
Ruthenacarborane and Quinoline: A Promising Combination for the Treatment of Brain Tumors

 , and
, and 
Abstract
:
1. Introduction
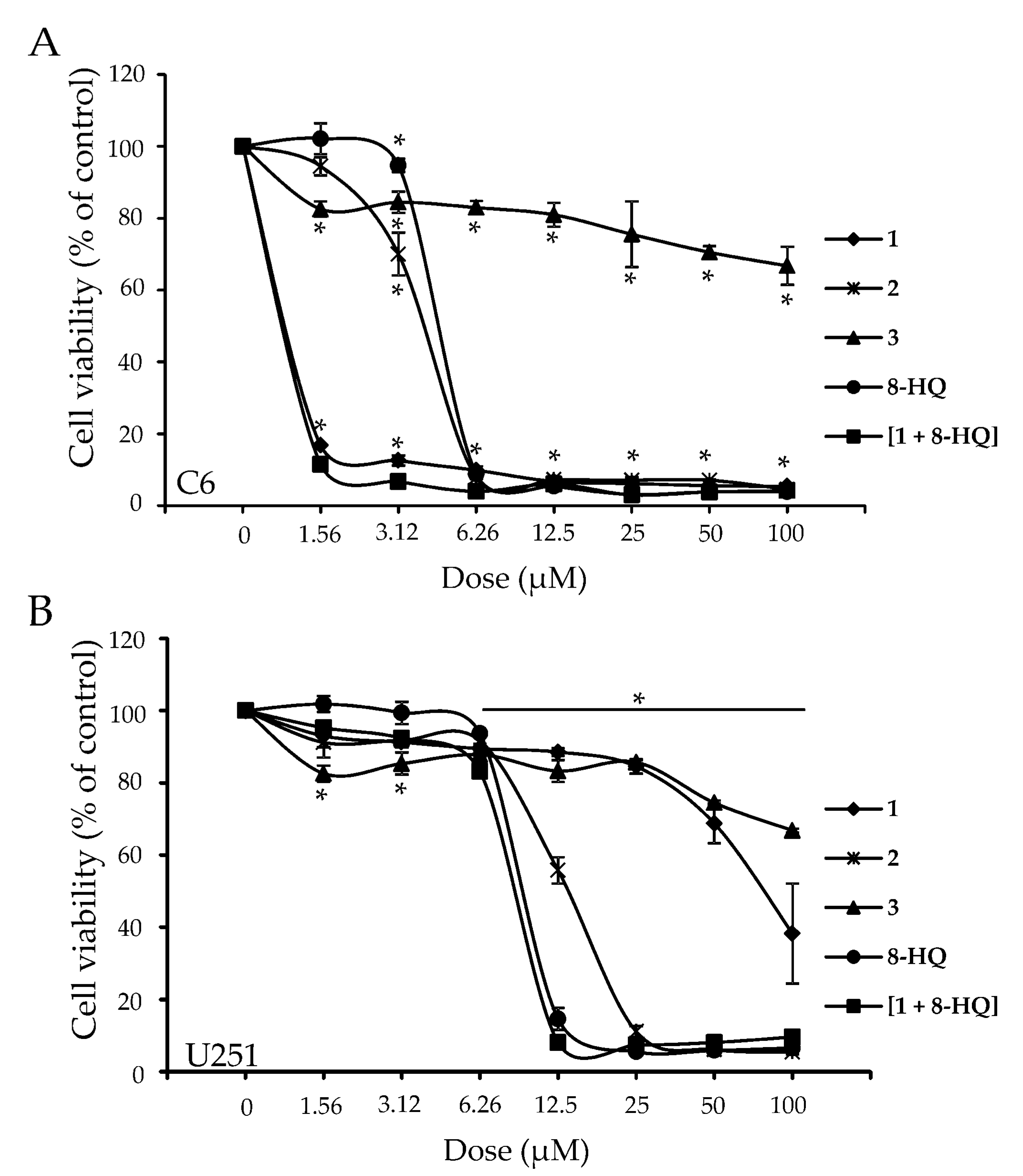
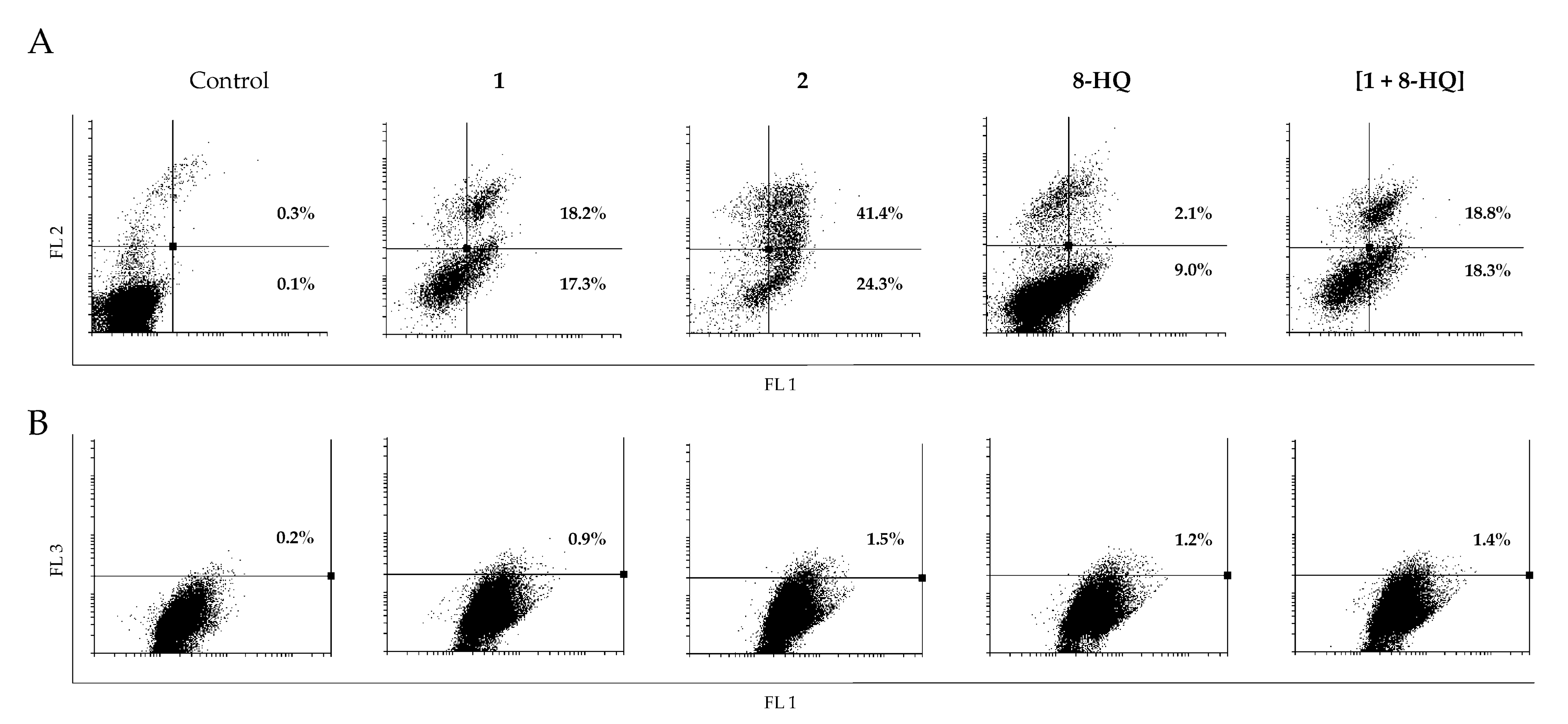
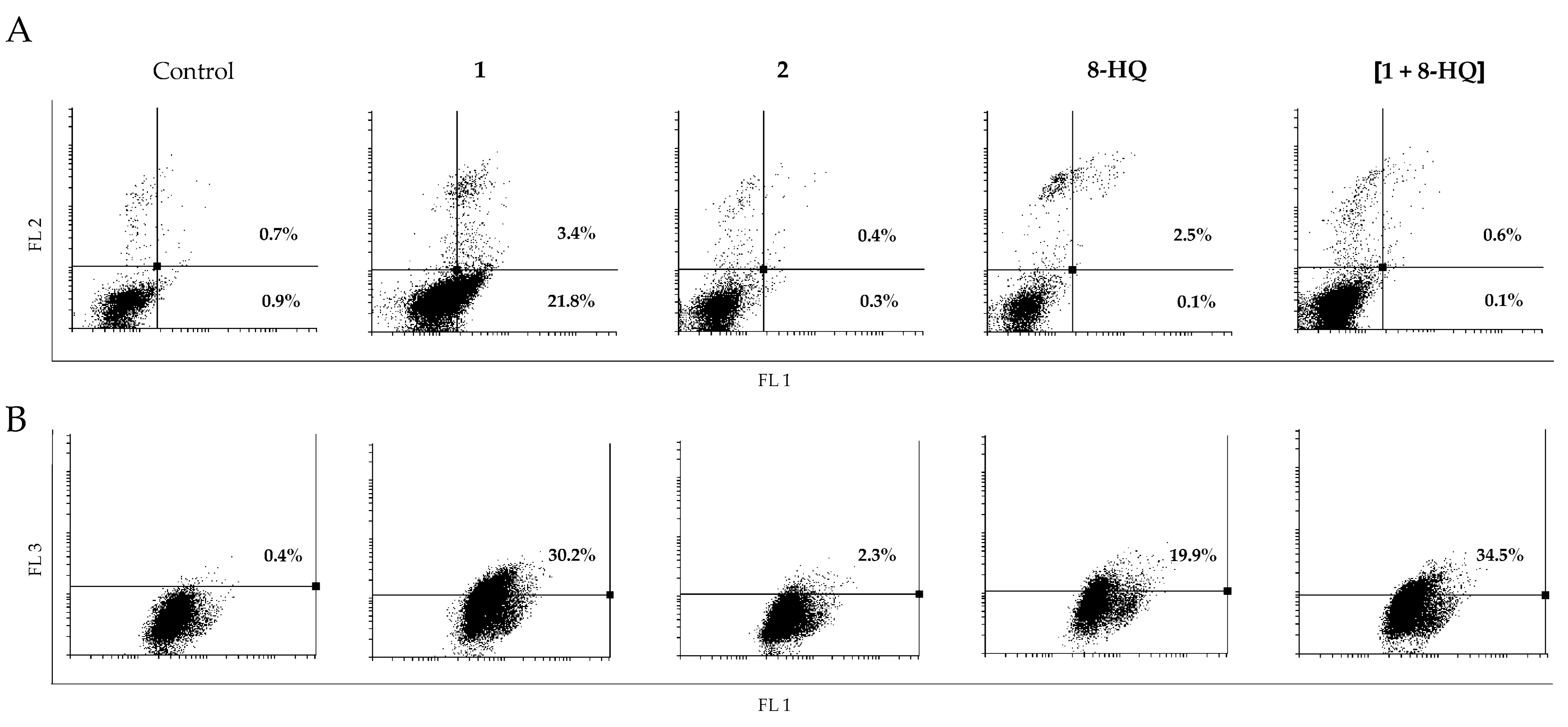
2. Results
3. Materials and Methods
3.1. Chemicals
3.2. Reagents and Cells
3.3. Preparation of the Drug Solutions
3.4. Cell Viability Assays
3.5. CFSE Staining
3.6. Annexin V-FITC/PI and AO Staining
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Murphy, N.; McCarthy, E.; Dwyer, R.; Farràs, P. Boron clusters as breast cancer therapeutics. J. Inorg. Biochem. 2021, 218, 111412–111422. [Google Scholar] [CrossRef] [PubMed]
- Stockmann, P.; Gozzi, M.; Kuhnert, R.; Sárosi, M.-B.; Hey-Hawkins, E. New keys for old locks: Carborane-containing drugs as platforms for mechanism-based therapies. Chem. Soc. Rev. 2019, 48, 3497–3512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zargham, E.O.; Mason, C.A.; Lee, M.W., Jr. The use of carboranes in cancer drug development. Int. J. Cancer Clin. Res. 2019, 6, 110–113. [Google Scholar]
- Hey-Hawkins, E.; Viñas-Teixidor, C. Boron-Based Compounds: Potential and Emerging Applications in Medicine; John Wiley & Sons Incorporated: Hoboken, NJ, USA, 2018; ISBN 978-1-119-27555-8. [Google Scholar]
- Leśnikowski, Z.J. Challenges and opportunities for the application of boron clusters in drug design. J. Med. Chem. 2016, 59, 7738–7758. [Google Scholar] [CrossRef] [PubMed]
- Barth, R.F.; Mi, P.; Yang, W. Boron delivery agents for neutron capture therapy of cancer. Cancer Commun. 2018, 38, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimes, R.N. Carboranes, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 945–984. ISBN 9780128018941. [Google Scholar]
