
Lactobacillus-Derived Bioactive Metabolites for the Regulation of Periodontal Health: Evidences to Clinical Setting


Abstract
:1. Introduction
2. Lactobacillus-Derived Bioactive Metabolites
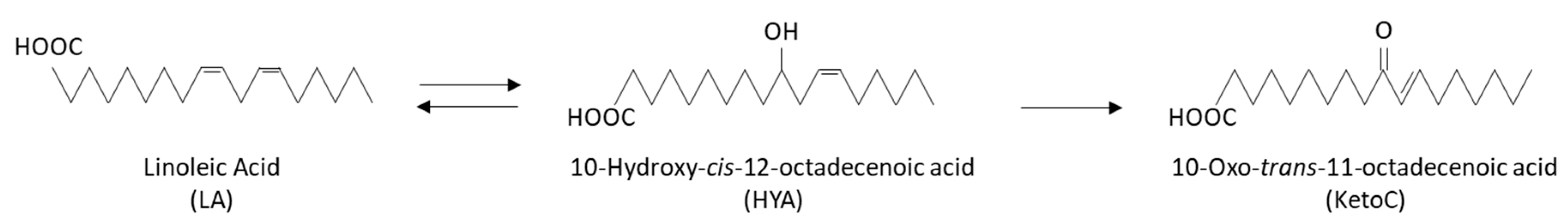
2.1. KetoC and HYA: Generation of Bioactive Metabolites by Lactobacillus
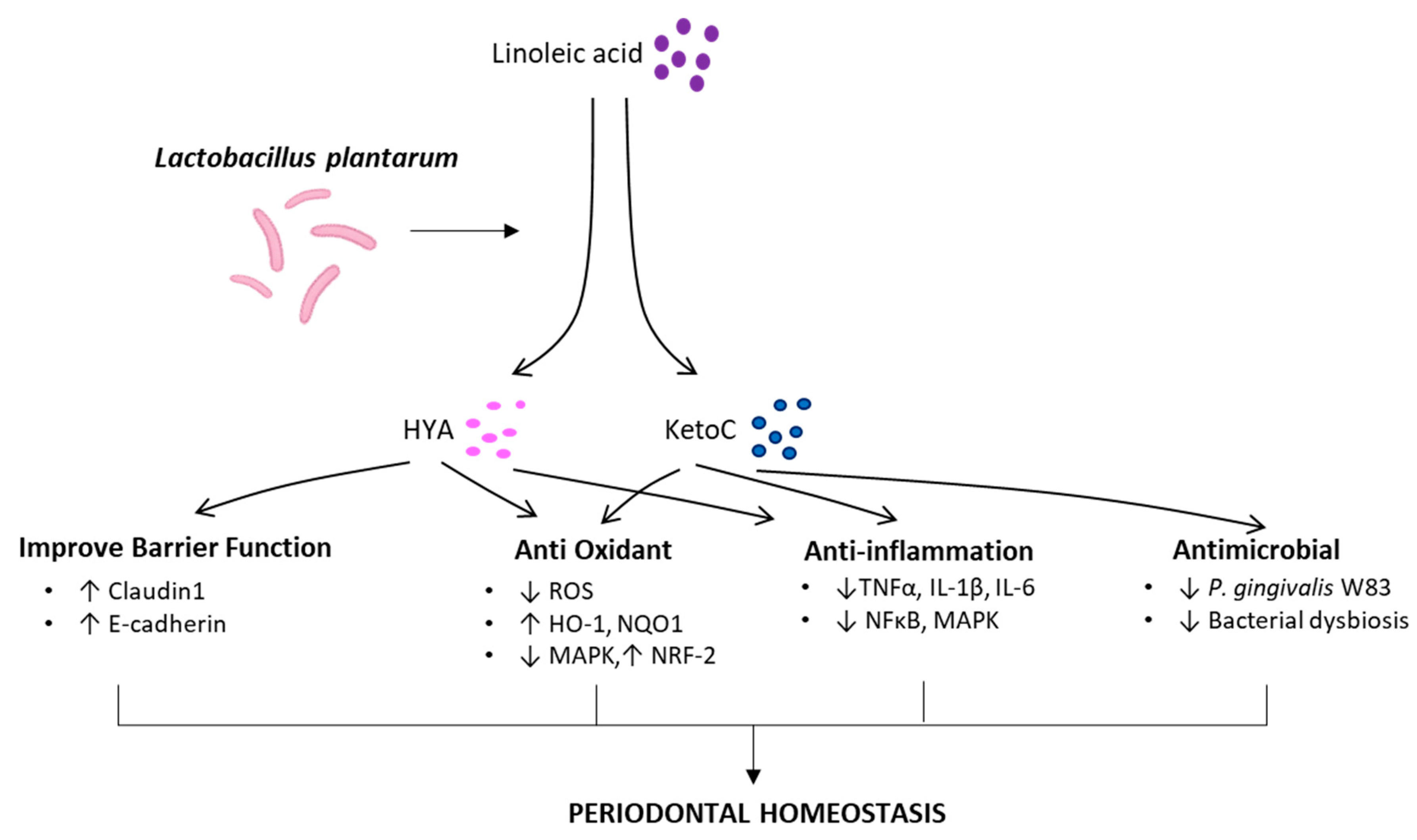
2.2. Beneficial Functions of KetoC and HYA in the Physiological and Pathological Processes
2.2.1. Anti-Inflammatory Function
2.2.2. Antimicrobial Function
2.2.3. Effect on Epithelial Barrier Function
2.2.4. Anti-Oxidant Function and Other
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scotti, E.; Boué, S.; Sasso, G.; Lo Zanetti, F.; Belcastro, V.; Poussin, C.; Sierro, N.; Battey, J.; Gimalac, A.; Ivanov, N.V.; et al. Exploring the microbiome in health and disease. Toxicol. Res. Appl. 2017, 1. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Eberl, G.; Lochner, M. The development of intestinal lymphoid tissues at the interface of self and microbiota. Mucosal Immunol. 