
A Chemometry of Aldrovanda vesiculosa L. (Waterwheel, Droseraceae) Populations

 ,
,  ,
,  ,
, 
 ,
, 

Abstract
:1. Introduction
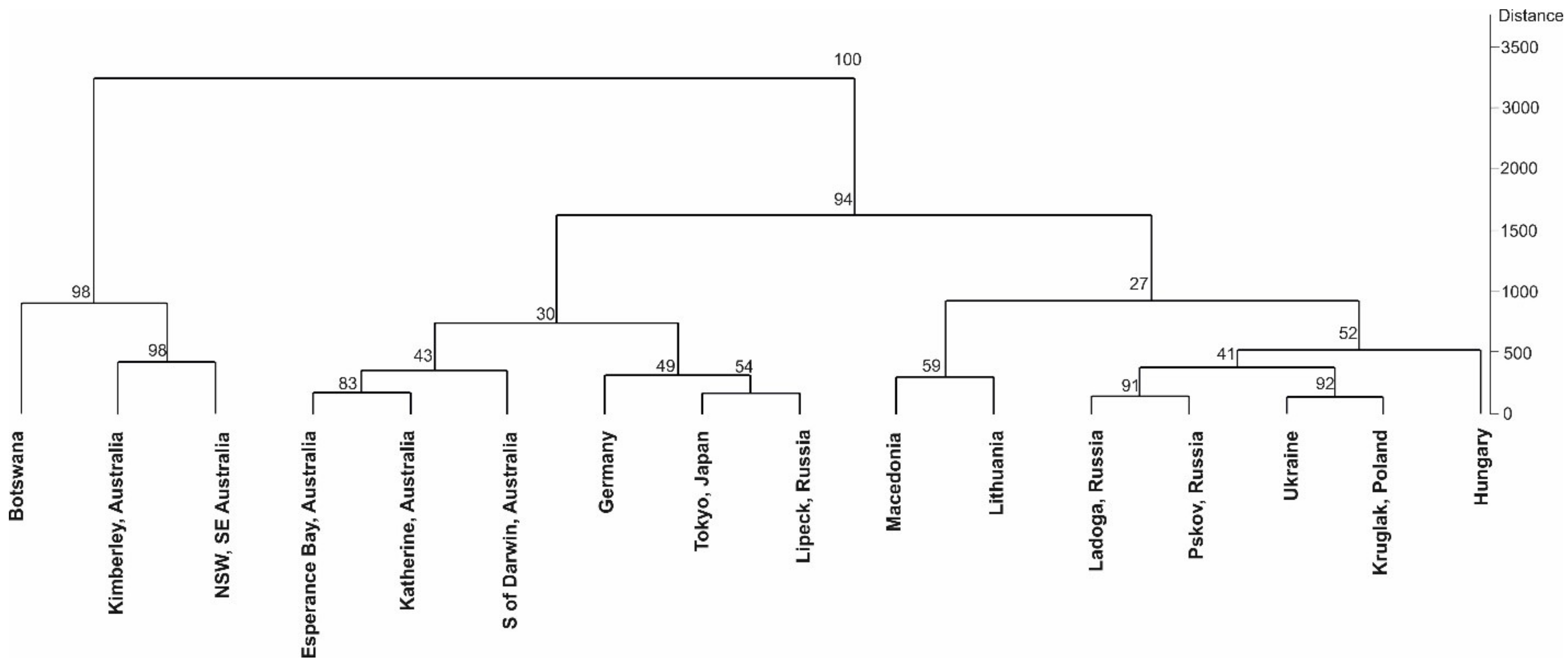
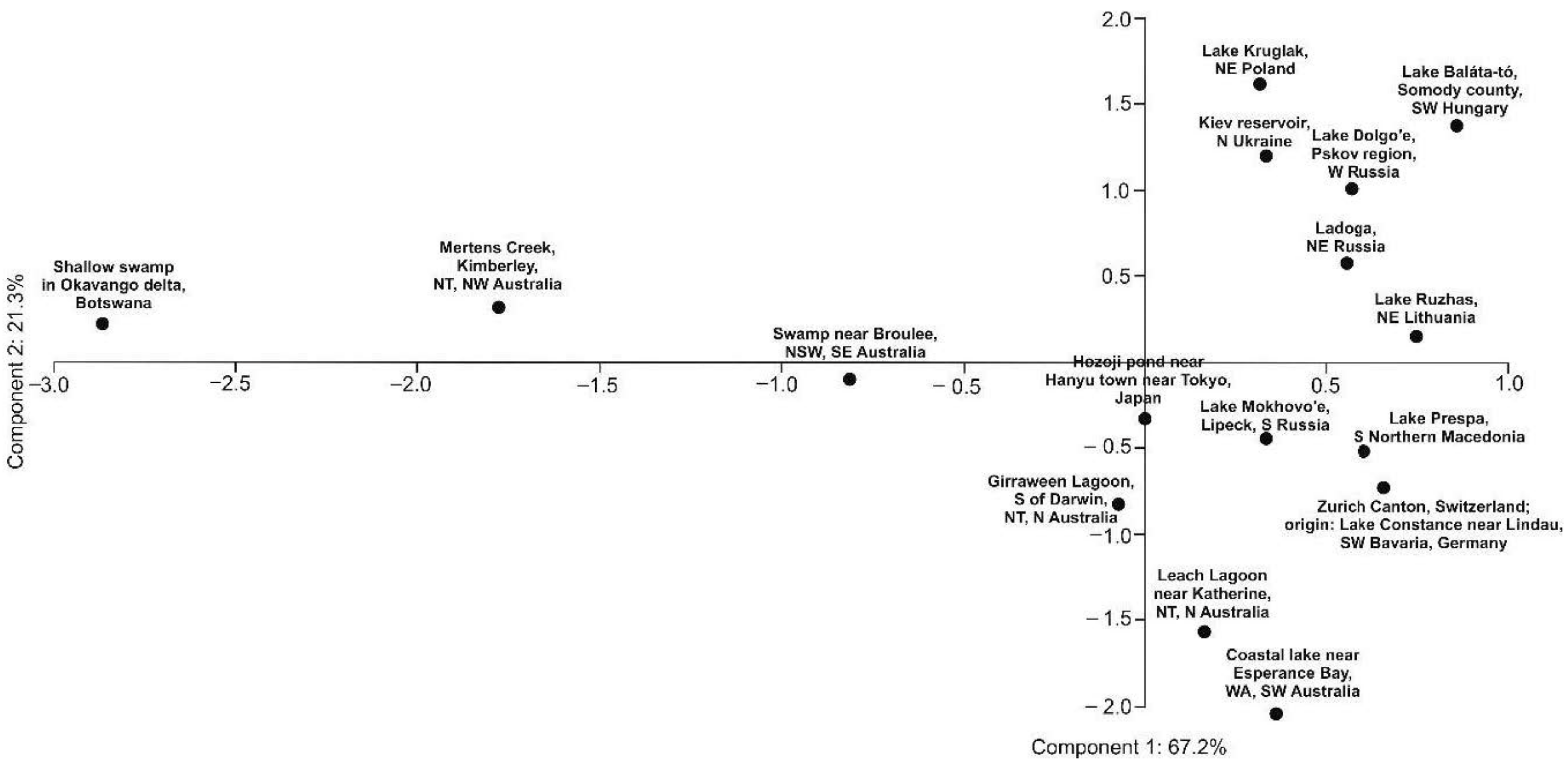
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Sample Preparation
4.3. Gas Chromatography Analysis
4.4. Preprocessing of Data and Chemometric Analysis
4.5. Mantel Test
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lloyd, F.E. The Carnivorous Plants; Verdoorn, F., Ed.; Waltham: New York, NY, USA, 1942. [Google Scholar]
- Król, E.; Płachno, B.J.; Adamec, L.; Stolarz, M.; Dziubińska, H.; Trebacz, K. Quite a few reasons for calling carnivores “the most wonderful plants in the world”. Ann. Bot. 2012, 109, 47–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, A. Aldrovanda: The Waterwheel Plant; Redfern Natural History Productions Ltd.: Poole, UK, 2012. [Google Scholar]
- Cameron, K.M.; Wurdack, K.J.; Jobson, R.W. Molecular evidence for the common origin of snap-traps among carnivorous plants. Am. J. Bot. 2002, 89, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Palfalvi, G.; Hackl, T.; Terhoeven, N.; Shibata, T.F.; Nishiyama, T.; Ankenbrand, M.; Becker, D.; Förster, F.; Freund, M.; Iosip, A.; et al. Genomes of the venus flytrap and close relatives unveil the roots of plant carnivory. Curr. Biol. 2020, 30, 2312–2320. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, A.; Schlauer, J.; Smith, S.A.; Givnish, T.J. Evolution of carnivory in angiosperms. In Carnivorous Plants: Physiology, Ecology, and Evolution; Elison, A., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; Chapter 3; pp. 22–42. ISBN 9780198779841. [Google Scholar]
- Yakubovskaya, T.V. The genus Aldrovanda (Droseraceae) in the Pleistocene of the Belorussian SSR. Bot. Zhurnal 1991, 76, 109–118. [Google Scholar]
- Adamec, L. Biological flora of Central Europe: Aldrovanda vesiculosa L. Perspect. Plant. Ecol. Evol. Syst. 2018, 35, 8–21. [Google Scholar] [CrossRef]
- Cross, A.; Adamec, L. Aldrovanda vesiculosa. The IUCN Red List of Threatened Species 2020: E.T162346A83998419. Available online: https://www.iucnredlist.org/ (accessed on 25 December 2020).