- Kuhnert, R.; Sárosi, M.-B.; George, S.; Lönnecke, P.; Hofmann, B.; Steinhilber, D.; Murganic, B.; Mijatović, S.; Maksimović-Ivanić, D.; Hey-Hawkins, E. CarbORev-5901: The first carborane-based inhibitor of the 5-lipoxygenase pathway. ChemMedChem 2017, 12, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, B.; Jelača, S.; Welcke, L.; Maksimović-Ivanić, D.; Mijatović, S.; Hey-Hawkins, E. 2,2’-Bipyridine-modified tamoxifen: A versatile vector for molybdacarboranes. ChemMedChem 2019, 14, 2075–2083. [Google Scholar] [CrossRef] [Green Version]
- Scholz, M.; Steinhagen, M.; Heiker, J.T.; Beck-Sickinger, A.G.; Hey-Hawkins, E. Asborin inhibits Aldo/Keto reductase 1A1. ChemMedChem 2011, 6, 89–93. [Google Scholar] [CrossRef]
- Neumann, W.; Xu, S.; Sárosi, M.-B.; Scholz, M.S.; Crews, B.C.; Ghebreselasie, K.; Banerjee, S.; Marnett, L.J.; Hey-Hawkins, E. nido-Dicarbaborate induces potent and selective inhibition of cyclooxygenase-2. ChemMedChem 2016, 11, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Gozzi, M.; Schwarze, B.; Hey-Hawkins, E. Preparing (metalla)carboranes for nanomedicine. ChemMedChem 2021, 16. [Google Scholar] [CrossRef] [PubMed]
- Gozzi, M.; Murganic, B.; Drača, D.; Popp, J.; Coburger, P.; Maksimović-Ivanić, D.; Mijatović, S.; Hey-Hawkins, E. Quinoline-conjugated ruthenacarboranes: Toward hybrid drugs with a dual mode of action. ChemMedChem 2019, 14, 2061–2074. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.E.; Gammoh, N. The impact of autophagy during the development and survival of glioblastoma. Open Biol. 2020, 10, 200184–200197. [Google Scholar] [CrossRef]
- Gallagher, L.E.; Radhi, O.A.; Abdullah, M.O.; McCluskey, A.G.; Boyd, M.; Chan, E.Y.W. Lysosomotropism depends on glucose: A chloroquine resistance mechanism. Cell Death Dis. 2017, 8, 3014–3027. [Google Scholar] [CrossRef] [PubMed]
- Varisli, L.; Cen, O.; Vlahopoulos, S. Dissecting pharmacological effects of chloroquine in cancer treatment: Interference with inflammatory signaling pathways. Immunology 2020, 159, 257–278. [Google Scholar] [CrossRef] [Green Version]
