
Δ8(14)-Ergostenol Glycoside Derivatives Inhibit the Expression of Inflammatory Mediators and Matrix Metalloproteinase

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Instruments
2.2. Synthesis of 5α-Ergost-8(14)-en-3β-ol (Ergn, 3)
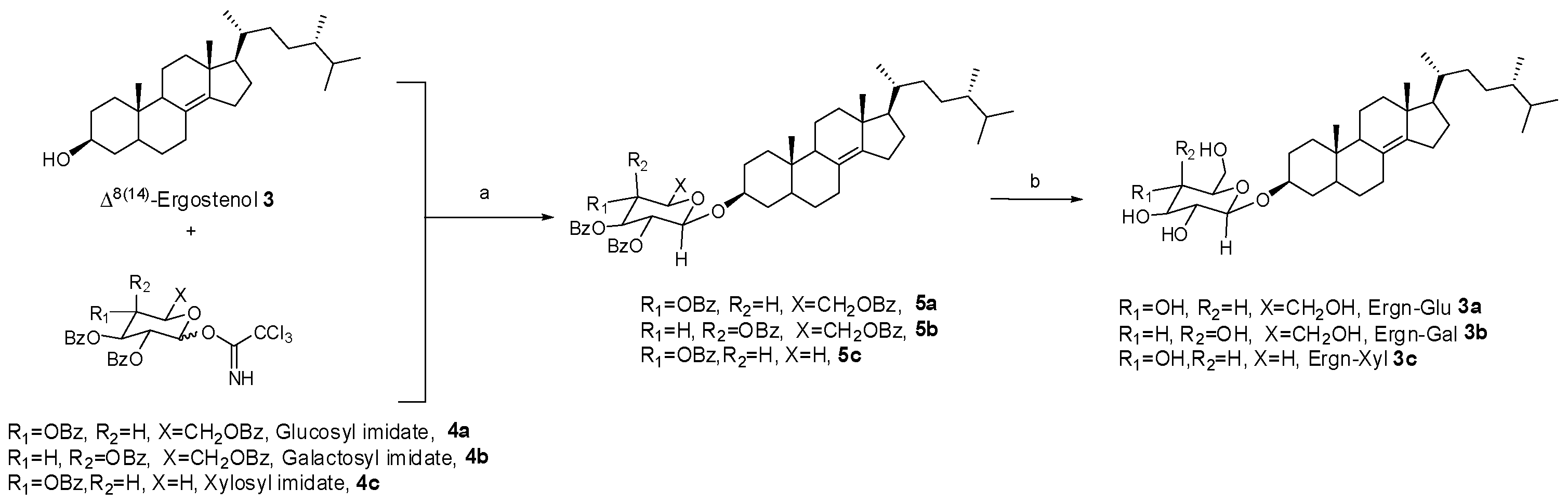
2.3. Synthesis of Ergostenol Glycosides (3a, 3b, and 3c) through Glycosylation and Hydrolysis
2.3.1. Synthesis of 5α-Ergost-8(14)-en-3β-yl-β-d-Glucopyranoside (Ergn-Glu 3a)
2.3.2. Synthesis of 5α-Ergost-8(14)-en-3β-yl-β-d-Galactopyranoside (Ergn-Gal, 3b)
2.3.3. Synthesis of 5α-Ergost-8(14)-en-3β-yl-β-d-Xylopyranoside (Ergn-Xyl, 3c)
2.4. Biological Evaluation of Materials
2.5. Cell Culture
2.6. Cell Viability Assay
2.7. Reverse Transcription-Polymerase Chain Reaction (PCR)
2.8. Western Blotting
2.9. Enzyme-Linked Immunosorbent Assay
2.10. Statistical Analysis
3. Results
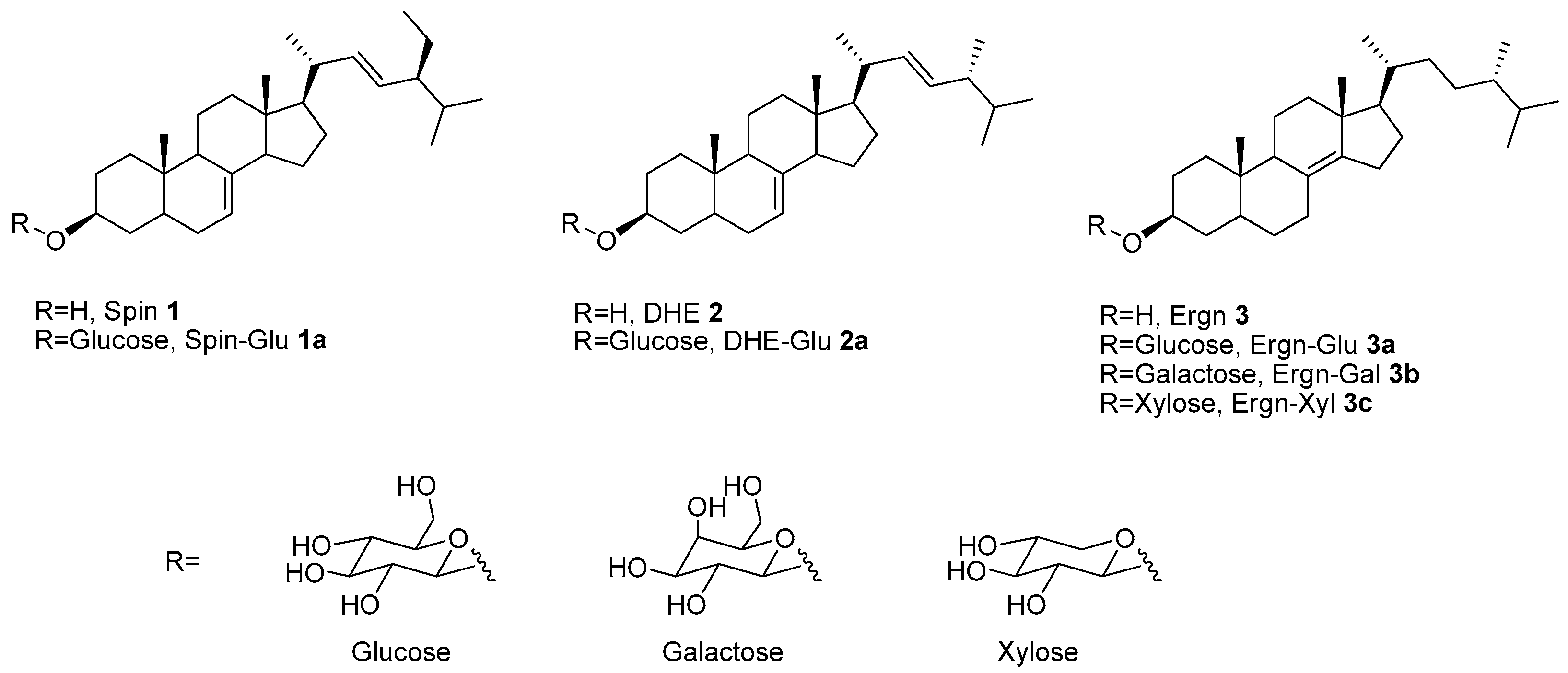
3.1. Synthesis of Δ8(14)-Ergostenol Glycosides (Ergn-Gly, 3a, 3b, and 3c)
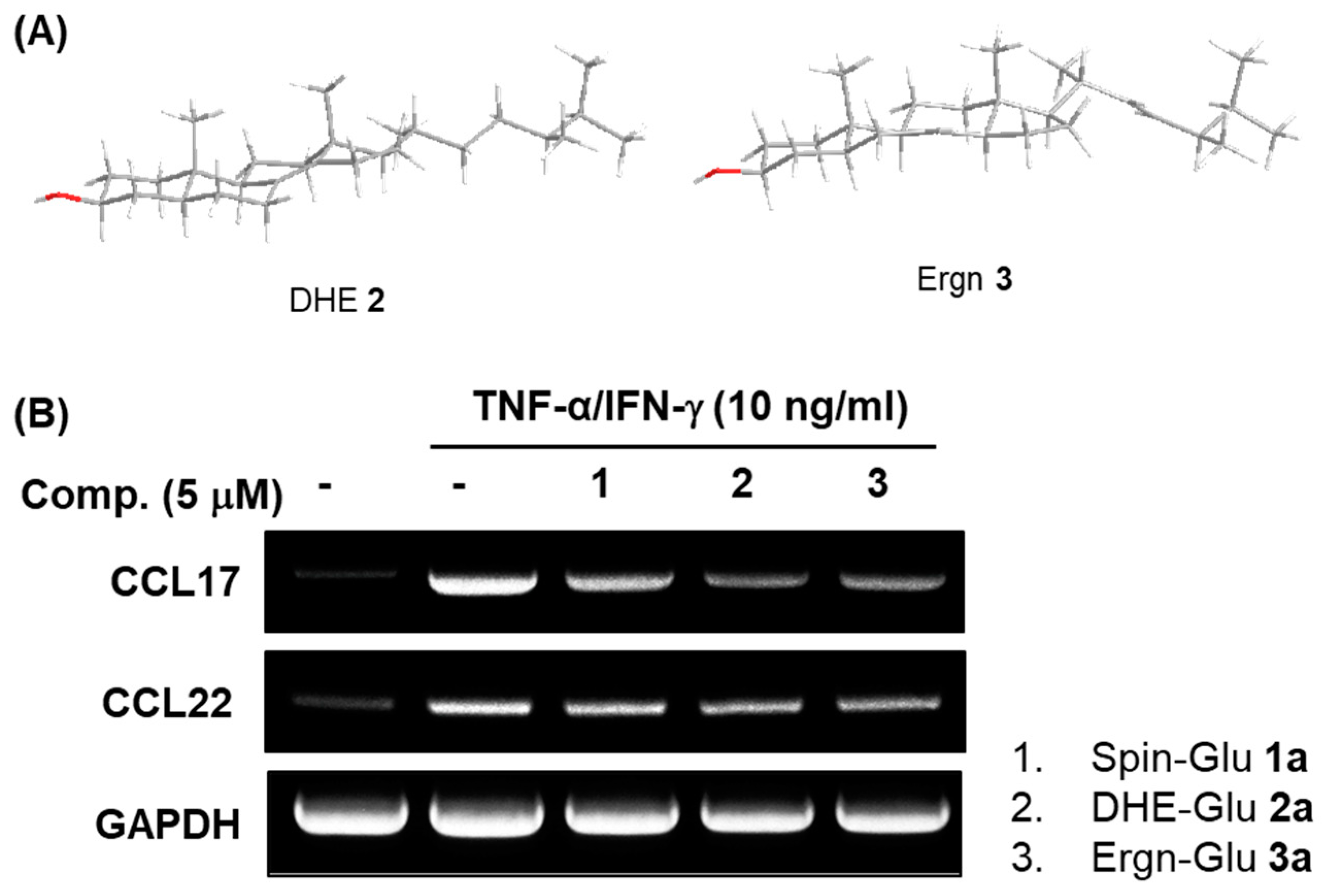
3.2. Comparison of the Structures of DHE (2) and Ergn (3) and Biological Activities of their Glycosides
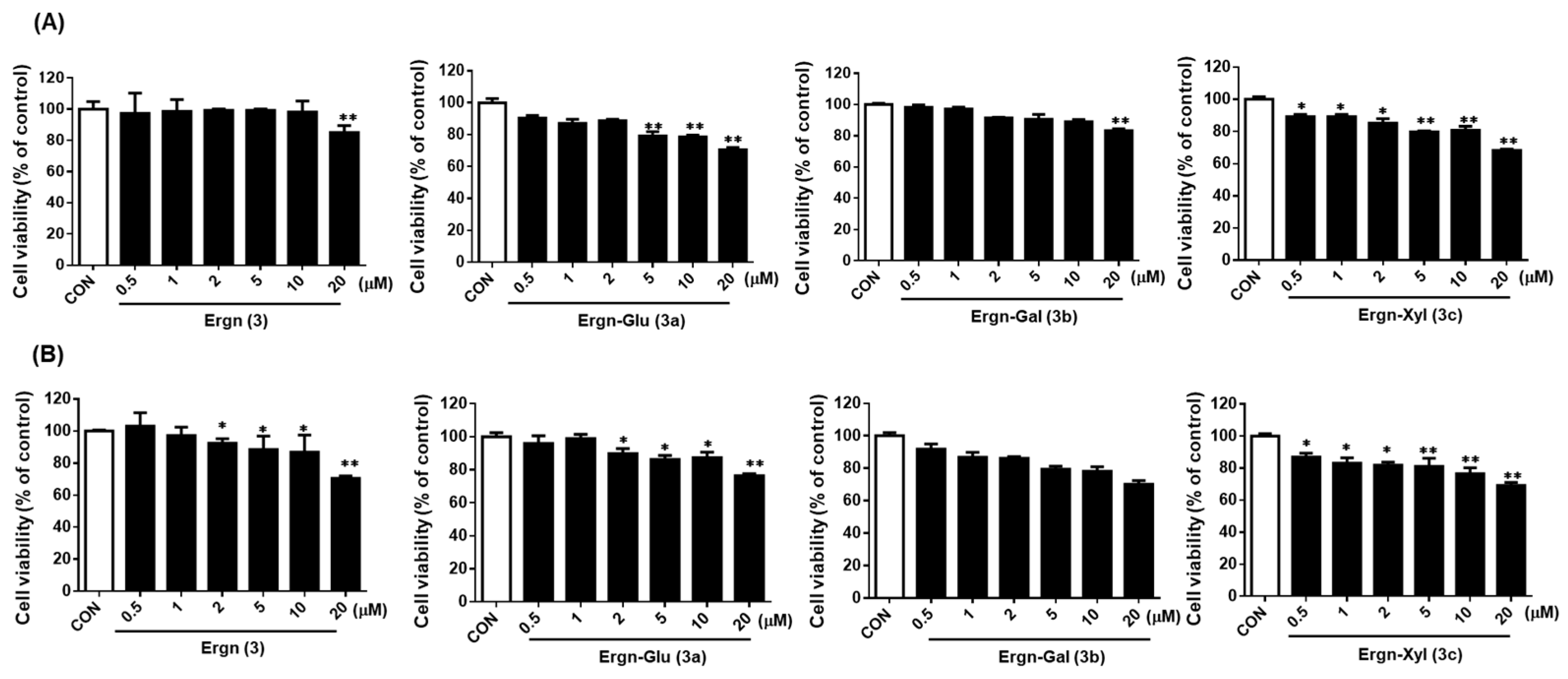
3.3. Effect of Ergostenol Glycosides on the Viability of Human Chondrocytes (HCs) and Human Fibroblast-Like Synoviocytes (FLSs)
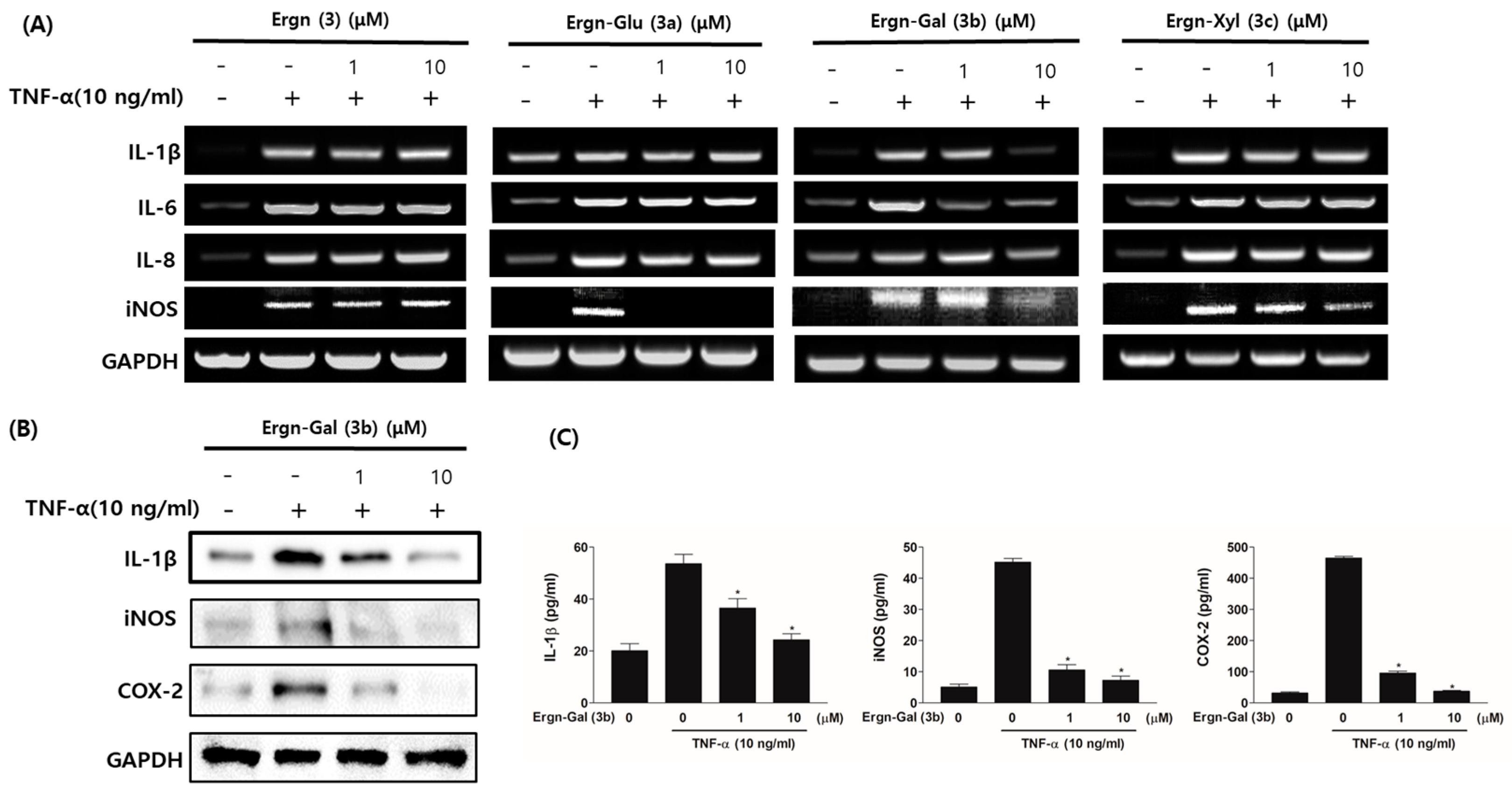
3.4. Ergostenol Glycosides Inhibited TNF-α-Induced Expression of the Pro-Inflammatory Cytokines in Human Fibroblast-Like Synoviocytes (FLSs)
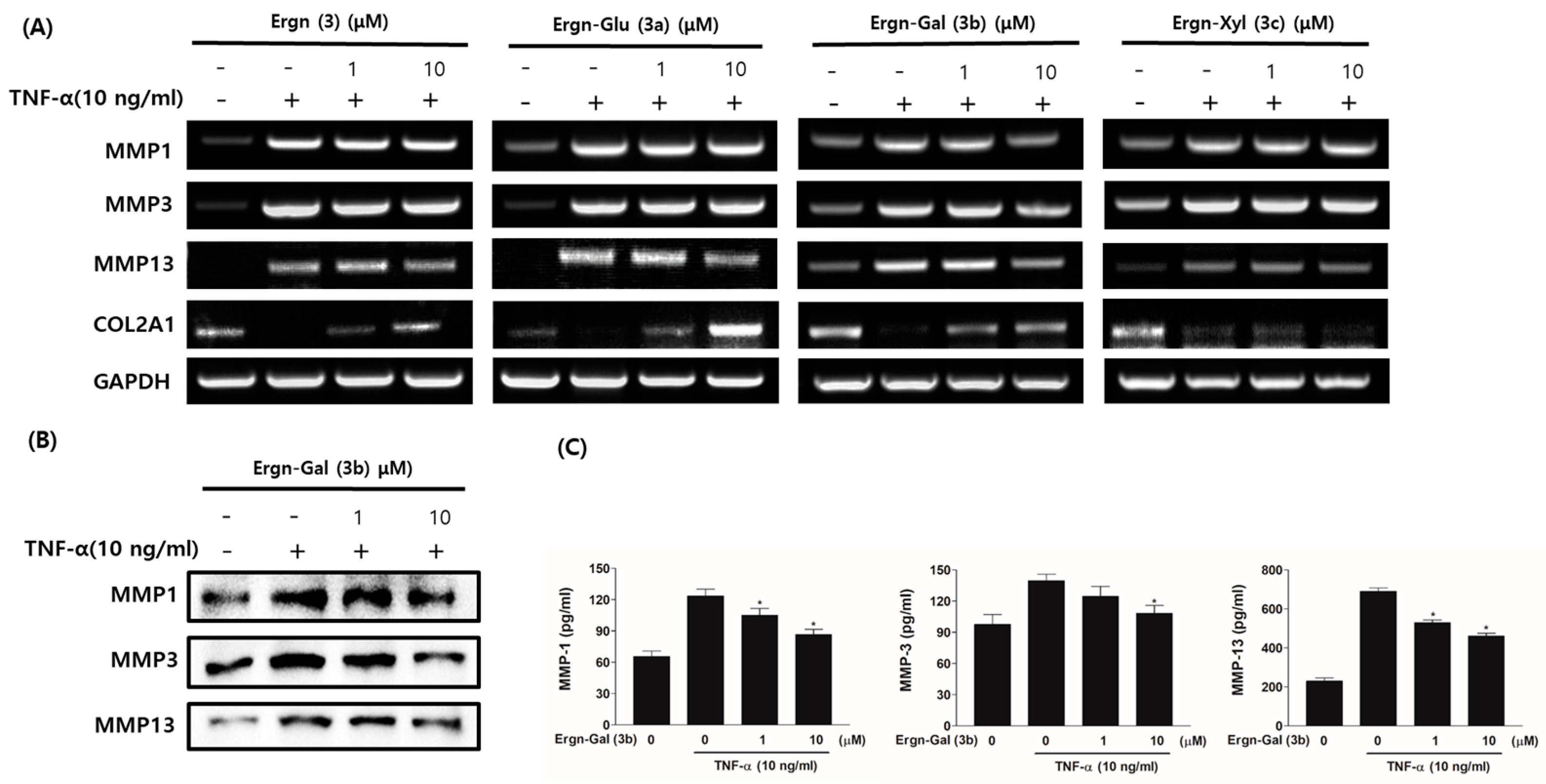