- Cross, A.T.; Krueger, T.A.; Gonella, P.M.; Robinson, A.S.; Fleischmann, A.S. Conservation of carnivorous plants in the age of extinction. Glob. Ecol. Conserv. 2020, 24, e01272. [Google Scholar] [CrossRef]
- Svobodová, I.; Adamec, L. Preliminary identification of the agent causing the fungal disease of Aldrovanda vesiculosa. Carniv. Plant. Newsl. 2020, 49, 56–64. [Google Scholar]
- Darwin, C. Insectivorous Plants; John Murray: London, UK, 1875. [Google Scholar]
- Akeret, B. Ein neuer Fundort von Aldrovanda vesiculosa L. in der Nordschweiz und einige Bemerkungen zu Stratiotes aloides L. Bot. Helv. 1993, 103, 193–199. [Google Scholar]
- Horstmann, M.; Heier, L.; Kruppert, S.; Weiss, L.C.; Tollrian, R.; Adamec, L.; Westermeier, A.; Speck, T.; Poppinga, S. Comparative prey spectra analyses on the endangered aquatic carnivorous waterwheel plant (Aldrovanda vesiculosa, Droseraceae) at several naturalized microsites in the Czech Republic and Germany. Integr. Org. Biol. 2019, 1, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Poppinga, S.; Joyeux, M. Different mechanics of snap-trapping in the two closely related carnivorous plants Dionaea muscipula and Aldrovanda vesiculosa. Phys. Rev. E 2011, 84. [Google Scholar] [CrossRef] [Green Version]
- Poppinga, S.; Smaij, J.; Westermeier, A.S.; Horstmann, M.; Kruppert, S.; Tollrian, R.; Speck, T. Prey capture analyses in the carnivorous aquatic waterwheel plant (Aldrovanda vesiculosa L., Droseraceae). Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poppinga, S.; Bauer, U.; Speck, T.; Volkov, A.G. Motile traps. In Carnivorous Plants: Physiology, Ecology, and Evolution; Elison, A., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 180–193. ISBN 9780198779841. [Google Scholar]
- Westermeier, A.S.; Sachse, R.; Poppinga, S.; Vögele, P.; Adamec, L.; Speck, T.; Bischoff, M. How the carnivorous waterwheel plant (Aldrovanda vesiculosa) snaps. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsuzawa, K.; Kanaizumi, D.; Ajisaka, M.; Kamada, T.; Sakamoto, K.; Matsushima, H.; Kaneko, Y. Fine structure of Aldrovanda vesiculosa L: The peculiar lifestyle of an aquatic carnivorous plant elucidated by electron microscopy using cryo-techniques. Microscopy 2020, 69, 214–226. [Google Scholar] [CrossRef]
