
Lycopene Inhibits Toll-Like Receptor 4-Mediated Expression of Inflammatory Cytokines in House Dust Mite-Stimulated Respiratory Epithelial Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
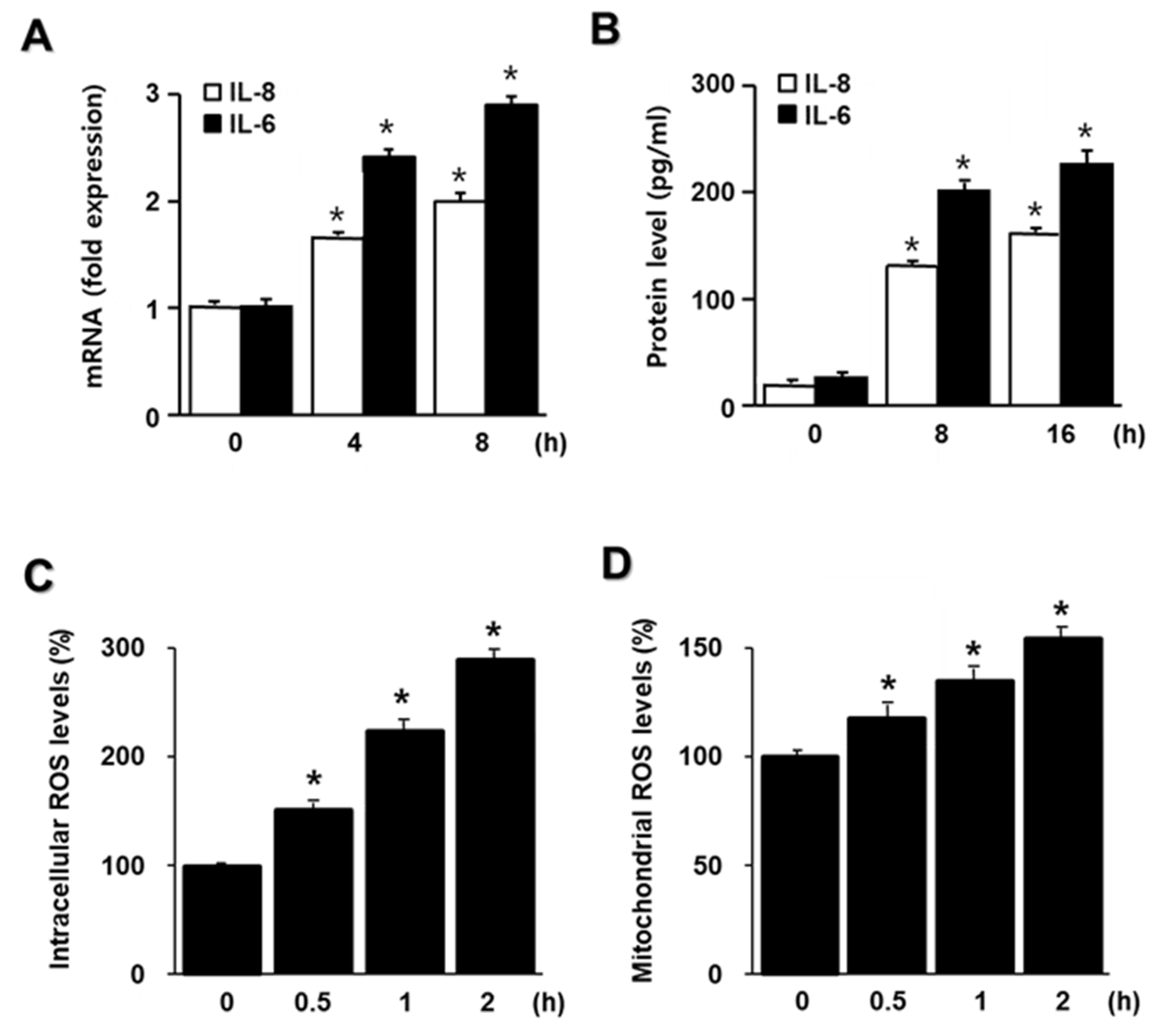
2.1. HDM Induce IL-6 and IL-8 Expression and Increase ROS in A549 Cells
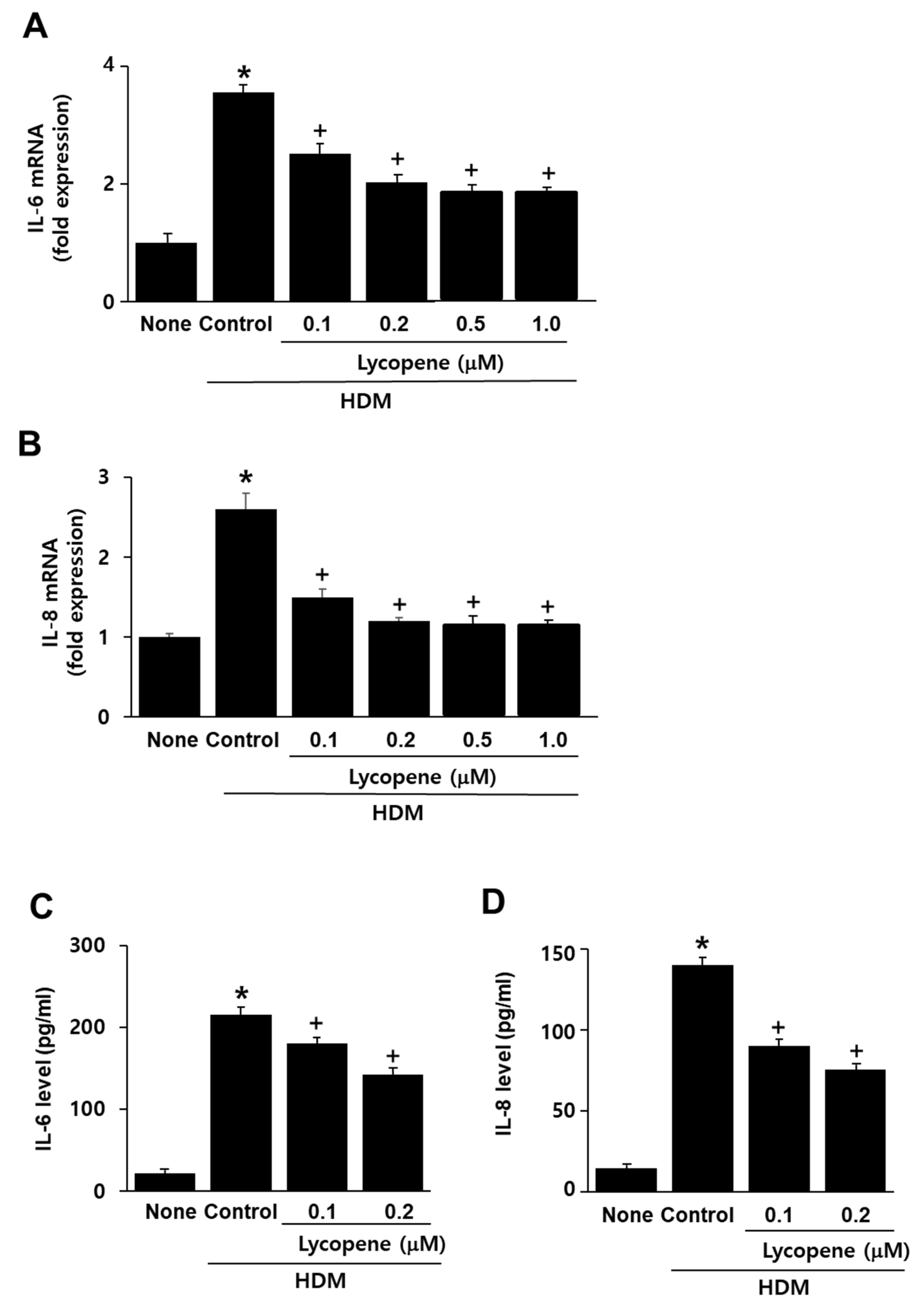
2.2. Lycopene Inhibits HDM-Induced Expression of IL-6 and IL-8 in A549 Cells
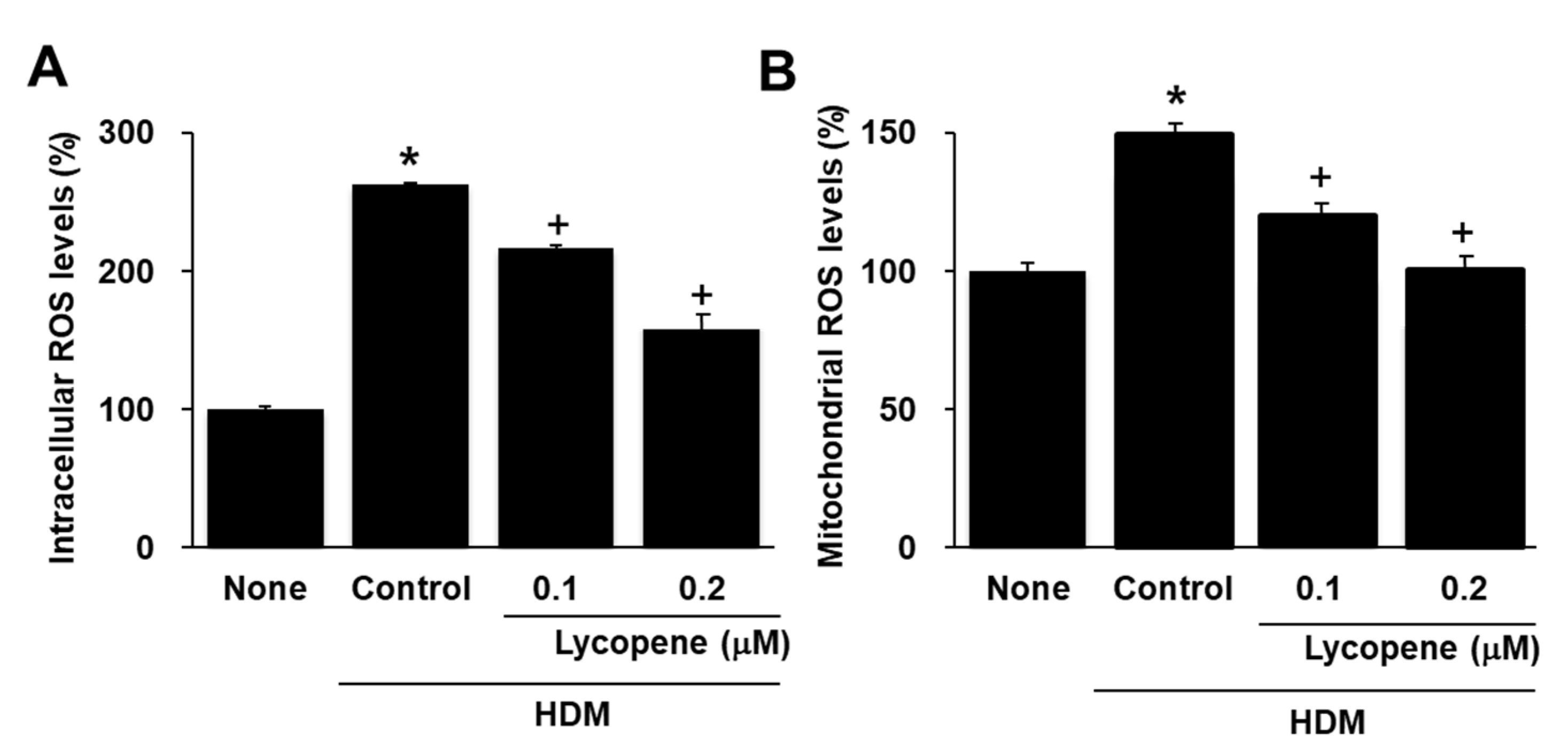
2.3. Lycopene Inhibits HDM-Induced Production of Intracellular and Mitochodrial ROS in A549 Cells
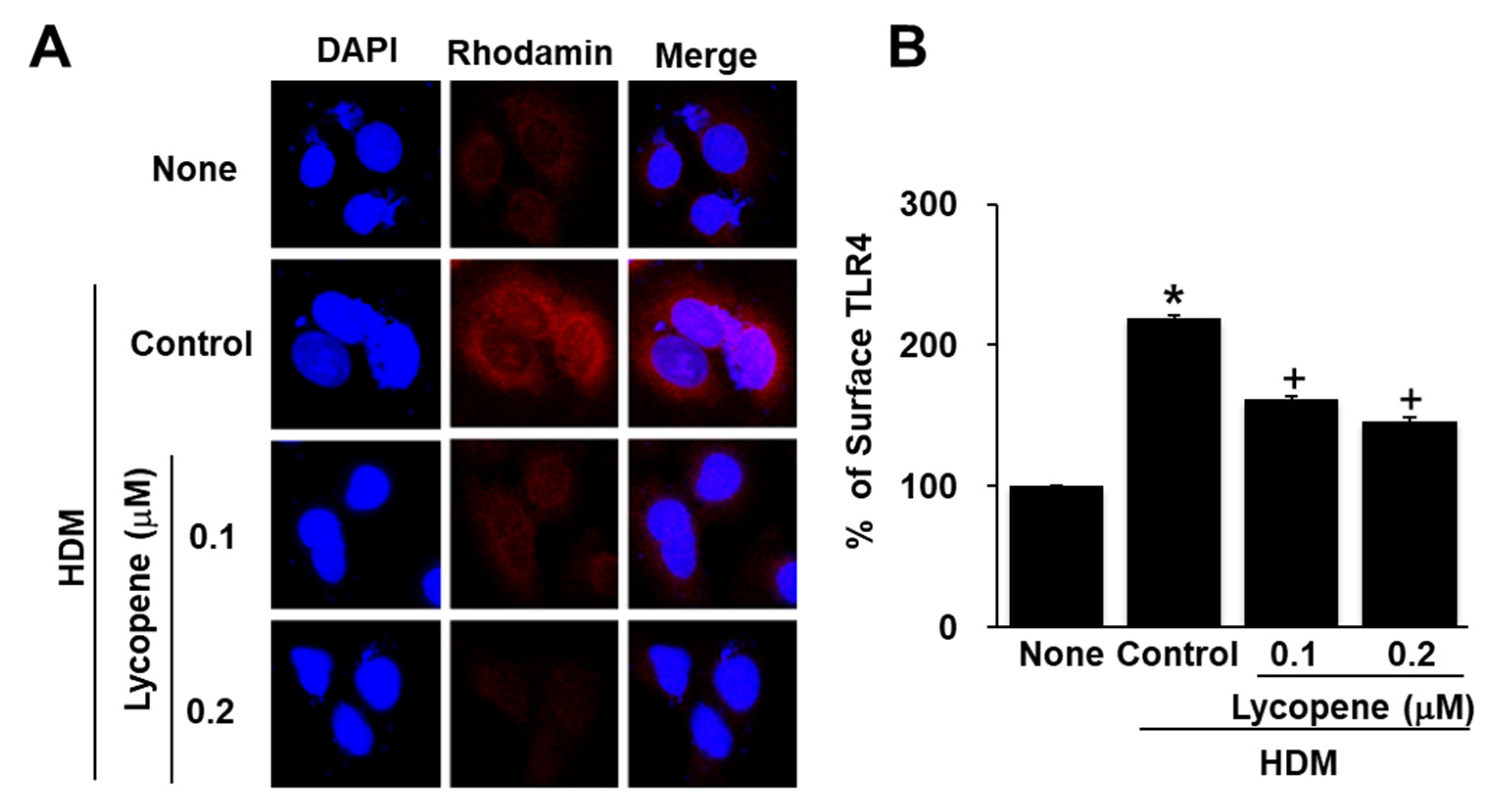
2.4. Lycopene Inhibits HDM-Induced Toll Like Receptor 4 (TLR4) Activation in A549 Cells
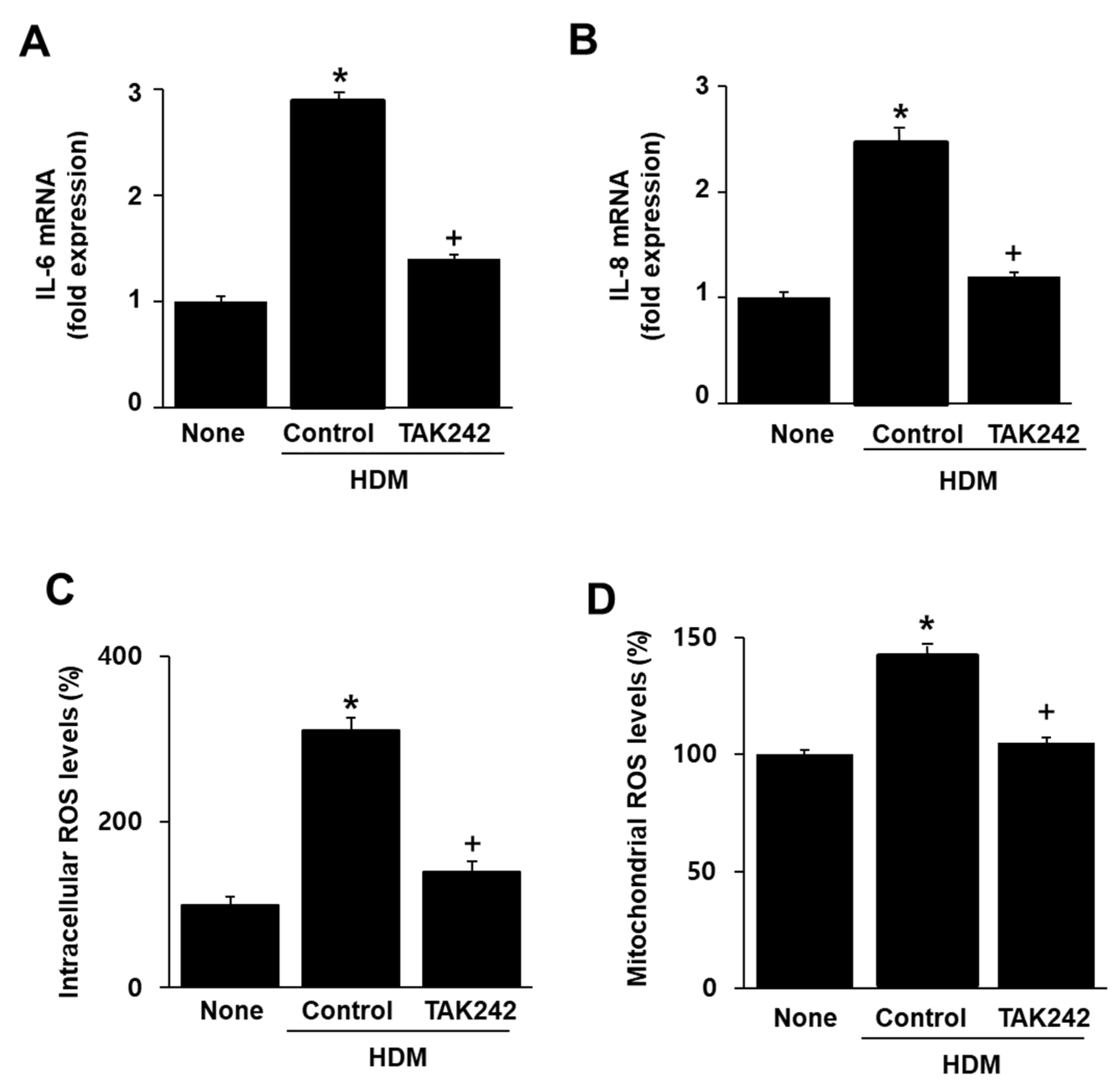
2.5. TAK242 Inhibits the HDM-Induced mRNA Expression of IL-6 and IL-8 and Intracellular and Mitochondrial ROS Increase in A549 Cells
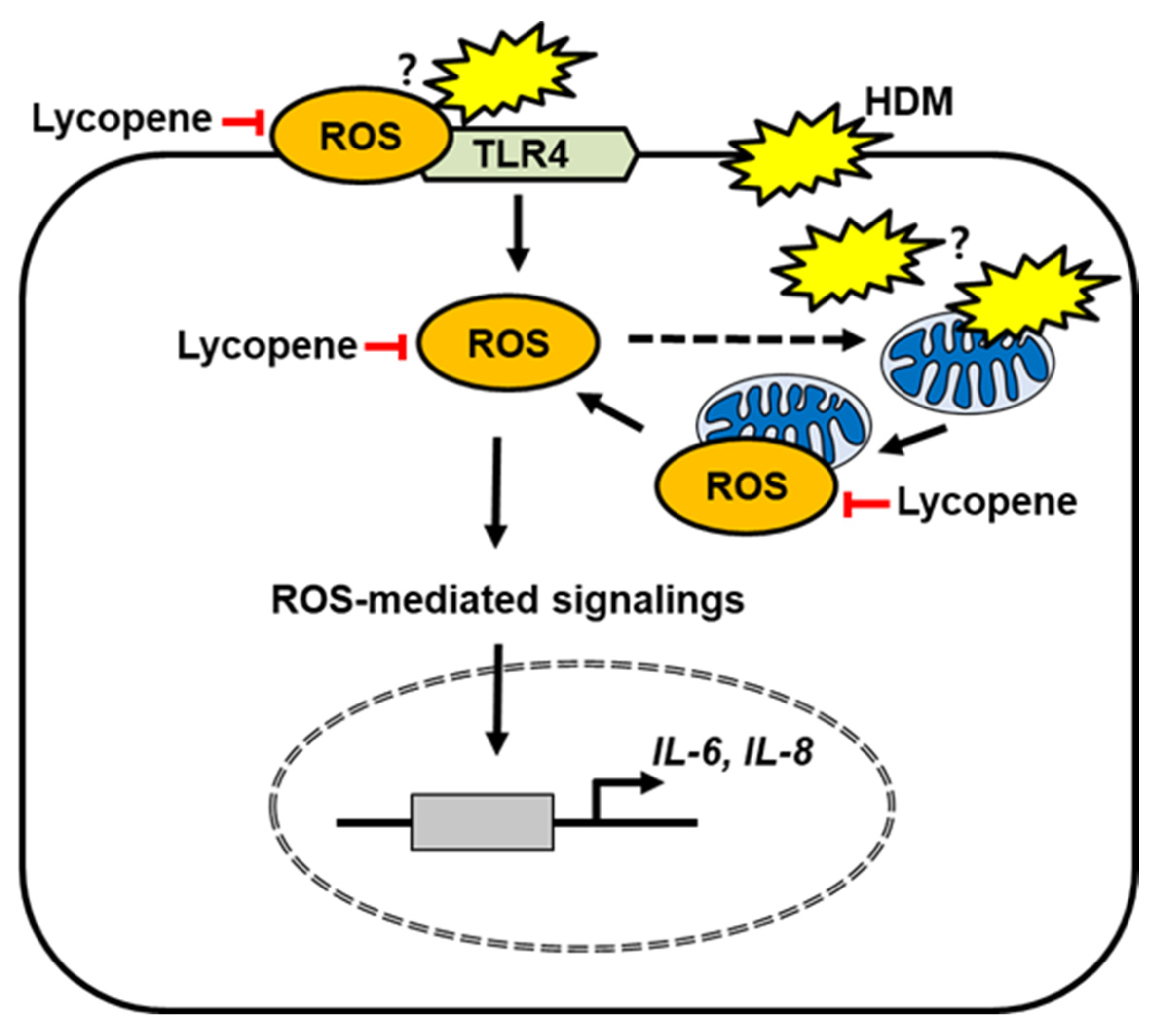
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Line and Culture Conditions
4.3. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) Analysis for IL-6 and IL-8
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Determination of Intracellular ROS and Mitochondrial ROS Levels
4.6. Measurement of Surface TLR4 Level
4.7. Immunofluorescence Staining
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AU | Allergy unit |
| HDM | House dust mites |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| DCF | Dichlorofluorescein |
| DCF-DA | Dichlorofluorescein diacetate |
| DMSO | Dimethyl sulfoxide |
| ELISA | Enzyme-linked immunosorbent assay |
| FACS | Fluorescence-activated single cell sorting |
| LPS | Lipopolysaccharide |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MD-2 | Myeloid differentiation protein 2 |
| NECs | Nasal epithelial cells |
| PAR-2 | Protease activated receptor-2 |
| PE | Phycoerythrin |
| qRT-PCR | Quantitative reverse transcription-polymerase chain reaction |
| ROS | Reactive oxygen species |
| TLR4 | Toll-like receptor 4 |
| THF | Tetrahydrofuran |
| TNF-α | Tumor necrosis factor-α |
References
- Platts-Mills, T.A.; Vervloet, D.; Thomas, W.R.; Aalberse, R.C.; Chapman, M.D. Indoor allergens and asthma: Report of the Third International Workshop. J. Allergy Clin. Immunol. 1997, 100, S2–S24. [Google Scholar] [CrossRef]
- Platts-Mills, T.A.; Carter, M.C. Asthma and Indoor Exposure to Allergens. N. Engl. J. Med. 1997, 336, 1382–1384. [Google Scholar] [CrossRef]