3.5. Matrix-Protective Effects of Ergostenol Glycosides in TNF-α-Induced HCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F. Aging processes and the development of osteoarthritis. Curr. Opin. Rheumatol. 2013, 25, 108–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathiessen, A.; Conaghan, P.G. Synovitis in osteoarthritis: Current understanding with therapeutic implications. Arthritis Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malemud, C.J. Matrix Metalloproteinases and Synovial Joint Pathology. Prog. Mol. Biol. Transl. Sci. 2017, 148, 305–325. [Google Scholar] [PubMed]
- Mehana, E.E.; Khafaga, A.F.; El-Blehi, S.S. The role of matrix metalloproteinases in osteoarthritis pathogenesis: An updated review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef]
- Zeng, G.Q.; Chen, A.B.; Li, W.; Song, J.H.; Gao, C.Y. High MMP-1, MMP-2, and MMP-9 protein levels in osteoarthritis. Genet. Mol. Res. 2015, 23, 14811–14822. [Google Scholar] [CrossRef]
- Fei, J.; Liang, B.; Jiang, C.; Ni, H.; Wang, L. Luteolin inhibits IL-1β-induced inflammation in rat chondrocytes and attenuates osteoarthritis progression in a rat model. Biomed. Pharmacother. 2019, 109, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Arosarena, O.A.; Barr, E.W.; Thorpe, R.; Yankey, H.; Tarr, J.T.; Safadi, F.F. Osteoactivin regulates head and neck squamous cell carcinoma invasion by modulating matrix metalloproteases. J. Cell Physiol. 2018, 23, 409–421. [Google Scholar] [CrossRef]