- Westermeier, A.S.; Hiss, N.; Speck, T.; Poppinga, S. Functional–morphological analyses of the delicate snap-traps of the aquatic carnivorous waterwheel plant (Aldrovanda vesiculosa) with 2D and 3D imaging techniques. Ann. Bot. 2020, 1099–1107. [Google Scholar] [CrossRef]
- Adamec, L.; Tichý, M. Flowering of Aldrovanda vesiculosa in outdoor culture in the Czech Republic and isozyme variability of its European populations. Carniv. Plant. Newsl. 1997, 26, 99–103. [Google Scholar]
- Maldonado San Martín, A.P.; Adamec, L.; Suda, J.; Mes, T.H.M.; Štorchová, H. Genetic variation within the endangered species Aldrovanda vesiculosa (Droseraceae) as revealed by RAPD analysis. Aquat. Bot. 2003, 75, 159–172. [Google Scholar] [CrossRef]
- Hoshi, Y.; Shirakawa, J.; Hasebe, M. Nucleotide sequence variation was unexpectedly low in an endangered species, Aldrovanda vesiculosa L. (Droseraceae). Chromosom. Bot. 2006, 1, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.M.; Adamec, L.; Štorchová, H. Uniformity of organellar DNA in Aldrovanda vesiculosa, an endangered aquatic carnivorous species, distributed across four continents. Aquat. Bot. 2010, 92, 214–220. [Google Scholar] [CrossRef]
- Govindaraghavan, S.; Hennell, J.R.; Sucher, N.J. From classical taxonomy to genome and metabolome: Towards comprehensive quality standards for medicinal herb raw materials and extracts. Fitoterapia 2012, 83, 979–988. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Y.; Liu, C.; Song, Z.; Li, Q.; Zha, Q.; Lu, C.; Wang, C.; Ning, Z.; Zhang, Y.; et al. The chemotaxonomic classification of Rhodiola plants and its correlation with morphological characteristics and genetic taxonomy. Chem. Cent. J. 2013, 7. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Ni, Y.; Kokot, S. Differentiation of cultivars of flos Chrysanthemum with the use of high-performance liquid chromatography fingerprints and chemometrics. Anal. Lett. 2014, 47, 2023–2034. [Google Scholar] [CrossRef]