- Ree, H.; Jeon, S.H.; Lee, I.Y.; Hong, C.S.; Lee, D.K. Fauna and geographical distribution of house dust mites in Korea. Korean J. Parasitol. 1997, 35, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Lim, J.W.; Kim, H. Ascorbic Acid Suppresses House Dust Mite-Induced Expression of Interleukin-8 in Human Respiratory Epithelial Cells. J. Cancer Prev. 2021, 26, 64–70. [Google Scholar] [CrossRef]
- Wan, H.; Winton, H.L.; Soeller, C.; Tovey, E.R.; Gruenert, D.C.; Thompson, P.J.; Stewart, G.A.; Taylor, G.W.; Garrod, D.R.; Cannell, M.B.; et al. Der p 1 facilitates transepithelial allergen delivery by disruption of tight junctions. J. Clin. Investig. 1999, 104, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Joldrichsen, M.R.; Amer, A.O.; Boyaka, P.N. Insights into mucosal innate immune responses in house dust mite-mediated allergic asthma. Front. Immunol. 2020, 11, 534501. [Google Scholar]
- Hammad, H.; Chieppa, M.; Perros, F.; Willart, M.A.; Germain, R.N.; Lambrecht, B.N. House dust mite allergen induces asthma via Toll-like receptor 4 triggering of airway structural cells. Nat. Med. 2009, 15, 410–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willart, M.A.; Deswarte, K.; Pouliot, P.; Braun, H.; Beyaert, R.; Lambrecht, B.N.; Hammad, H. Interleukin-1alpha controls allergic sensitization to inhaled house dust mite via the epithelial release of GM-CSF and IL-33. J. Exp. Med. 2012, 209, 1505–1517. [Google Scholar] [CrossRef] [Green Version]
- McAlees, J.W.; Whitehead, G.S.; Harley, I.T.; Cappelletti, M.; Rewerts, C.L.; Holdcroft, A.M. Distinct Tlr4-expressing cell compartments control neutrophilic and eosinophilic airway inflammation. Mucosal Immunol. 2015, 8, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Hristova, M.; Habibovic, A.; Veith, C.; Janssen-Heininger, Y.M.; Dixon, A.E.; Geiszt, M.; van der Vliet, A. Airway epithelial dual oxidase 1 mediates allergen-induced IL-33 secretion and activation of type 2 immune responses. J. Allergy Clin. Immunol. 2016, 137, 1545–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Pei, C.; Yan, S.; Liu, G.; Liu, G.; Chen, W.; Cui, Y.; Liu, Y. NADPH oxidase 1-dependent ROS is crucial for TLR4 signaling to promote tumor metastasis of non-small cell lung cancer. Tumor Biol. 2015, 36, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- . Ryan, K.A.; Smith, M.F., Jr.; Sanders, M.K.; Ernst, P.E. Reactive oxygen and nitrogen species differentially regulate Toll-like receptor 4-mediated activation of NF-kappa B and interleukin-8 expression. IInfect. mmun. 2004, 72, 2123–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brubaker, S.W.; Bonham, K.S.; Zanoni, I.; Kagan, J.C. Innate immune pattern recognition: A cell biological perspective. Annu. Rev. Immunol. 