- Motaln, H.; Koren, A.; Gruden, K.; Ramšak, Ž.; Schichor, C.; Lah, T.T. Heterogeneous glioblastoma cell cross-talk promotes phenotype alterations and enhanced drug resistance. Oncotarget 2015, 6, 40998–41017. [Google Scholar] [CrossRef] [Green Version]
- Diao, W.; Tong, X.; Yang, C.; Zhang, F.; Bao, C.; Chen, H.; Liu, L.; Li, M.; Ye, F.; Fan, Q.; et al. Behaviors of glioblastoma cells in in vitro microenvironments. Sci. Rep. 2019, 9, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couto, M.; Mastandrea, I.; Cabrera, M.; Cabral, P.; Teixidor, F.; Cerecetto, H.; Viñas, C. Small-molecule kinase-inhibitors-loaded boron cluster as hybrid agents for glioma-cell-targeting therapy. Chem. Eur. J. 2017, 23, 9233–9238. [Google Scholar] [CrossRef]
- Couto, M.; García, M.F.; Alamón, C.; Cabrera, M.; Cabral, P.; Merlino, A.; Teixidor, F.; Cerecetto, H.; Viñas, C. Discovery of potent EGFR inhibitors through the incorporation of a 3D-aromatic-boron-rich-cluster into the 4-anilinoquinazoline scaffold: Potential drugs for glioma treatment. Chem. Eur. J. 2018, 24, 3122–3126. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.; Alamón, C.; Nievas, S.; Perona, M.; Dagrosa, M.A.; Teixidor, F.; Cabral, P.; Viñas, C.; Cerecetto, H. Bimodal therapeutic agents against glioblastoma, one of the most lethal forms of cancer. Chem. Eur. J. 2020, 26, 14335–14340. [Google Scholar] [CrossRef]
- Wilkinson, S.M.; Gunosewoyo, H.; Barron, M.L.; Boucher, A.; McDonnell, M.; Turner, P.; Morrison, D.E.; Bennett, M.R.; McGregor, I.S.; Rendina, L.M.; et al. The first CNS-active carborane: A novel P2 × 7 receptor antagonist with antidepressant activity. ACS Chem. Neurosci. 