- Radziejewska, I.; Supruniuk, K.; Nazaruk, J.; Karna, E.; Popławska, B.; Bielawska, A.; Galicka, A. Rosmarinic acid influences collagen, MMPs, TIMPs, glycosylation and MUC1 in CRL-1739 gastric cancer cell line. Biomed. Pharmacother. 2018, 107, 397–407. [Google Scholar] [CrossRef]
- Tetlow, L.C.; Woolley, D.E. Comparative immunolocalization studies of collagenase 1 and collagenase 3 production in the rheumatoid lesion, and by human chondrocytes and synoviocytes in vitro. Br. J. Rheumatol. 1998, 37, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.C.; Martel-Pelletier, J.; Pelletier, J.P. The role of cytokines in osteoarthritis pathophysiology. Biorheology 2002, 39, 237–246. [Google Scholar]
- Rubbert-Roth, A.; Szabó, M.Z.; Kedves, M.; Nagy, G.; Atzeni, F.; Sarzi-Puttini, P. Failure of anti-TNF treatment in patients with rheumatoid arthritis: The pros and cons of the early use of alternative biological agents. Autoimmun. Rev. 2019, 18, 102398. [Google Scholar] [CrossRef]
- Tavakolpour, S.; Alesaeidi, S.; Darvishi, M.; GhasemiAdl, M.; Darabi-Monadi, S.; Akhlaghdoust, M.; Elikaei Behjati, S.; Jafarieh, A. A comprehensive review of rituximab therapy in rheumatoid arthritis patients. Clin. Rheumatol. 2019, 38, 2977–2994. [Google Scholar] [CrossRef]
- Park, H.; Lee, T.H.; Chang, F.; Kwon, H.J.; Kim, J.Y.; Kim, H. Synthesis of ergosterol and 5,6-dihydroergosterol glycosides and their inhibitory activities on lipopolysaccharide-induced nitric oxide production. Bull. Korean Chem. Soc. 2013, 34, 1330–1344. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.; Lee, T.H.; Oh, H.J.; Kim, H.; Son, Y.; Lee, E.H.; Kim, J. Inhibitory effect of 5,6-dihydroergosteol-glucoside on atopic dermatitis-like skin lesions via suppression of NF-κB and STAT activation. J. Dermatol. Sci. 2015, 79, 252–261. [Google Scholar] [CrossRef]