2015, 33, 257–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, R.A.; Brown, G.D. The role of Dectin-1 in the host defence against fungal infections. Curr. Opin. Microbiol. 2011, 14, 392–399. [Google Scholar] [CrossRef]
- Deretic, V.; Saitoh, T.; Akira, S. Autophagy in infection, inflammation and immunity. Nat. Rev. Immunol. 2013, 13, 722–737. [Google Scholar] [CrossRef]
- Lamkanfi, M.; Dixit, V.M. Mechanisms and functions of inflammasomes. Cell 2014, 157, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, P.C.; Castell, J.V.; Andus, T. Interleukin-6 and the acute phase response. Biochem. J. 1990, 265, 621–636. [Google Scholar] [CrossRef]
- Kaplanski, G.; Marin, V.; Montero-Julian, F.; Mantovani, A.; Farnarier, C. IL-6: A regulator of the transition from neutrophil to monocyte recruitment during inflammation. Trends Immunol. 2003, 24, 25–29. [Google Scholar] [CrossRef]
- Tanaka, T.; Kishimoto, T. The biology and medical implications of interleukin-6. Cancer Immunol. Res. 2014, 2, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Atreya, R.; Mudter, J.; Finotto, S.; Müllberg, J.; Jostock, T.; Wirtz, S.; Schütz, M.; Bartsch, B.; Holtmann, M.; Becker, C.; et al. Blockade of interleukin 6 trans signaling suppresses T-cell resistance against apoptosis in chronic intestinal inflammation: Evidence in Crohn disease and experimental colitis in vivo. Nat. Med. 2000, 6, 583–588. [Google Scholar] [CrossRef]
- Roux, J.; McNicholas, C.M.; Carles, M.; Goolaerts, A.; Houseman, B.T.; Dickinson, D.A.; Iles, K.E.; Ware, L.B.; Matthay, M.A.; Pittet, J.-F. IL-8 inhibits cAMP-stimulated alveolar epithelial fluid transport via a GRK2/PI3K-dependent mechanism. FASEB J. 2013, 27, 1095–1106. [Google Scholar] [CrossRef]
- Vlahopoulos, S.; Boldoghm, I.; Casola, A.; Brasierm, A.R. Nuclear factor-kappaB-dependent induction of interleukin-8 gene expression by tumor necrosis factor alpha: Evidence for an antioxidant sensitive activating pathway distinct from nuclear translocation. Blood 1999, 94, 1878–1889. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.Y.; Kim, J.H.; Ahn, S.C.; Lee, H.J.; Moon, D.O.; Lee, C.M.; Park, Y.M. Lycopene suppresses the lipopolysaccharide-induced phenotypic and functional maturation of murine dendritic cells through inhibition of mitogen-activated protein kinases and nuclear factor-κB. Immunology 2004, 113, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Ghorat, F.; Ul-Haq, I.; Ur-Rehman, H.; Aslam, F.; Heydari, M.; Shariati, M.A.; Okuskhanova, E.; Yessimbekov, Z.; Thiruvengadam, M.; et al. Lycopene as a natural antioxidant used to prevent human health disorders. Antioxidants 2020, 9, 706. [Google Scholar] [CrossRef] [PubMed]
- .Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Khachik, F.; Carvalho, L.; Bernstein, P.S.; Muir, G.J.; Zhao, D.Y.; Katz, N.B. Chemistry, distribution and metabolism of tomato carotenoids and their impact on human health. Exp. Biol. Med. 2002, 227, 845–851. [Google Scholar] [CrossRef]
- Rao, A.V.; Ray, M.R.; Rao, L.G. Lycopene. Adv. Food Nutr. Res. 2006, 51, 99–164. [Google Scholar]
- Stahl, W.; Schwarz, W.; Sundquist, A.R.; Sies, H. Cis-trans isomers of lycopene and beta-carotene in human serum and tissues. Arch. Biochem. Biophys. 1992, 294, 173–177. [Google Scholar] [CrossRef]
- Saedisomeolia, A.; Moghadam, A.M. Does lycopene decrease the inflammation in airway epithelial cells? JABS 2011, 5, 81–84. [Google Scholar]
- Dong, J.; Li, W.; Cheng, L.-M.; Wang, G.-G. Lycopene attenuates LPS-induced liver injury by inactivation of NF-κB/COX-2 signaling. Int. J. Clin. Exp. Pathol. 2019, 12, 817–825. [Google Scholar]
- Zou, J.; Feng, D.; Ling, W.-H.; Duan, R.-D. Lycopene suppresses proinflammatory response in lipopolysaccharide-stimulated macrophages by inhibiting ROS-induced trafficking of TLR4 to lipid raft-like domains. J. Nutr. Biochem. 2013, 24, 1117–1122. [Google Scholar] [CrossRef]
- Lee, J.; Lim, J.W.; Kim, H. Lycopene inhibits oxidative stress-mediated inflammatory responses in ethanol/palmitoleic acid-stimulated pancreatic acinar AR42J cells. Int. J. Mol. Sci. 2021, 22, 2101. [Google Scholar] [CrossRef] [PubMed]
- Jhou, B.-Y.; Song, T.-Y.; Lee, I.; Hu, M.-L.; Yang, N.-C. Lycopene inhibits metastasis of human liver adenocarcinoma SKHep-1 cells by downregulation of NADPH oxidase 4 protein expression. J. Agric. Food Chem. 2017, 65, 6893–6903. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Zhuge, F.; Nagashimada, M.; Nagata, N.; Xu, L.; Yamamoto, S.; Fuke, N.; Ushida, Y.; Suganuma, H.; Kaneko, S.; et al. Lycopene prevents the progression of lipotoxicity-induced nonalcoholic steatohepatitis by decreasing oxidative stress in mice. Free Radic. Biol. Med. 2020, 152, 571–582. [Google Scholar] [CrossRef]
