
Evaluation of Cocaine Effect on Endogenous Metabolites of HepG2 Cells Using Targeted Metabolomics

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
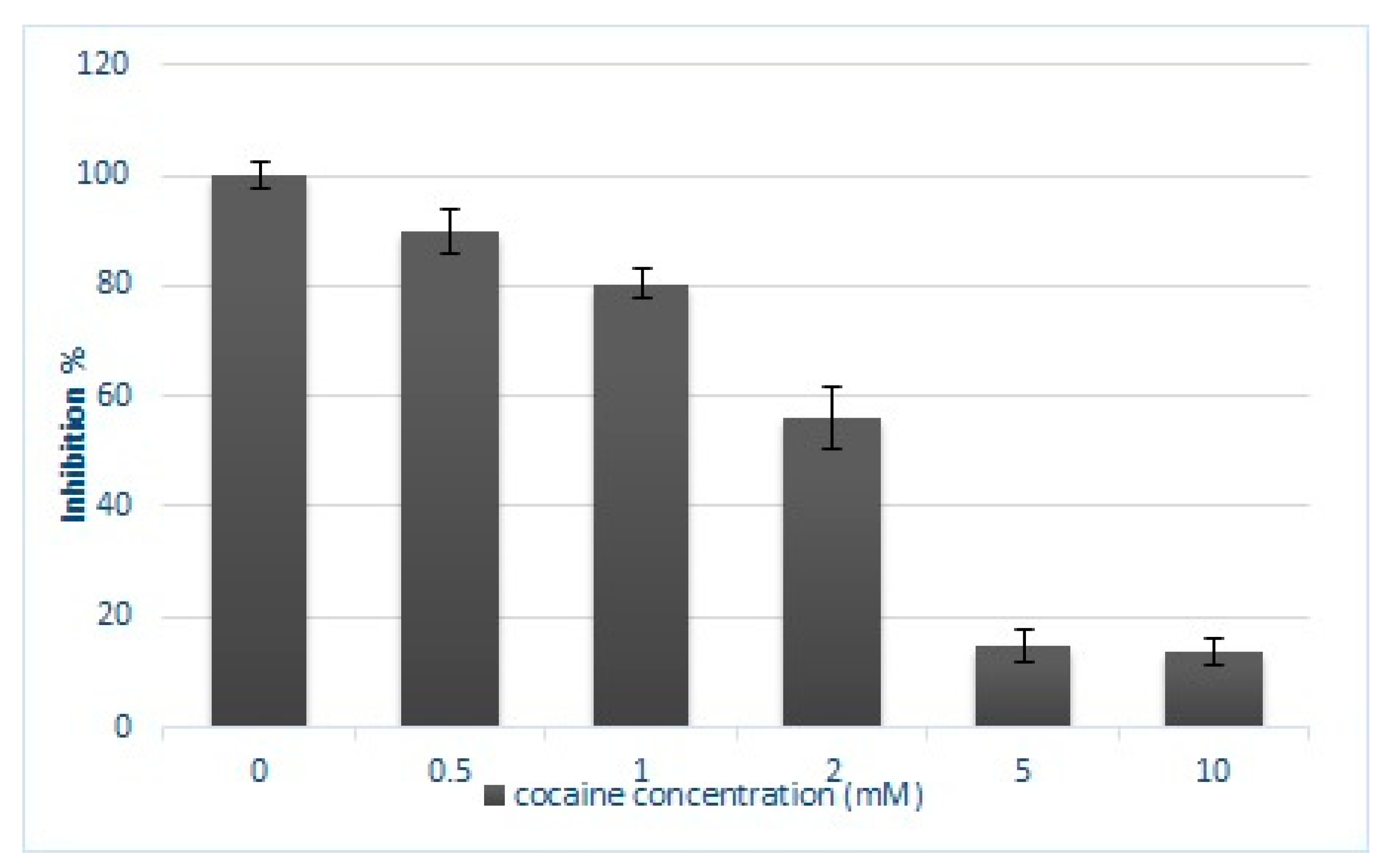
2.1. XTT Cell Proliferation Assay
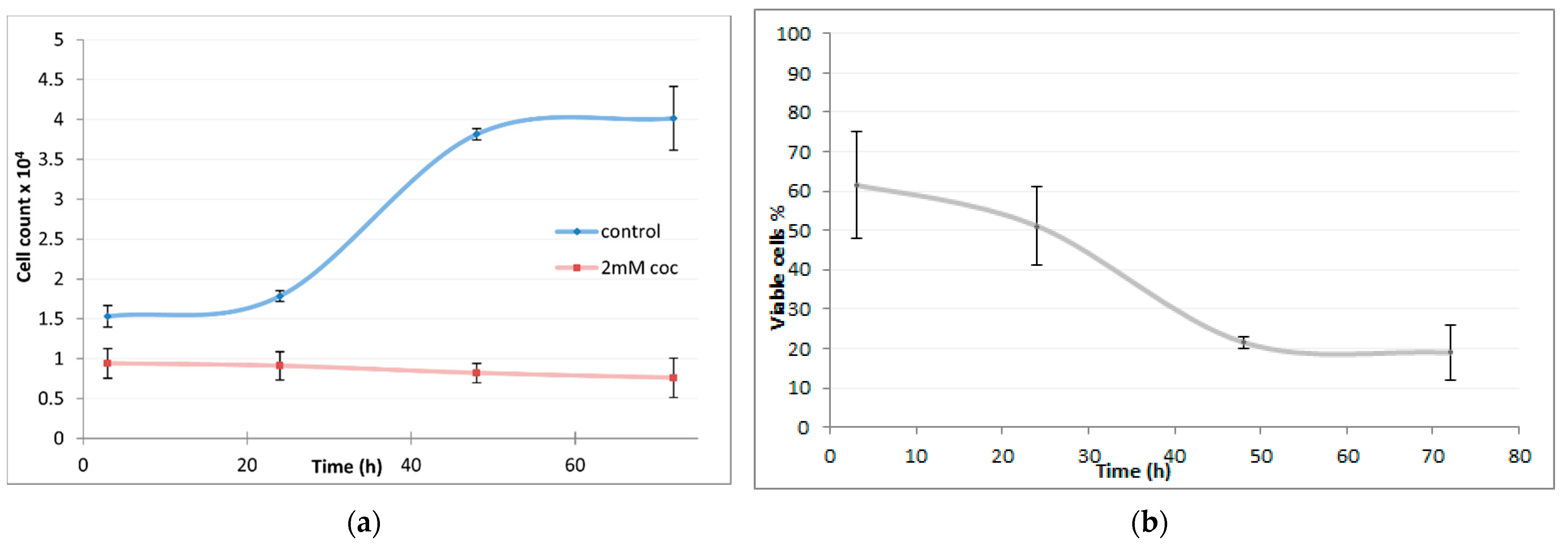
2.2. Cell Growth Rate
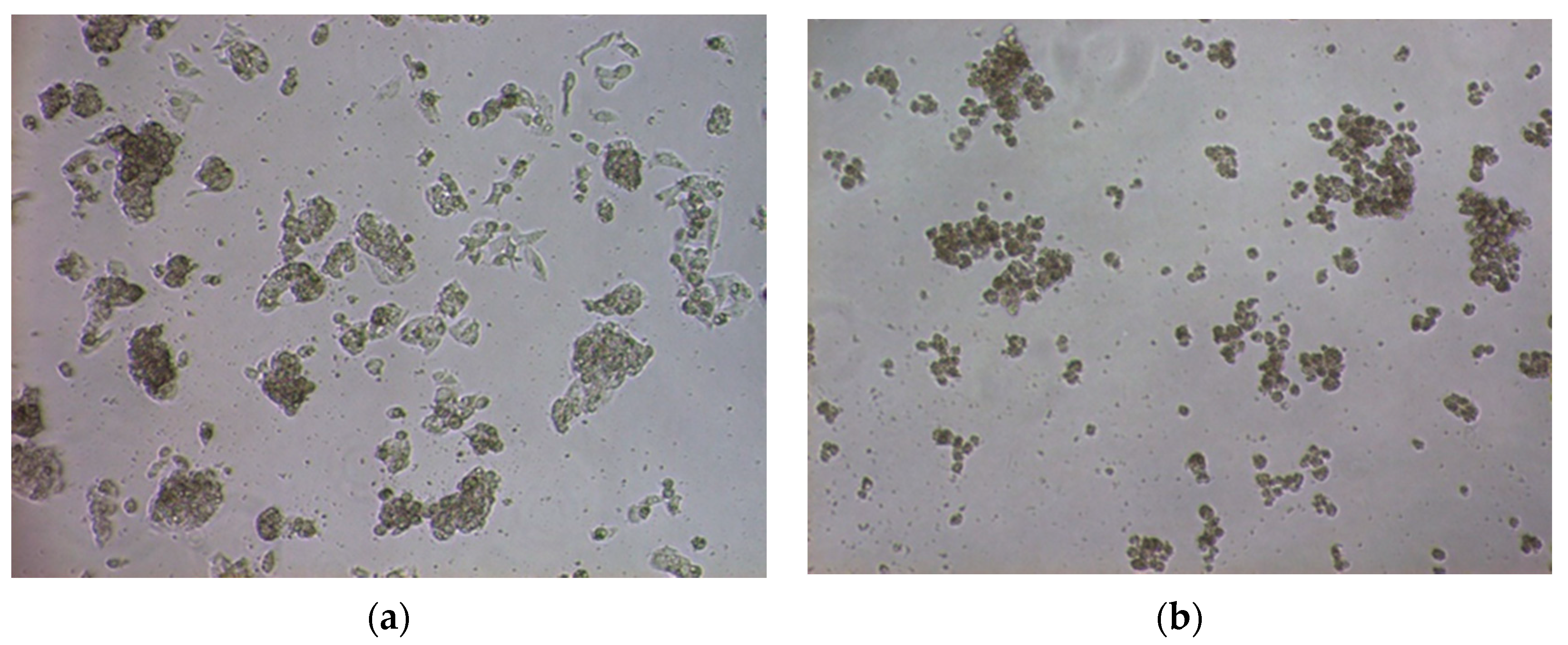
2.3. Microscopic View of HepG2
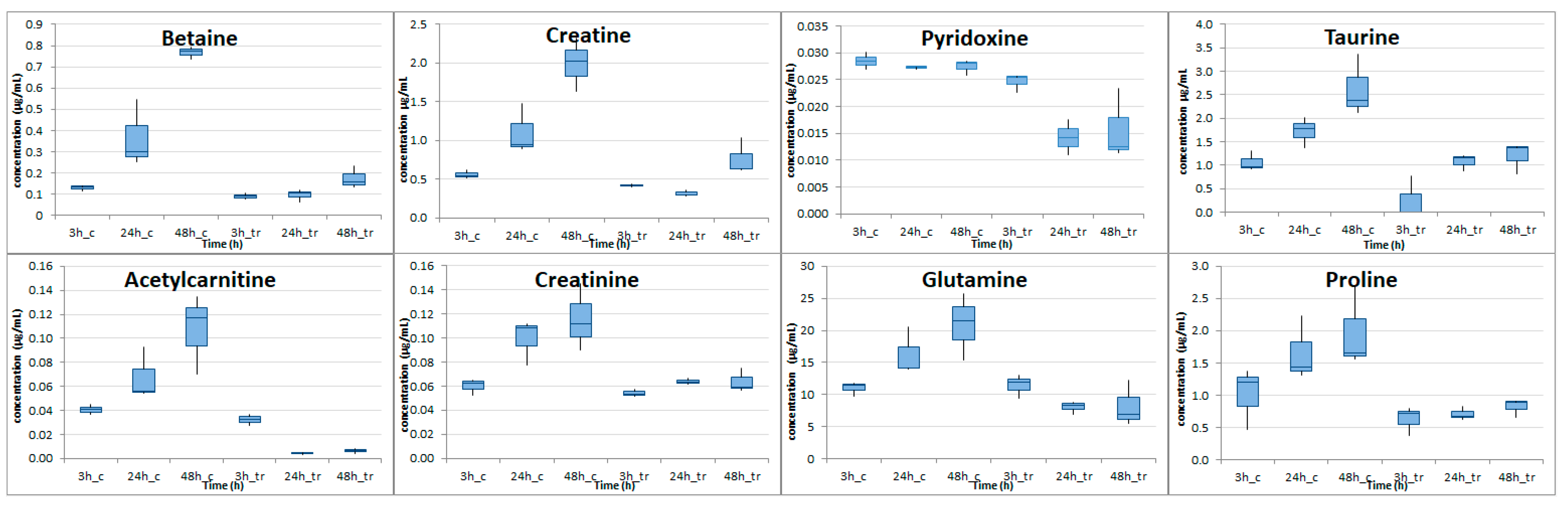
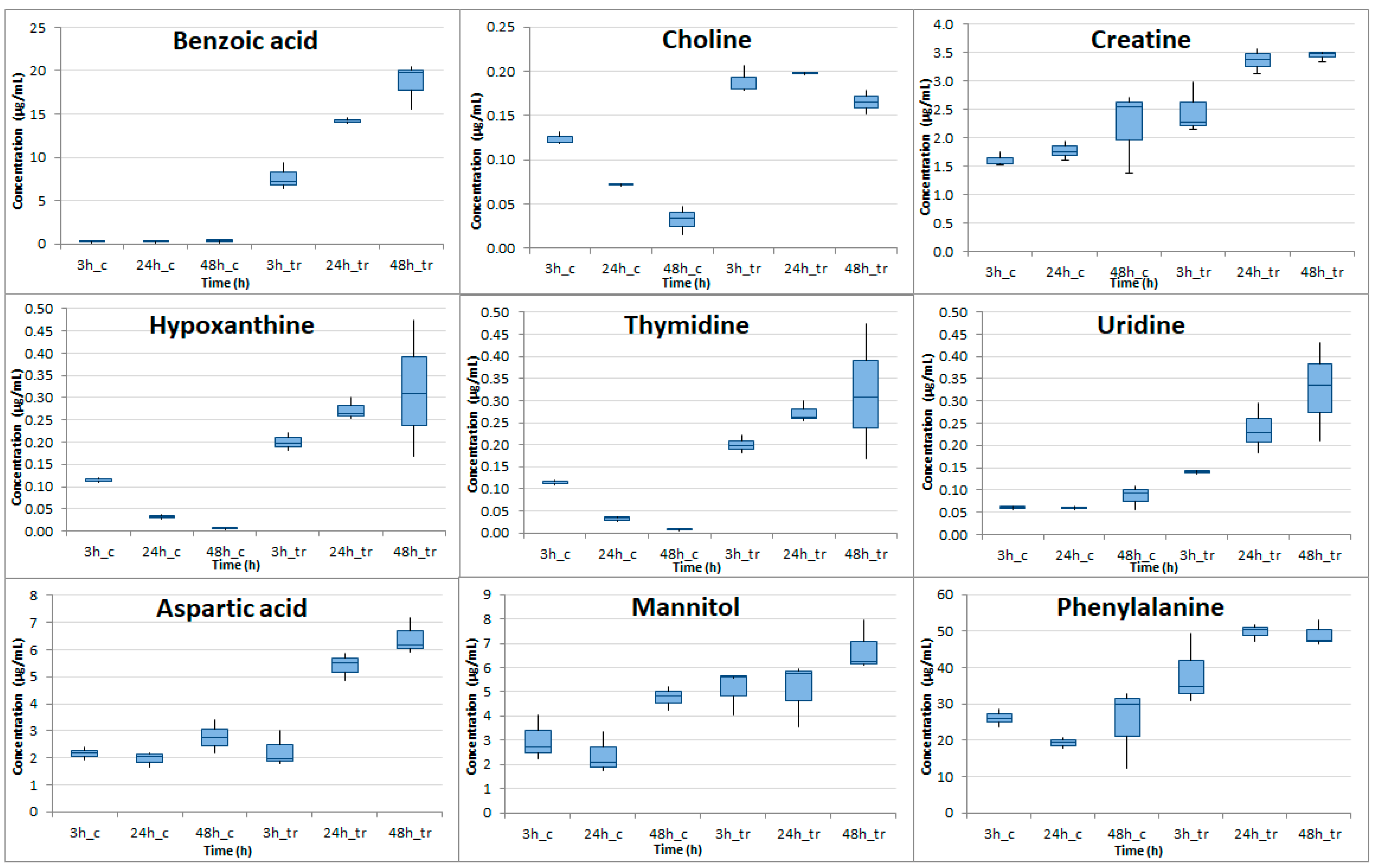