- Ku, Y.; Palmer, J.; Siegel, S. Stereochemistry of the palladium catalyzed exchange of deuterium with allylic hydrogens. J. Catal. 1976, 44, 449–459. [Google Scholar] [CrossRef]
- Mbadugha, B.N.S.; Menger, F.M. Sugar/Steroid/Sugar conjugates: Sensitivity of lipid binding to sugar structure. Org. Lett. 2003, 5, 4041–4044. [Google Scholar] [CrossRef]
- Sara, M.S.; Leticia, Q.C.; Jesus, S.R. Regioselective reductions of steroidal conjugated dienes by DIBAH. Tetrahedron Lett. 1995, 36, 8359–8362. [Google Scholar]
- Sepe, V.; di Leva, F.S.; D’Amore, C.; Festa, C.; de Marino, S.; Renga, B.; D’Auria, M.V.; Novellino, E.; Limongelli, V.; D’Souza, L.; et al. Marine and Semi-Synthetic Hydroxysteroids as New Scaffolds for Pregnane X Receptor Modulation. Mar. Drugs 2014, 12, 3091–3114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Lee, T.H.; Park, H.; Oh, H. Novel Ergostenol Glycoside Derivative. U.S. Patent 2019/0337976 A1, 7 November 2019. [Google Scholar]
- Yu, B.; Xie, J.; Deng, S.; Hui, Y. First synthesis of a bidesmosidic triterpene saponin by a highly efficient procedure. J. Am. Chem. Soc. 1999, 121, 12196–12197. [Google Scholar] [CrossRef]