- Fukunaga, M.; Gon, Y.; Nunomura, S.; Inoue, T.; Yoshioka, M.; Hashimoto, S.; Ra, C. Protease-mediated house dust mite allergen-induced reactive oxygen species production by neutrophils. Int. Arch. Allergy Immunol. 2011, 155, 104–109. [Google Scholar] [CrossRef]
- Barnes, P.J. Reactive oxygen species and airway inflammation. Free Radic. Biol. Med. 1990, 9, 235–243. [Google Scholar] [CrossRef]
- Wang, E.; Liu, X.; Tu, W.; Do, D.C.; Yu, H.; Yang, L.; Zhou, Y.; Xu, D.; Huang, S.-K.; Yang, P.; et al. Benzo(a)pyrene facilitates dermatophagoides group 1 (Der f 1)-induced epithelial cytokine release through aryl hydrocarbon receptor in asthma. Allergy 2019, 74, 1675–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, M.H.; Kim, H.-P.; Jeong, K.Y.; Kim, C.-R.; Kim, T.Y.; Yong, T.-S. House dust mite allergen Der f 1 induces IL-8 in human basophilic cells via ROS-ERK and p38 signal pathways. Cytokine 2015, 75, 356–364. [Google Scholar] [CrossRef]
- Osterlund, C.; Grönlund, H.; Polovic, N.; Sundström, S.; Gafvelin, G.; Bucht, A. The non-proteolytic house dust mite allergen Der p 2 induce NF-kB and MAPK dependent activation of bronchial epithelial cells. Clin. Exp. Allergy. 2009, 39, 1199–1208. [Google Scholar] [CrossRef]
- Ye, Y.L.; Wu, H.T.; Lin, C.F.; Hsieh, C.Y.; Wang, J.Y.; Liu, F.H.; Ma, C.T.; Bei, C.H.; Cheng, Y.L.; Chen, C.C.; et al. Dermatophagoides pteronyssinus 2 regulates nerve growth factor release to induce airway inflammation via a reactive oxygen species-dependent pathway. Am. J. Physiol Lung Cell Mol. Physiol. 2011, 300, 216–224. [Google Scholar] [CrossRef]
- Jeong, K.Y.; Choi, S.-Y.; Lee, J.-H.; Lee, I.-Y.; Yong, T.-S.; Lee, J.-S.; Hong, C.-S.; Park, J.-W. Standardization of house dust mite extracts in Korea. Allergy Asthma Immunol. Res. 2012, 4, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Ha, J.-H.; Kim, S.-M.; Kim, W.; Kim, K.; Chung, S.-I.; Yoon, Y. β-catenin mediates the inflammatory cytokine expression induced by the Der p 1 house dust mite allergen. Mol. Med. Rep. 2014, 9, 633–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Luo, Q.; Chen, F.; Chen, D.; Xu, G.; Li, H. Induction of IL-6 and IL-8 by house dust mite allergen Der p1 in cultured human nasal epithelial cells is associated with PAR/PI3K/NF B signaling. ORL 2010, 72, 256–265. [Google Scholar] [CrossRef]