- Dresler, S.; Kubrak, T.; Rutkowska, E.; Gagoś, M.; Bogucka-Kocka, A.; Świeboda, R.; Wójcik, M. Comparison of analytical methods in chemometric fingerprinting of metallicolous and non-metallicolous populations of Echium vulgare L. Phytochem. Anal. 2016, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Dresler, S.; Szymczak, G.; Wójcik, M. Comparison of some secondary metabolite content in the seventeen species of the Boraginaceae family. Pharm. Biol. 2017, 55, 691–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strzemski, M.; Dresler, S.; Sowa, I.; Kurach, Ł.; Kováčik, J.; Wojas-Krawczyk, K.; Wójciak, M. Direct spectroscopic and GC profiling combined with chemometric analysis as an alternative approach to investigate Hypericum species: Is it possible to replace HPLC? Ind. Crops Prod. 2020, 157. [Google Scholar] [CrossRef]
- Culham, A.; Gornall, R.J. The taxonomic significance of naphthoquinones in the Droseraceae. Biochem. Syst. Ecol. 1994, 22, 507–515. [Google Scholar] [CrossRef]
- Egan, P.A.; Van Der Kooy, F. Phytochemistry of the carnivorous sundew genus Drosera (Droseraceae)-Future perspectives and ethnopharmacological relevance. Chem. Biodivers. 2013, 10, 1774–1790. [Google Scholar] [CrossRef] [PubMed]
- Braunberger, C.; Zehl, M.; Conrad, J.; Wawrosch, C.; Strohbach, J.; Beifuss, U.; Krenn, L. Flavonoids as chemotaxonomic markers in the genus Drosera. Phytochemistry 2015, 118, 74–82. [Google Scholar] [CrossRef]
- Huber, H. Aldrovanda, L. In Illustrierte Flora von Mitteleuropa; Hegi, G., Ed.; Carl Hanser Verlag: Műnchen, Germany, 1961; pp. 18–20. [Google Scholar]
- Sculthorpe, C.D. The Biology of Aquatic Vascular Plants; Edward Arnold Ltd.: London, UK, 1967. [Google Scholar]
- Huang, Y.J.; Ji, X.P.; Su, T.; Deng, C.L.; Ferguson, D.K.; Yu, T.S.; Yang, X.; Sun, H.; Zhou, Z.K. Habitat, climate and potential plant food resources for the late Miocene Shuitangba hominoid in Southwest China: Insights from carpological remains. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 470, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Messing, J. Analysis of ADP-glucose pyrophosphorylase expression during turion formation induced by abscisic acid in Spirodela polyrhiza (greater duckweed). BMC Plant. Biol. 2012, 12. [Google Scholar] [CrossRef] [Green Version]