2.4. Targeted Metabolic Profiling Analysis
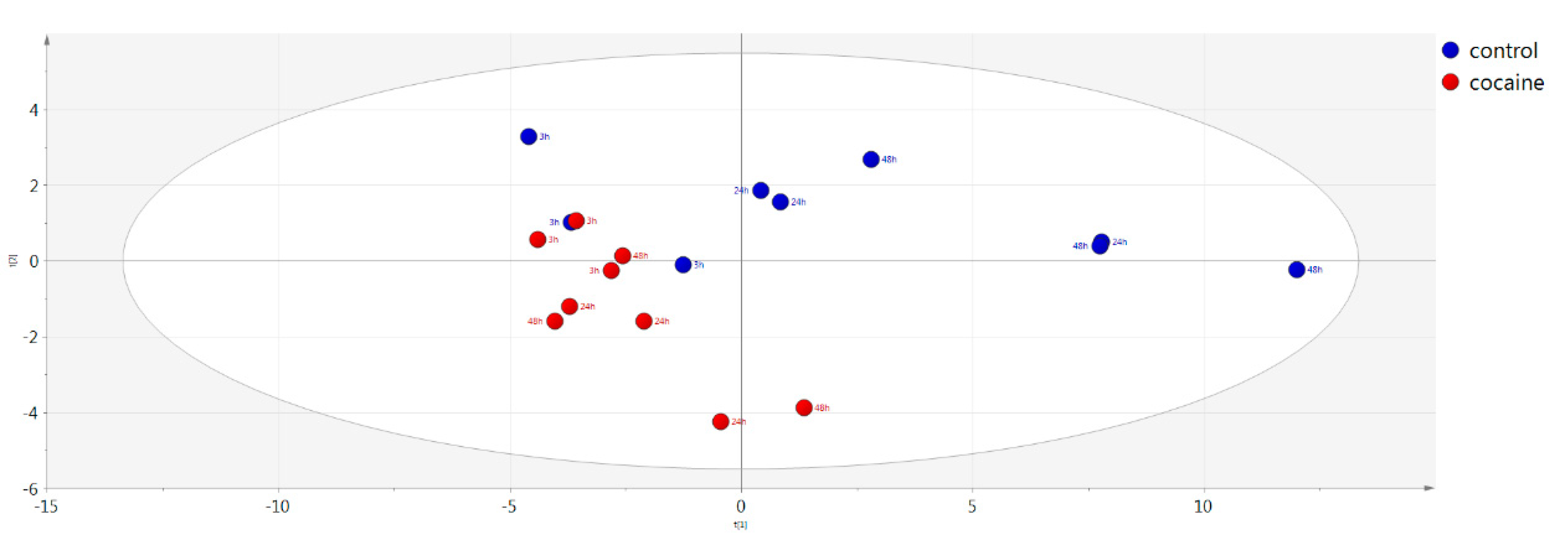
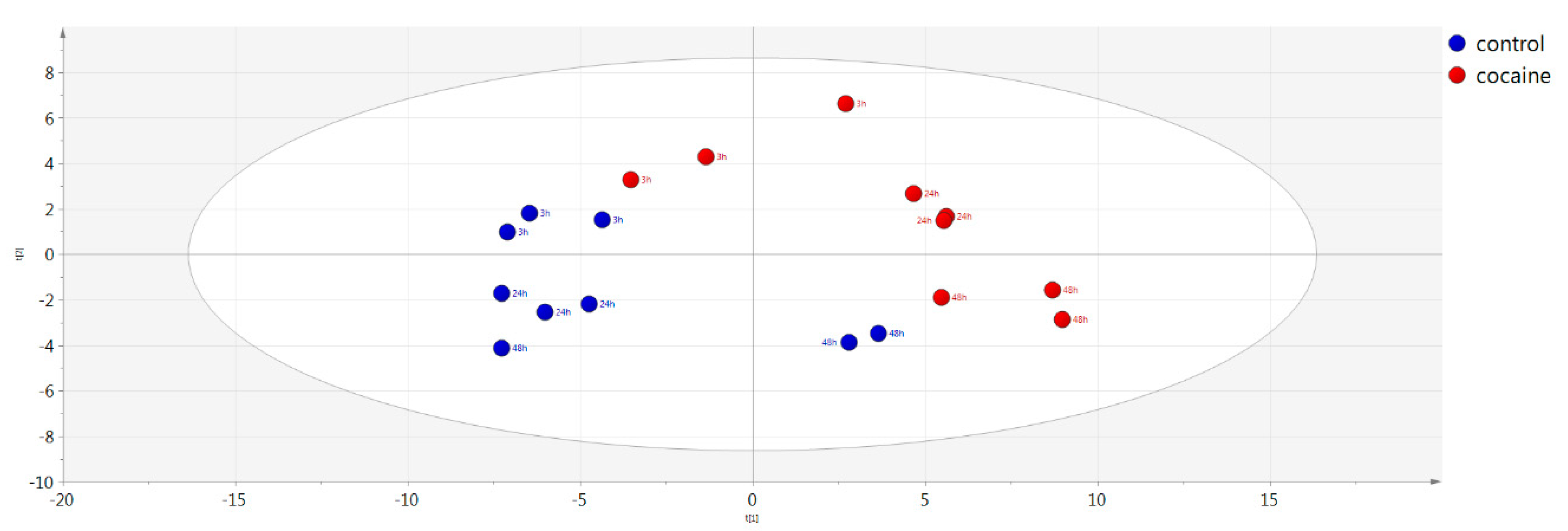
2.5. Multivariate Analysis
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture Growth
4.3. XTT Assay
4.4. Cell Growth Rate
4.5. Metabolic Profiling Analysis
4.6. Sample Preparation
4.7. LC-MS/MS Analysis
4.8. Data Handling
Supplementary Materials
Author Contributions
Funding

Acknowledgments
Conflicts of Interest