- Wong, C.K.; Li, M.L.Y.; Wang, C.B.; Ip, W.K.; Tian, Y.P.; Lam, C.W.K. House dust mite allergen Der p 1 elevates the release of inflammatory cytokines and expression of adhesion molecules in co-culture of human eosinophils and bronchial epithelial cells. Int. Immunol. 2006, 18, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.K.; Tan, W.S.D.; Peh, H.Y.; Wong, W.S.F. Aeroallergens induce reactive oxygen species production and DNA damage and dampen antioxidant responses in bronchial epithelial cells. J. Immunol. 2017, 199, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Lowe, J.; Adcock, I.; Wiegman, C. Oxidative stress and mitochondrial dysfunction in a novel in vivo exacerbation model of severe asthma. Eur. Res. J. 2020, 56, 4083. [Google Scholar]
- Ishii, T.; Niikura, Y.; Kurata, K.; Muroi, M.; Tanamoto, K.; Nagase, T.; Sakaguchi, M.; Yamashita, N. Time-dependent distinct roles of Toll-like receptor 4 in a house dust mite-induced asthma mouse model. Scand. J. Immunol. 2018, 87. [Google Scholar] [CrossRef]
- Park, B.S.; Lee, N.R.; Kim, M.J.; Kim, S.Y.; Kim, I.S. Interaction of Der p 2 with Toll-like receptor 4 and its effect on cytokine secretion. Biomed. Sci. Lett. 2015, 21, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, M.; Miyake, K.; Golenbock, D.T.; Triantafilou, K. Mediators of innate immune recognition of bacteria concentrate in lipid rafts and facilitate lipopolysaccharide-induced cell activation. J. Cell Sci. 2002, 115, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Nakahira, K.; Kim, H.P.; Geng, X.H.; Nakao, A.; Wang, X.; Murase, N.; Drain, P.F.; Wang, X.; Sasidhar, M.; Nabel, E.G.; et al. Carbon monoxide differentially inhibits TLR signaling pathways by regulating ROS-induced trafficking of TLRs to lipid rafts. J. Exp. Med. 2006, 203, 2377–2389. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Guo, S.; Zhang, T.; Yang, Y.; Zhao, G.; Shaukat, A.; Wu, H.; Deng, G. Downregulation of TLR4 by miR-181a provides negative feedback regulation to lipopolysaccharide-induced inflammation. Front. Pharmacol. 2018, 9, 142. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, N.; Tsuchimori, N.; Matsumoto, T.; Ii, M. TAK-242 (resatorvid), a small-molecule inhibitor of Toll-like receptor (TLR) 4 signaling, binds selectively to TLR4 and interferes with interactions between TLR4 and its adaptor molecules. Mol. Pharmacol. 