- Zhu, Z.Y.; Yao, Q.; Liu, Y.; Si, C.L.; Chen, J.; Jiu, N.; Lian, H.Y.; Ding, L.N.; Zhang, Y.M. Highly efficient synthesis and antitumor activity of monosaccharide saponins mimicking components of Chinese folk medicine Cordyceps sinensis. J. Asian Nat. Pro. Res. 2012, 14, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Cho, Y.K.; Park, H.; Lee, T.H.; Kim, H. Comparison of the inhibitory activities of 5,6-dihydroergosterol Glycoside α- and β-anomers on skin inflammation. Molecules 2019, 24, 371. [Google Scholar] [CrossRef] [Green Version]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revision C.01 ed.; Gaussian, Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Harrison, P.; Pointon, J.J.; Chapman, K.; Roddam, A.; Wordsworth, B.P. Interleukin-1 promoter region polymorphism role in rheumatoid arthritis: A meta-analysis of IL-1B-511A/G variant reveals association with rheumatoid arthritis. Rheumatology 2008, 47, 1768–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Q.; Yan, W.; Zhao, Y.; Yu, K. Enhanced neutrophil autophagy and increased concentrations of IL-6, IL-8, IL-10 and MCP-1 in rheumatoid arthritis. Int. Immunopharmacol. 2018, 65, 119–128. [Google Scholar] [CrossRef]
- Spiller, F.; Oliveira Formiga, R.; Fernandes da Silva Coimbra, J.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q. Targeting nitric oxide as a key modulator of sepsis, arthritis and pain. Nitric Oxide 2019, 89, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.G. Therapeutic potential of COX-2 inhibitors in arthritis. Expert Opin. Investig. Drugs 2001, 10, 1317–1325. [Google Scholar] [CrossRef]