2014, 5, 335–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyerhäuser, P.; Kantelhardt, S.R.; Kim, E.L. Re-purposing chloroquine for glioblastoma: Potential merits and confounding variables. Front. Oncol. 2018, 8, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Giakoumettis, D.; Kritis, A.; Foroglou, N. C6 cell line: The gold standard in glioma research. Hippokratia 2018, 22, 105–112. [Google Scholar]
- Song, Y.; Xu, H.; Chen, W.; Zhan, P.; Liu, X. 8-Hydroxyquinoline: A privileged structure with a broad-ranging pharmacological potential. MedChemComm 2015, 6, 61–74. [Google Scholar] [CrossRef]
- Madonna, S.; Béclin, C.; Laras, Y.; Moret, V.; Marcowycz, A.; Lamoral-Theys, D.; Dubois, J.; Barthelemy-Requin, M.; Lenglet, G.; Depauw, S.; et al. Structure-activity relationships and mechanism of action of antitumor bis 8-hydroxyquinoline substituted benzylamines. Eur. J. Med. Chem. 2010, 45, 623–638. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, Z.; Wei, Z.-Z.; Qin, Q.-P.; Yang, L.; Liang, H. High anticancer activity and apoptosis- and autophagy-inducing properties of novel lanthanide (III) complexes bearing 8-hydroxyquinoline-N-oxide and 1,10-phenanthroline. Dalton Trans. 2021, 50, 5828–5834. [Google Scholar] [CrossRef]
- Al-Bari, M.A.A. Chloroquine analogues in drug discovery: New directions of uses, mechanisms of actions and toxic manifestations from malaria to multifarious diseases. J. Antimicrob. Chemother. 2015, 70, 1608–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, Y.H.; Cho, K.S.; Hwang, J.J.; Lee, S.-J.; Choi, J.A.; Koh, J.-Y. Induction of lysosomal dilatation, arrested autophagy, and cell death by chloroquine in cultured ARPE-19 cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6030–6037. [Google Scholar] [CrossRef] [Green Version]