2011, 79, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and physiological relevance of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rissanen, S.; Grzybek, M.; Orłowski, A.; Róg, T.; Cramariuc, O.; Levental, I.; Eggeling, C.; Sezgin, E.; Vattulainen, I. Phase partitioning of GM1 and its bodipy-labeled analog determine their different binding to cholera toxin. Front. Physiol. 2017, 8, 252. [Google Scholar] [CrossRef] [Green Version]
- Anathy, V.; Cunniff, B.; Cahoon, J.M.; Hoffman, S.M.; Taatjes, D.J.; Bouffard, N.A.; Dixon, A.E.; Poynter, M.E.; Heintz, N.H. Endoplasmic reticulum (ER) and mitochondrial interactions modulate house dust mite induced pro-inflammatory response. Am. J. Res. Crit. Care Med. 2015, 191, A5556. [Google Scholar]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Maiani, G.; Casto´n, M.J.; Catasta, G.; Toti, E.; Cambrodón, I.G.; Bysted, A.; Granado-Lorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M.; et al. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food Res. 2009, 53, S194–S218. [Google Scholar] [CrossRef]
- Marcotorchino, J.; Romier, B.; Gouranton, E.; Riollet, C.; Gleize, B.; Malezet-Desmoulins, C.; Landrier, J.-F. Lycopene attenuates LPS-induced TNF-α secretion in macrophages and inflammatory markers in adipocytes exposed to macrophage-conditioned media. Mol. Nutr. Food Res. 2012, 56, 725–732. [Google Scholar] [CrossRef]
- McDevitt, T.M.; Tchao, R.; Harrison, E.H.; Morel, D.W. Carotenoids normally present in serum inhibit proliferation and induce differentiation of a human monocyte/macrophage cell line (U937). J. Nutr. 2005, 135, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Sandhir, R.; Mehrotra, A.; Kamboj, S.S. Lycopene prevents 3-nitropropionic acid-induced mitochondrial oxidative stress and dysfunctions in nervous system. Neurochem. Int. 2010, 57, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Yue, R.; Hu, H.; Yiu, K.H.; Luo, T.; Zhou, Z.; Xu, L.; Zhang, S.; Li, K.; Yu, Z. Lycopene protects against hypoxia/reoxygenation-induced apoptosis by preventing mitochondrial dysfunction in primary neonatal mouse cardiomyocytes. PLoS ONE 2012, 7, e50778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.; Lim, J.W.; Kim, H. Lycopene inhibits reactive oxygen species-mediated NF-κB signaling and induces apoptosis in pancreatic cancer cells. Nutrients. 