- Burrage, P.S.; Mix, K.S. Brinckerhoff CE Matrix metalloproteinases: Role in arthritis. Front. Biosci. 2006, 11, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, Y.; Nakamura, H.; Obata, K.; Yamada, H.; Hayakawa, T.; Fujikawa, K.; Okada, Y. Matrix metalloproteinases and tissue inhibitors of metalloproteinases in synovial fluids from patients with rheumatoid arthritis or osteoarthritis. Ann. Rheum. Dis. 2000, 59, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Tolboom, T.C.; Pieterman, E.; van der Laan, W.H.; Toes, R.E.; Huidekoper, A.L.; Nelissen, R.G.; Breedveld, F.C.; Huizinga, T.W. Invasive properties of fibroblast-like synoviocytes: Correlation with growth characteristics and expression of MMP-1, MMP-3, and MMP-10. Ann. Rheum. Dis. 2002, 61, 975–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, M.; Hirakawa, K.; Kato, A.; Fujii, K. The possible role of c-fos expression in rheumatoid cartilage destruction. J. Rheumatol. 2000, 27, 1606–1621. [Google Scholar]
- Dai, Q.; Zhou, D.; Xu, L.; Song, X. Curcumin alleviates rheumatoid arthritis-induced inflammation and synovial hyperplasia by targeting mTOR pathway in rats. Drug Des. Devel. Ther. 2018, 12, 4095–4105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Chang, C.C.; Yang, Y.; Yuan, L.; Xu, L.; Ho, C.T.; Li, S. Resveratrol Alleviates Rheumatoid Arthritis via Reducing ROS and Inflammation, Inhibiting MAPK Signaling Pathways, and Suppressing Angiogenesis. J. Agric. Food Chem. 2018, 66, 12953–12960. [Google Scholar] [CrossRef]