- Adamec, L.; Lev, J. The introduction of the aquatic carnivorous plant Aldrovanda vesiculosa to new potential sites in the Czech Republic: A five-year investigation. Folia Geobot. 1999, 34, 299–305. [Google Scholar] [CrossRef]
- Cross, A.T.; Adamec, L.; Turner, S.R.; Dixon, K.W.; Merritt, D.J. Seed reproductive biology of the rare aquatic carnivorous plant Aldrovanda vesiculosa (Droseraceae). Bot. J. Linn. Soc. 2016, 180, 515–529. [Google Scholar] [CrossRef] [Green Version]
- Onelli, E.; Beretta, M.; Moscatelli, A.; Caccianiga, M.; Pozzi, M.; Stroppa, N.; Adamec, L. The aquatic carnivorous plant Aldrovanda vesiculosa (Droseraceae) exhibits altered developmental stages in male gametophyte. Protoplasma 2020. [Google Scholar] [CrossRef]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The ITS region of nuclear ribosomal DNA: A valuable source of evidence on angiosperm phylogeny. Ann. Missouri Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- Buckler, E.S.; Ippolito, A.; Holtsford, T.P. The evolution of ribosomal DNA: Divergent paralogues and phylogenetic implications. Genetics 1997, 145, 821–832. [Google Scholar] [PubMed]
- Xiao, L.Q.; Möller, M.; Zhu, H. High nrDNA ITS polymorphism in the ancient extant seed plant Cycas: Incomplete concerted evolution and the origin of pseudogenes. Mol. Phylogenet. Evol. 2010, 55, 168–177. [Google Scholar] [CrossRef]
- De Miranda, V.F.O.; Martins, V.G.; Furlan, A.; Bacci, M. Plant or fungal sequences? An alternative optimized PCR protocol to avoid ITS (nrDNA) misamplification. Brazilian Arch. Biol. Technol. 2010, 53, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Strzemski, M.; Wójciak-Kosior, M.; Sowa, I.; Kocjan, R.; Tyszczuk-Rotko, K. Methodological approach to determine carlina oxide–a main volatile constituent of Carlina acaulis L. essential oil. Talanta 2019, 191, 504–508. [Google Scholar] [CrossRef]
- Jiang, W.; Zhang, Z.M.; Yun, Y.; Zhan, D.J.; Zheng, Y.B.; Liang, Y.Z.; Yang, Z.Y.; Yu, L. Comparisons of five algorithms for chromatogram alignment. Chromatographia 2013, 76, 1067–1078. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar]
- Ersts, P.J. Geographic Distance Matrix Generator (version 1.2.3). Available online: http://biodiversityinformatics.amnh.org/open_source/gdmg (accessed on 25 December 2020).