2009, 2, 478–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; De los Reyes-Gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-Induced Obesity Is Linked to Marked but Reversible Alterations in the Mouse Distal Gut Microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Koopen, A.M.; Groen, A.K.; Nieuwdorp, M. Human microbiome as therapeutic intervention target to reduce cardiovascular disease risk. Curr. Opin. Lipidol. 2016, 27, 615–622. [Google Scholar] [CrossRef]
- Kato, I.; Vasquez, A.; Moyerbrailean, G.; Land, S.; Djuric, Z.; Sun, J.; Lin, H.-S.; Ram, J.L. Nutritional Correlates of Human Oral Microbiome. J. Am. Coll. Nutr. 2017, 36, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.M.; Gleber-Netto, F.O.; Fernandes, G.R.; Amorim, M.; Barbosa, L.F.; Francisco, A.L.N.; Andrade, A.G.; Setubal, J.C.; Kowalski, L.P.; Nunes, D.N.; et al. Alcohol and tobacco consumption affects bacterial richness in oral cavity mucosa biofilms. BMC Microbiol. 2014, 14, 250. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, K. The Human Microbiota and Chronic Disease. In The Human Microbiota and Chronic Disease: Dysbiosis as a Cause of Human Pathology; Nibali, L., Henderson, B., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; Available online: http://doi.wiley.com/10.1002/9781118982907 (accessed on 14 January 2020).