- Lee, Y.R.; Lee, J.H.; Noh, E.M.; Kim, E.K.; Song, M.Y.; Jung, W.S.; Park, S.J.; Kim, J.S.; Park, J.W.; Kwon, K.B.; et al. Guggulsterone blocks IL-1beta-mediated inflammatory responses by suppressing NF-kappaB activation in fibroblast-like synoviocytes. Life Sci. 2008, 82, 1203–1209. [Google Scholar] [CrossRef]
- Yan, X.; Huang, G.; Liu, Q.; Zheng, J.; Chen, H.; Huang, Q.; Chen, J.; Huang, H. Withaferin A protects against spinal cord injury by inhibiting apoptosis and inflammation in mice. Pharm. Biol. 2017, 55, 1171–1176. [Google Scholar] [CrossRef]
- Villalvilla, A.; da Silva, J.A.; Largo, R.; Gualillo, O.; Vieira, P.C.; Herrero-Beaumont, G.; Gómez, R. 6-Shogaol inhibits chondrocytes’ innate immune responses and cathepsin-K activity. Mol. Nutr. Food Res. 2014, 58, 256–266. [Google Scholar] [CrossRef]
- Wang, Q.; Pan, X.; Wong, H.H.; Wagner, C.A.; Lahey, L.J.; Robinson, W.H.; Sokolove, J. Oral and topical boswellic acid attenuates mouse osteoarthritis. Osteoarthr. Cartil. 2014, 22, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Lu, J.; Li, W.; Wu, A.; Zhang, X.; Tong, W.; Ho, K.K.; Qin, L.; Song, H.; Mak, K.K. Reciprocal inhibition of YAP/TAZ and NF-kappaB regulates osteoarthritic cartilage degradation. Nat. Commun. 2018, 9, 4564. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Size (bp) | |
|---|---|---|---|
| MMP-1 | sense antisense | CTGTTCAGGACAGAATGTG TTGGACTCACACCATGTGTT | 395 |
| MMP-3 | sense antisense | TGCGTGGCAGTTTGCTCAGC GAATGTGAGTGGAGTCACCT | 573 |
| MMP-13 | sense antisense | GGCTCCGAGAAATGCAGTCT ATCAAATGGGTAGAAGTCGC | 273 |
| COL2A1 | sense antisense | AATCCAGCAAACGTTCCCAA CGGTACTCGATAACAGTCAA | 380 |
