, 1597. https://doi.org/10.3390/molecules25071597
Hanh BTB, Park J-W, Kim TH, Kim J-S, Yang C-S, Jang K, Cui J, Oh D-C, Jang J. Rifamycin O, An Alternative Anti-Mycobacterium abscessus Agent. Molecules. 2020; 25(7):1597. https://doi.org/10.3390/molecules25071597
Chicago/Turabian StyleHanh, Bui Thi Bich, June-Woo Park, Tae Ho Kim, Jae-Sung Kim, Chul-Su Yang, Kiseok Jang, Jinsheng Cui, Dong-Chan Oh, and Jichan Jang. 2020. "Rifamycin O, An Alternative Anti-Mycobacterium abscessus Agent" Molecules 25, no. 7: 1597. https://doi.org/10.3390/molecules25071597
APA StyleHanh, B. T. B., Park, J. -W., Kim, T. H., Kim, J. -S., Yang, C. -S., Jang, K., Cui, J., Oh, D. -C., & Jang, J. (2020). Rifamycin O, An Alternative Anti-Mycobacterium abscessus Agent. Molecules, 25(7), 1597. https://doi.org/10.3390/molecules25071597