- Papageorgiou, S.N.; Hagner, M.; Nogueira, A.V.B.; Franke, A.; Jäger, A.; Deschner, J. Inflammatory bowel disease and oral health: Systematic review and a meta-analysis. J. Clin. Periodontol. 2017, 44, 382–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habashneh, R.A.; Khader, Y.S.; Alhumouz, M.K.; Jadallah, K.; Ajlouni, Y. The association between inflammatory bowel disease and periodontitis among Jordanians: A case-control study. J. Periodontal Res. 2012, 47, 293–298. [Google Scholar] [CrossRef]
- Lira-Junior, R.; Figueredo, C.M. Periodontal and inflammatory bowel diseases: Is there evidence of complex pathogenic interactions? World J. Gastroenterol. 2016, 22, 7963–7972. [Google Scholar] [CrossRef]
- Takahashi, N.; Sulijaya, B.; Yamada-Hara, M.; Tsuzuno, T.; Tabeta, K.; Yamazaki, K. Gingival epithelial barrier: Regulation by beneficial and harmful microbes. Tissue Barriers 2019, 7, e1651158-2. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Arimatsu, K.; Kato, T.; Matsuda, Y.; Minagawa, T.; Takahashi, N.; Ohno, H.; Yamazaki, K. Oral administration of P. gingivalis induces dysbiosis of gut microbiota and impaired barrier function leading to dissemination of enterobacteria to the liver. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulijaya, B.; Takahashi, N.; Yamazaki, K.; Yamazaki, K. Nutrition as Adjunct Therapy in Periodontal Disease Management. Curr. Oral Health Rep. 2019, 6, 61–69. [Google Scholar] [CrossRef]
- Sulijaya, B.; Takahashi, N.; Yamazaki, K. Host modulation therapy using anti-inflammatory and antioxidant agents in periodontitis: A review to a clinical translation. Arch. Oral Biol. 2019, 105, 72–80. [Google Scholar] [CrossRef]
- Sulijaya, B.; Yamada-Hara, M.; Yokoji-Takeuchi, M.; Matsuda-Matsukawa, Y.; Yamazaki, K.; Matsugishi, A.; Tsuzuno, T.; Sato, K.; Aoki-Nonaka, Y.; Takahashi, N.; et al. Antimicrobial function of the polyunsaturated fatty acid ketoC in an experimental model of periodontitis. J. Periodontol. 2019. [Google Scholar] [CrossRef]
- Sulijaya, B.; Takahashi, N.; Yamada, M.; Yokoji, M.; Sato, K.; Aoki-Nonaka, Y.; Nakajima, T.; Kishino, S.; Ogawa, J.; Yamazaki, K. The anti-inflammatory effect of 10-oxo-trans-11-octadecenoic acid (KetoC) on RAW 264.7 cells stimulated with Porphyromonas gingivalis lipopolysaccharide. J. Periodontal Res. 2018, 53, 777–784. [Google Scholar] [CrossRef]
- Yamada, M.; Takahashi, N.; Matsuda, Y.; Sato, K.; Yokoji, M.; Sulijaya, B.; Maekawa, T.; Ushiki, T.; Mikami, Y.; Hayatsu, M.; et al. A bacterial metabolite ameliorates periodontal pathogen-induced gingival epithelial barrier disruption via GPR40 signaling. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Miyamoto, J.; Mizukure, T.; Park, S.-B.; Kishino, S.; Kimura, I.; Hirano, K.; Bergamo, P.; Rossi, M.; Suzuki, T.; Arita, M.; et al. A Gut Microbial Metabolite of Linoleic Acid, 10-Hydroxy-cis-12-octadecenoic Acid, Ameliorates Intestinal Epithelial Barrier Impairment Partially via GPR40-MEK-ERK Pathway. J. Biol. Chem. 2014, 290, 2902–2918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanthirudjanar, T.; Furumoto, H.; Zheng, J.; Kim, Y.-I.; Goto, T.; Takahashi, N.; Kawada, T.; Park, S.-B.; Hirata, A.; Kitamura, N.; et al. Gut microbial fatty acid metabolits reduce triacyglycerol levels in hepatocytes. Lipids 2015, 50, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Furumoto, H.; Nanthirudjanar, T.; Kume, T.; Izumi, Y.; Park, S.B.; Kitamura, N.; Kishino, S.; Ogawa, J.; Hirataa, T.; Sugawara, T. 10-Oxo-trans-11-octadecenoic acid generated from linoleic acid by a gut lactic acid bacterium Lactobacillus plantarum is cytoprotective against oxidative stress. Toxicol. Appl. Pharmacol. 