2019, 11, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Lim, J.W.; Kim, H. Lycopene inhibits activation of epidermal growth factor receptor and expression of cyclooxygenase-2 in gastric cancer cells. Nutrients. 2019, 11, 2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teodoro, A.J.; Oliveira, F.L.; Martins, N.B.; de Azevedo Maia, G.; Martucci, R.B.; Borojevic, R. Effect of lycopene on cell viability and cell cycle progression in human cancer cell lines. Cancer Cell Int. 2012, 12, 36. [Google Scholar] [CrossRef] [Green Version]
- Trejo-Solís, C.; Chaverrí, J.P.; Ramos, M.T.; Farfán, D.; Salgado, A.-C.; Serrano-García, N.; Rico, L.O.; Sotelo, J. Multiple molecular and cellular mechanisms of action of lycopene in cancer inhibition. Evid-Based-Comp. Alt. Med. 2013, 2013, 705121. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chen, J.; Mangat, S.C.; Perera Baruhupolage, C.; Garrod, D.R.; Robinson, C. Pathways of airway oxidant formation by house dust mite allergens and viral RNA converge through myosin motors, pannexons and Toll-like receptor 4. Immun. Inflamm. Dis. 2018, 6, 276–296. [Google Scholar] [CrossRef]
- Ryu, J.H.; Yoo, J.Y.; Kim, M.J.; Hwang, S.G.; Ahn, K.C.; Ryu, J.C.; Choi, M.K.; Joom, J.H.; Kim, C.H.; Lee, S.N.; et al. Distinct TLR-mediated pathways regulate house dust mite-induced allergic disease in the upper and lower airways. J. Allergy Clin. Immunol. 2013, 131, 549–561. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.; Lim, J.W.; Kim, H. Lycopene Inhibits Toll-Like Receptor 4-Mediated Expression of Inflammatory Cytokines in House Dust Mite-Stimulated Respiratory Epithelial Cells. Molecules 2021, 26, 3127. https://doi.org/10.3390/molecules26113127
Choi J, Lim JW, Kim H. Lycopene Inhibits Toll-Like Receptor 4-Mediated Expression of Inflammatory Cytokines in House Dust Mite-Stimulated Respiratory Epithelial Cells. Molecules. 2021; 26(11):3127. https://doi.org/10.3390/molecules26113127
Chicago/Turabian StyleChoi, Jiyeon, Joo Weon Lim, and Hyeyoung Kim. 2021. "Lycopene Inhibits Toll-Like Receptor 4-Mediated Expression of Inflammatory Cytokines in House Dust Mite-Stimulated Respiratory Epithelial Cells" Molecules 26, no. 11: 3127. https://doi.org/10.3390/molecules26113127
APA StyleChoi, J., Lim, J. W., & Kim, H. (2021). Lycopene Inhibits Toll-Like Receptor 4-Mediated Expression of Inflammatory Cytokines in House Dust Mite-Stimulated Respiratory Epithelial Cells. Molecules, 26(11), 3127. https://doi.org/10.3390/molecules26113127