- Mauthe, M.; Orhon, I.; Rocchi, C.; Zhou, X.; Luhr, M.; Hijlkema, K.-J.; Coppes, R.P.; Engedal, N.; Mari, M.; Reggiori, F. Chloroquine inhibits autophagic flux by decreasing autophagosome-lysosome fusion. Autophagy 2018, 14, 1435–1455. [Google Scholar] [CrossRef] [PubMed]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef]
- Lau, A.N.; Vander Heiden, M.G. Metabolism in the tumor microenvironment. Annu. Rev. Cancer Biol. 2020, 4, 17–40. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (μM) | |||
|---|---|---|---|
| Compound | Assay 1 | C6 | U251 |
| 1 | MTT | 0.8 ± 0.1 | 61.9 ± 11.4 |
| CV | 1.0 ± 0.1 | 75.3 ± 7.6 | |
| 2 | MTT | 3.3 ± 0.3 | 9.2 ± 1.0 |
| CV | 14.4 ± 0.1 | 11.4 ± 3.9 | |
| 3 | MTT | >100 | >100 |
| CV | >100 | >100 | |
| 8-HQ | MTT | 3.8 ± 0.4 | 9.0 ± 0.8 |
| CV | 4.7 ± 0.1 | 9.6 ± 0.2 | |
| [1 + 8-HQ] 2 | MTT | 0.9 ± 0.1 | 7.4 ± 0.2 |
| CV | 1.0 ± 0.1 | 8.8 ± 0.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drača, D.; Marković, M.; Gozzi, M.; Mijatović, S.; Maksimović-Ivanić, D.; Hey-Hawkins, E. Ruthenacarborane and Quinoline: A Promising Combination for the Treatment of Brain Tumors. Molecules 2021, 26, 3801. https://doi.org/10.3390/molecules26133801
Drača D, Marković M, Gozzi M, Mijatović S, Maksimović-Ivanić D, Hey-Hawkins E. Ruthenacarborane and Quinoline: A Promising Combination for the Treatment of Brain Tumors. Molecules. 2021; 26(13):3801. https://doi.org/10.3390/molecules26133801
Chicago/Turabian StyleDrača, Dijana, Milan Marković, Marta Gozzi, Sanja Mijatović, Danijela Maksimović-Ivanić, and Evamarie Hey-Hawkins. 2021. "Ruthenacarborane and Quinoline: A Promising Combination for the Treatment of Brain Tumors" Molecules 26, no. 13: 3801. https://doi.org/10.3390/molecules26133801
APA StyleDrača, D., Marković, M., Gozzi, M., Mijatović, S., Maksimović-Ivanić, D., & Hey-Hawkins, E. (2021). Ruthenacarborane and Quinoline: A Promising Combination for the Treatment of Brain Tumors. Molecules, 26(13), 3801. https://doi.org/10.3390/molecules26133801