2016, 296, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.E.; Li, Y.; Nishimura, A.; Jheng, H.F.; Yuliana, A.; Kitano-Ohue, R.; Nomura, W.; Takahashi, N.; Kim, C.S.; Yu, R.; et al. Synthesized enone fatty acids resembling metabolites from gut microbiota suppress macrophage-mediated inflammation in adipocytes. Mol. Nutr. Food Res. 2017, 61, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Furuzono, T.; Yamakuni, K.; Li, Y.; Kim, Y.I.; Takahashi, H.; Ohue-Kitano, R.; Jheng, H.F.; Takahashi, N.; Kano, Y.; et al. 10-oxo-12(Z)-octadecenoic acid, a linoleic acid metabolite produced by gut lactic acid bacteria, enhances energy metabolism by activation of TRPV1. FASEB J. 2017, 31, 5036–5048. [Google Scholar] [CrossRef] [Green Version]
- Kishino, S.; Takeuchi, M.; Park, S.-B.; Hirata, A.; Kitamura, N.; Kunisawa, J.; Kiyono, H.; Iwamoto, R.; Isobe, Y.; Arita, M.; et al. Polyunsaturated fatty acid saturation by gut lactic acid bacteria affecting host lipid composition. Proc. Natl. Acad. Sci. USA 2013, 110, 17808–17813. [Google Scholar] [CrossRef] [Green Version]
- Ha, Y.L.; Grimm, N.K.; Pariza, M.W. Anticarcinogens from fried ground beef: Heat-altered derivatives of linoleic acid. Carcinogenesis 1987, 8, 1881–1887. [Google Scholar] [CrossRef]
- Ip, C.; Singh, M.; Thompson, H.J.; Scimeca, J.A. Conjugated Linoleic Acid Suppresses Mammary Carcinogenesis and Proliferative Activity of the Mammary Gland in the Rat. Cancer Res. 1994, 54, 1212–1215. [Google Scholar]
- Cook, M.E.; Miller, C.C.; Park, Y.; Pariza, M. Immune modulation by altered nutrient metabolism: Nutritional control of immune-induced growth depression. Poult. Sci. 1993, 72, 1301–1305. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.C.; Park, Y.; Pariza, M.W.; Cook, M.E. Feeding conjugated linoleic acid to animals partially overcomes catabolic responses due to endotoxin injection. Biochem. Biophys. Res. Commun. 1994, 198, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Polan, C.E.; Mc Neill, J.J.; Tove, S.B. Biohydrogenation of Unsaturated Fatty Acids by Rumen Bacteria. J. Bacteriol. 1964, 88, 1056–1064. [Google Scholar] [CrossRef] [Green Version]
- Hou, F.; Miyakawa, T.; Kitamura, N.; Takeuchi, M.; Park, S.B.; Kishino, S.; Ogawa, J.; Tanokura, M. Structure and reaction mechanism of a novel enone reductase. FEBS J. 2015, 282, 1526–1537. [Google Scholar] [CrossRef] [Green Version]
- Kishino, S.; Ogawa, J.; Omura, Y.; Matsumura, K.; Shimizu, S. Conjugated linoleic acid production from linoleic acid by lactic acid bacteria. JAOCS J. Am. Oil Chem. Soc. 2002, 79, 159–163. [Google Scholar] [CrossRef]
- Ogawa, J.; Kishino, S.; Ando, A.; Sugimoto, S.; Mihara, K.; Shimizu, S. Production of conjugated fatty acids by lactic acid bacteria. J. Biosci. Bioeng. 2005, 100, 355–364. [Google Scholar] [CrossRef]
- Kaikiri, H.; Miyamoto, J.; Kawakami, T.; Park, S.B.; Kitamura, N.; Kishino, S.; Yonejima, Y.; Hisa, K.; Watanabe, J.; Ogita, T.; et al. Supplemental feeding of a gut microbial metabolite of linoleic acid, 10-hydroxy-cis-12-octadecenoic acid, alleviates spontaneous atopic dermatitis and modulates intestinal microbiota in NC/nga mice. Int. J. Food Sci. Nutr. 2017, 68, 941–951. [Google Scholar] [CrossRef]
- Ikeguchi, S.; Izumi, Y.; Kitamura, N.; Kishino, S.; Ogawa, J.; Akaike, A.; Kume, T. Inhibitory effect of the gut microbial linoleic acid metabolites, 10-oxo-trans-11-octadecenoic acid and 10-hydroxy-cis-12-octadecenoic acid, on BV-2 microglial cell activation. J. Pharmacol. Sci. 2018, 138, 9–15. [Google Scholar] [CrossRef]