| IL-1β | sense antisense | ATGTACCAGTTGGGGAACTG GGATATGGAGCAACAAGTGG | 264 |
| IL-6 | sense antisense | TGACAAACAAATTCGGTACATCC ATCTGAGGTGCCCATGCTAC | 497 |
| gIL-8 | sense antisense | ATGACTTCCAAGCTGGCCGTGGCT TCTCAGCCCTCTTCAAAAACTTCTC | 292 |
| INOS | sense antisense | GGCCTCTCAGCTCACCCCGA CCAGGCGCACTGTCTGGTGG | 496 |
| GAPDH | sense antisense | AGGTCGGAGTCAACGGATTTGG ACAGTCTTCTGGGTGGCAGT | 226 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, H.; Ko, M.; Park, Y.; Kim, J.; Yoon, D.; Lee, E.; Lee, T.; Kim, H. Δ8(14)-Ergostenol Glycoside Derivatives Inhibit the Expression of Inflammatory Mediators and Matrix Metalloproteinase. Molecules 2021, 26, 4547. https://doi.org/10.3390/molecules26154547
Moon H, Ko M, Park Y, Kim J, Yoon D, Lee E, Lee T, Kim H. Δ8(14)-Ergostenol Glycoside Derivatives Inhibit the Expression of Inflammatory Mediators and Matrix Metalloproteinase. Molecules. 2021; 26(15):4547. https://doi.org/10.3390/molecules26154547
Chicago/Turabian StyleMoon, Hyejin, Myoungsil Ko, Yujin Park, Jeonguk Kim, Dowon Yoon, Eunjoohwang Lee, Taehoon Lee, and Hakwon Kim. 2021. "Δ8(14)-Ergostenol Glycoside Derivatives Inhibit the Expression of Inflammatory Mediators and Matrix Metalloproteinase" Molecules 26, no. 15: 4547. https://doi.org/10.3390/molecules26154547
APA StyleMoon, H., Ko, M., Park, Y., Kim, J., Yoon, D., Lee, E., Lee, T., & Kim, H. (2021). Δ8(14)-Ergostenol Glycoside Derivatives Inhibit the Expression of Inflammatory Mediators and Matrix Metalloproteinase. Molecules, 26(15), 4547. https://doi.org/10.3390/molecules26154547