{kind=link}
{kind=link}
{kind=link}
| Origin, Site, Country | Geographical Coordinates | Color | Turion Formation | Turion Formation Time |
|---|---|---|---|---|
| Lake Kruglak, NE Poland | 53°54′ N 23°19′ E | green | Yes | Sep |
| Lake Ruzhas, NE Lithuania | 55°30′ N 25°28′ E | green | Yes | Early Sep |
| Ladoga, NE Russia | 60°29′ N 32°57′ E | green | Yes | Early Aug |
| Lake Dolgo’e, Pskov region, W Russia | 56°08′ N 28°22′ E | green | Yes | Late Aug |
| Lake Mokhovo’e, Lipeck, S Russia | 52°24′ N 39°34′ E | green | Yes | Sep |
| Kiev reservoir, N Ukraine | 51°03′ N 30°25′ E | green | Yes | Sep |
| Lake Baláta-tó, Somody county, SW Hungary | 46°19′ N 17°12′ E | red | Yes | Sep |
| Zurich Canton, Switzerland; origin: Lake Constance near Lindau, SW Bavaria, Germany | 47°34′ N 9°41′ E | green | Yes | Sep |
| Lake Prespa, S Northern Macedonia | 41°01′ N 20°59′ E | green | Yes | Sep |
| Hozoji pond near Hanyu town near Tokyo, Japan | 36°12′ N 139°42′ E | green | Yes | Sep |
| Shallow swamp in Okavango delta, Botswana | 19°33′ S 23°13′ E | red | No | - |
| Swamp near Broulee, NSW, SE Australia | 35°35′ S 150°09′ E | red | No | - |
| Girraween Lagoon, S of Darwin, NT, N Australia | 12°31′ S 131°05′ E | red | No | - |
| Leach Lagoon near Katherine, NT, N Australia | 14°38′ S 132°37′ E | red | Yes or No | Oct–Nov |
| Mertens Creek, Kimberley, NT, NW Australia | 14°50′ S 125°41′ E | red | No | - |
| Coastal lake near Esperance Bay, WA, SW Australia | 33°48′ S 121°49′ E | red | No | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Płachno, B.J.; Strzemski, M.; Dresler, S.; Adamec, L.; Wojas-Krawczyk, K.; Sowa, I.; Danielewicz, A.; Miranda, V.F.O. A Chemometry of Aldrovanda vesiculosa L. (Waterwheel, Droseraceae) Populations. Molecules 2021, 26, 72. https://doi.org/10.3390/molecules26010072
Płachno BJ, Strzemski M, Dresler S, Adamec L, Wojas-Krawczyk K, Sowa I, Danielewicz A, Miranda VFO. A Chemometry of Aldrovanda vesiculosa L. (Waterwheel, Droseraceae) Populations. Molecules. 2021; 26(1):72. https://doi.org/10.3390/molecules26010072
Chicago/Turabian StylePłachno, Bartosz J., Maciej Strzemski, Sławomir Dresler, Lubomír Adamec, Kamila Wojas-Krawczyk, Ireneusz Sowa, Anna Danielewicz, and Vitor F. O. Miranda. 2021. "A Chemometry of Aldrovanda vesiculosa L. (Waterwheel, Droseraceae) Populations" Molecules 26, no. 1: 72. https://doi.org/10.3390/molecules26010072
APA StylePłachno, B. J., Strzemski, M., Dresler, S., Adamec, L., Wojas-Krawczyk, K., Sowa, I., Danielewicz, A., & Miranda, V. F. O. (2021). A Chemometry of Aldrovanda vesiculosa L. (Waterwheel, Droseraceae) Populations. Molecules, 26(1), 72. https://doi.org/10.3390/molecules26010072