- Tomofuji, T.; Irie, K.; Sanbe, T.; Azuma, T.; Ekuni, D.; Tamaki, N.; Yamamoto, T.; Morita, M. Periodontitis and increase in circulating oxidative stress. Jpn. Dent. Sci. Rev. 2009, 45, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Sofyana, N.T.; Zheng, J.; Manabe, Y.; Yamamoto, Y.; Kishino, S.; Ogawa, J.; Sugawara, T. Gut microbial fatty acid metabolites (KetoA and KetoC) affect the progression of nonalcoholic steatohepatitis and reverse cholesterol transport metabolism in mouse model. Lipids 2020, 55. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef]
- Itoh, Y.; Hinuma, S. GPR40, a free fatty acid receptor on pancreatic β cells, regulates insulin secretion. Hepatol. Res. 2005, 33, 171–173. [Google Scholar] [CrossRef]
- Miyamoto, J.; Hasegawa, S.; Kasubuchi, M.; Ichimura, A.; Nakajima, A.; Kimura, I. Nutritional signaling via free fatty acid receptors. Int. J. Mol. Sci. 2016, 17, 450. [Google Scholar] [CrossRef]
- Tazoe, H.; Otomo, Y.; Kaji, I.; Tanaka, R.; Karaki, S.I.; Kuwahara, A. Roles of short-chain fatty acids receptors, GPR41 and GPR43 on colonic functions. J. Physiol. Pharmacol. 2008, 59 (Suppl. 2), 251–262. [Google Scholar]
- Tazoe, H.; Otomo, Y.; Karaki, S.I.; Kato, I.; Fukami, Y.; Terasaki, M.; Kuwahara, A. Expression of short-chain fatty acid receptor GPR41 in the human colon. Biomed. Res. 2009, 30, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briscoe, C.P.; Tadayyon, M.; Andrews, J.L.; Benson, W.G.; Chambers, J.K.; Eilert, M.M.; Ellis, C.; Elshourbagy, N.A.; Goetz, A.S.; Minnick, D.T.; et al. The orphan G protein-coupled receptor GPR40 is activated by medium and long chain fatty acids. J. Biol. Chem. 2003, 278, 11303–11311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, T.; Hirasawa, A.; Ichimura, A.; Kimura, I.; Tsujimoto, G. Free fatty acid receptors FFAR1 and GPR120 as novel therapeutic targets for metabolic disorders. J. Pharm. Sci. 2011, 100, 3594–3601. [Google Scholar] [CrossRef] [PubMed]
- Saba, F.; Sirigu, A.; Pillai, R.; Caria, P.; Cordeddu, L.; Carta, G.; Murru, E.; Sogos, V.; Banni, S. Downregulation of inflammatory markers by conjugated linoleic acid isomers in human cultured astrocytes. Nutr. Neurosci. 2019, 22, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Cestari, J.A.F.; Fabri, G.M.C.; Kalil, J.; Nitrini, R.; Jacob-Filho, W.; De Siqueira, J.T.T.; Siqueira, S.R. Oral Infections and Cytokine Levels in Patients with Alzheimer’s Disease and Mild Cognitive Impairment Compared with Controls. J. Alzheimer’s Dis. 2016, 52, 1479–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, J.M.; Scarmeas, N.; Celenti, R.S.; Elkind, M.S.V.; Wright, C.B.; Schupf, N.; Papapanou, P.N. Serum IgG antibody levels to periodontal microbiota are associated with incident alzheimer disease. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.B.; Saito, M.T.; Matheus, F.C.; Prediger, R.D.; Yamada, E.S.; Maia, C.S.F.; Lima, R.R. Periodontitis and alzheimer’s disease: A possible comorbidity between oral chronic inflammatory condition and neuroinflammation. Front. Aging Neurosci. 2017, 10, 327. [Google Scholar] [CrossRef]
- Sun, M.; Dong, J.; Xia, Y.; Shu, R. Antibacterial activities of docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) against planktonic and biofilm growing Streptococcus mutans. Microb. Pathog. 2017, 107, 212–218. [Google Scholar] [CrossRef]
- Correia, M.; Michel, V.; Matos, A.A.; Carvalho, P.; Oliveira, M.J.; Ferreira, R.M.; Dillies, M.A.; Huerre, M.; Seruca, R.; Figueiredo, C.; et al. Docosahexaenoic acid inhibits Helicobacter pylori growth in vitro and mice gastric mucosa colonization. PLoS ONE 2012, 7, e35072. [Google Scholar] [CrossRef]
- Thompson, L.; Cockayne, A.; Spiller, R.C. Inhibitory effect of polyunsaturated fatty acids on the growth of Helicobacter pylori: A possible explanation of the effect of diet on peptic ulceration. Gut 1994, 35, 1557–1561. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.B.; Ebersole, J.L. A novel bioactivity of omega-3 polyunsaturated fatty acids and their ester derivatives. Mol. Oral Microbiol. 2010, 25, 75–80. [Google Scholar] [CrossRef]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An overview of periodontopathic pathogen below the gum line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhou, Z.; Dong, J.; Zhang, J.; Xia, Y.; Shu, R. Antibacterial and antibiofilm activities of docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) against periodontopathic bacteria. Microb. Pathog. 2016, 99, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Bergsson, G.; Steingrímsson, Ó.; Thormar, H. Bactericidal effects of fatty acids and monoglycerides on Helicobacter pylori. Int. J. Antimicrob. Agents. 2002, 20, 258–262. [Google Scholar] [CrossRef]
- DiRienzo, J.M. Breaking the Gingival Epithelial Barrier: Role of the Aggregatibacter actinomycetemcomitans Cytolethal Distending Toxin in Oral Infectious Disease. Cells 2014, 3, 476–499. [Google Scholar] [CrossRef]
- Brooke, M.A.; Nitoiu, D.; Kelsell, D.P. Cell-cell connectivity: Desmosomes and disease. J. Pathol. 2012, 226, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Ostaff, M.J.; Stange, E.F.; Wehkamp, J. Antimicrobial peptides and gut microbiota in homeostasis and pathology. EMBO Mol. Med. 2013, 5, 1465–1483. [Google Scholar] [CrossRef] [PubMed]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K. The Roles of Antimicrobial Peptides in Innate Host Defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulijaya, B.; Masulili, S.R.I.L.C.; Lessang, R.; Soeroso, Y.; Auerkari, E.I. The Human Beta-Defensin-1 Level from Smokers with Chronic Periodontitis. Asian J. Pharm. Clin. Res. 2016, 9, 174–176. [Google Scholar]
- Amano, A.; Takeuchi, H.; Furuta, N. Outer membrane vesicles function as offensive weapons in host-parasite interactions. Microbes Infect. 2010, 12, 791–798. [Google Scholar] [CrossRef]
- Nagano, K.; Hasegawa, Y.; Yoshida, Y.; Yoshimura, F. A major fimbrilin variant of Mfa1 fimbriae in Porphyromonas gingivalis. J. Dent. Res. 2015, 94, 1143–1148. [Google Scholar] [CrossRef]
- Takahashi, N.; Honda, T.; Domon, H.; Nakajima, T.; Tabeta, K.; Yamazaki, K. Interleukin-1 receptor-associated kinase-M in gingival epithelial cells attenuates the inflammatory response elicited by Porphyromonas gingivalis. J. Periodontal Res. 2010, 45, 512–519. [Google Scholar]
- Ogawa, T.; Yagi, T. Bioactive mechanism of Porphyromonas gingivalis lipid A. Periodontology 2000 2010, 54, 71–77. [Google Scholar] [CrossRef]
- Laine, M.L.; Appelmelk, B.J.; Van Winkelhoff, A.J. Prevalence and distribution of six capsular serotypes of Porphyromonas gingivalis in periodontitis patients. J. Dent. Res. 1997, 76, 1840–1844. [Google Scholar] [CrossRef]
- Singh, A.; Wyant, T.; Anaya-Bergman, C.; Aduse-Opoku, J.; Brunner, J.; Laine, M.L.; Curtis, M.A.; Lewis, J.P. The capsule of Porphyromonas gingivalis leads to a reduction in the host inflammatory response, evasion of phagocytosis, and increase in Virulence. Infect. Immun. 2011, 79, 4533–4542. [Google Scholar] [CrossRef] [Green Version]
- Potempa, J.; Banbula, A.; Travis, J. Role of bacterial proteinases in matrix destruction and modulation of host responses. Periodontology 2000 2000, 24, 153–192. [Google Scholar] [CrossRef] [PubMed]
- De Diego, I.; Veillard, F.; Sztukowska, M.N.; Guevara, T.; Potempa, B.; Pomowski, A.; Huntington, J.A.; Potempa, J.; Gomis-Rüth, F.X. Structure and mechanism of cysteine peptidase gingipain K (Kgp), a major virulence factor of Porphyromonas gingivalis. J. Biol. Chem. 2014, 289, 32291–32302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moens, E.; Veldhoen, M. Epithelial barrier biology: Good fences make good neighbours. Immunology 2012, 135, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Liu, Z.; Niu, B.; Zhang, J.; Tan, T.K.; Lee, S.R.; Zhao, Y.; Harris, D.C.; Zheng, G. E-Cadherin/β-catenin complex and the epithelial barrier. J. Biomed. Biotechnol. 2011, 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, P.; Chapple, C.C.; Kumar, R.K.; Hunter, N. Expression patterns of E-cadherin, involucrin, and connexin gap junction proteins in the lining epithelia of inflamed gingiva. J. Pathol. 2000, 192, 58–66. [Google Scholar] [CrossRef]
- Nagarakanti, S.; Ramya, S.; Babu, P.; Arun, K.V.; Sudarsan, S. Differential Expression of E-Cadherin and Cytokeratin 19 and Net Proliferative Rate of Gingival Keratinocytes in Oral Epithelium in Periodontal Health and Disease. J. Periodontol. 2007, 78, 2197–2202. [Google Scholar] [CrossRef]
- Chapple, I.L.C.; Brock, G.R.; Milward, M.R.; Ling, N.; Matthews, J.B. Compromised GCF total antioxidant capacity in periodontitis: Cause or effect? J. Clin. Periodontol. 2007, 34, 103–110. [Google Scholar] [CrossRef]
- Bullon, P. New Theories and Their Clinical Relevance to the Onset and Development of Periodontal Diseases. In Studies on Periodontal Disease, Oxidative Stress in Applied Basic Research and Clinical Practice; Springer: Berlin/Heidelberg, Germany, 2014; pp. 227–249. [Google Scholar]
- Varela-López, A.; Battino, M.; Bullón, P.; Quiles, J.L. Dietary antioxidants for chronic periodontitis prevention and its treatment. A review on current evidences from animal and human studies. ARS Pharm. 2015, 56, 131–140. [Google Scholar] [CrossRef]
- Wang, Y.; Andrukhov, O.; Rausch-Fan, X. Oxidative stress and antioxidant system in periodontitis. Front. Physiol. 2017, 8, 910. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.; Kim, H.J.; Kim, H.M. RhoA-JNK Regulates the E-Cadherin Junctions of Human Gingival Epithelial Cells. J. Dent. Res. 2016, 95, 284–291. [Google Scholar] [CrossRef]
- Yokoji-Takeuchi, M.; Takahashi, N.; Yamada-Hara, M.; Sulijaya, B.; Tsuzuno, T.; Aoki-Nonaka, Y.; Tabeta, K.; Kishino, S.; Ogawa, J.; Yamazaki, K. A bacterial metabolite induces Nrf2-mediated anti-oxidative responses in gingival epithelial cells by activating the MAPK signaling pathway. Arch. Oral Biol. 2020, 110, 104602. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Takagi, T.; Uchiyama, K.; Yoshikawa, T. Heme oxygenase-1: A novel therapeutic target for gastrointestinal diseases. J. Clin. Biochem. Nutr. 2011, 48, 126–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.Y.; Chen, Z.; Jasmer, K.J.; Chuang, D.Y.; Gu, Z.; Hannink, M.; Simonyi, A. Quercetin attenuates inflammatory responses in BV-2 microglial cells: Role of MAPKs on the Nrf2 pathway and induction of heme oxygenase-1. PLoS ONE 2015, 10, e0141509. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Dong, H.; Song, E.; Xu, X.; Liu, L.; Song, Y. Nrf2/ARE pathway activation, HO-1 and NQO1 induction by polychlorinated biphenyl quinone is associated with reactive oxygen species and PI3K/AKT signaling. Chem. Biol. Interact. 2014, 209, 56–67. [Google Scholar] [CrossRef]
- Park, J.Y.; Kang, K.A.; Kim, K.C.; Cha, J.W.; Kim, E.H.; Hyun, J.W. Morin Induces Heme Oxygenase-1 via ERK-Nrf2 Signaling Pathway. J. Cancer Prev. 2013, 18, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Alam, J.; Venkatesan, M.I.; Eiguren-Fernandez, A.; Schmitz, D.; Di Stefano, E.; Slaughter, N.; Killeen, E.; Wang, X.; Huang, A.; et al. Nrf2 Is a Key Transcription Factor that Regulates Antioxidant Defense in Macrophages and Epithelial Cells: Protecting against the Proinflammatory and Oxidizing Effects of Diesel Exhaust Chemicals. J. Immunol. 2004, 173, 3467–3481. [Google Scholar] [CrossRef] [PubMed]
- Michelotti, G.A.; Machado, M.V.; Diehl, A.M. NAFLD, NASH and liver cancer. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 656–665. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No | Author (Year) | Metabolite | Modality | Study | Outcome |
|---|---|---|---|---|---|
| 1 | Miyamoto et al. (2015) [24] | HYA | Improve epithelial barrier function | In vitro | HYA improves intestinal epithelial barrier impairment partially via GPR40-MEK-ERK pathway. |
| 2 | Furumoto et al. (2016) [26] | KetoC | Antioxidant | In vitro | KetoC increases antioxidant genes by upregulating the NRF2-ARE pathway in HepG2 cells. |
| 3 | Yang et al. (2017) [27] | KetoC | Anti-inflammatory | In vitro | KetoC exerts anti-inflammatory function via MAPK and NFκB signaling in macrophages induced with bacterial lipopolysaccharide. |
| 4 | Kaikiri et al. (2017) [38] | HYA | Anti-allergic and anti-inflammatory | In vivo | HYA feeding decreased TNF-α and increased claudin-1 (tight junction protein) levels in the mouse skin of atopic dermatitis (AD) model. |
| 5 | Yamada et al. (2018) [23] | HYA | Improve epithelial barrier function | In vivo | HYA tends to prevent alveolar bone loss in periodontitis model by improving the expression of E-cadherin in gingival tissue. |
| In vitro | HYA increases beta defensin, thereby inhibiting inflammation | ||||
| 6 | Ikeguchi et al. (2018) [39] | KetoC and HYA | Anti-inflammatory | In vitro | KetoC and HYA were found to inhibit ERK phosphorylation induced by LPS in microglial cells. |
| 7 | Sulijaya et al. (2018) [22] | KetoC | Anti-inflammatory | In vitro | GPR120 mediates the suppression function of KetoC towards TNFα in P. gingivalis LPS-induced inflammation through NfκB p65 pathway. |
| 8 | Sulijaya et al. (2019) [21] | KetoC | Antimicrobial | In vivo | Oral gavage of KetoC reduces alveolar bone loss in P. gingivalis W83-induced periodontitis mice model. |
| In vitro | KetoC inhibits P. gingivalis strain W83 growth in a dose-dependent manner. | ||||
| 9 | Takeuchi et al. (2020) [40] | KetoC | Antioxidant | In vitro | KetoC counters oxidative stress condition in gingival epithelial cells through GPR120-Nrf2 ARE-MAPK pathway. |
| 10 | Sofyana et al. (2020) [41] | KetoC | HDL modulator | In vivo | KetoC upregulates HDL related genes and HDL cholesterol levels in the plasma. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulijaya, B.; Takahashi, N.; Yamazaki, K. Lactobacillus-Derived Bioactive Metabolites for the Regulation of Periodontal Health: Evidences to Clinical Setting. Molecules 2020, 25, 2088. https://doi.org/10.3390/molecules25092088
Sulijaya B, Takahashi N, Yamazaki K. Lactobacillus-Derived Bioactive Metabolites for the Regulation of Periodontal Health: Evidences to Clinical Setting. Molecules. 2020; 25(9):2088. https://doi.org/10.3390/molecules25092088
Chicago/Turabian StyleSulijaya, Benso, Naoki Takahashi, and Kazuhisa Yamazaki. 2020. "Lactobacillus-Derived Bioactive Metabolites for the Regulation of Periodontal Health: Evidences to Clinical Setting" Molecules 25, no. 9: 2088. https://doi.org/10.3390/molecules25092088
APA StyleSulijaya, B., Takahashi, N., & Yamazaki, K. (2020). Lactobacillus-Derived Bioactive Metabolites for the Regulation of Periodontal Health: Evidences to Clinical Setting. Molecules, 25(9), 2088. https://doi.org/10.3390/molecules25092088