Sample Availability
References
- Schindler, C.W.; Goldberg, S.R. Accelerating cocaine metabolism as an approach to the treatment of cocaine abuse and toxicity. Future Med. Chem. 2012, 4, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Kolbrich, E.A.; Barnes, A.J.; Gorelick, D.A.; Boyd, S.J.; Cone, E.J.; Huestis, M.A. Major and minor metabolites of cocaine in human plasma following controlled subcutaneous cocaine administration. J. Anal. Toxicol. 2006, 30, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziani, M.; Antonilli, L.; Togna, A.R.; Grassi, M.C.; Badiani, A.; Saso, L. Cardiovascular and Hepatic Toxicity of Cocaine: Potential Beneficial Effects of Modulators of Oxidative Stress. Oxidative Med. Cell. Longev. 2016, 2016, 8408479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanel, G.C.; Cassidy, W.; Shuster, L.; Reynolds, T.B. Cocaine-induced liver cell injury: Comparison of morphological features in man and in experimental models. Hepatology 1990, 11, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Dinis-Oliveira, R.J. Metabolomics of cocaine: Implications in toxicity. Toxicol. Mech. Methods 2015, 25, 494–500. [Google Scholar] [PubMed]
- Valente, M.; Carvalho, M.; Bastos, M.D.L.; de Pinho, P.G. Contribution of Oxidative Metabolism to Cocaine-Induced Liver and Kidney Damage. Curr. Med. Chem. 2012, 19, 5601–5606. [Google Scholar] [CrossRef]
- Dwyer, T.M. Chemical Signaling in the Nervous System. In Fundamental Neuroscience for Basic and Clinical Applications, 5th ed.; Elsevier: Philadelphia, PA, USA, 2018; pp. 54–71.e1. ISBN 978-0-323-39632-5. [Google Scholar]
- Steinmetz, A.; Steffens, L.; Morás, A.M.; Prezzi, F.; Braganhol, E.; Saffi, J.; Ortiz, R.S.; Barros, H.M.; Moura, D.J. In vitro model to study cocaine and its contaminants. Chem. Interact. 2018, 285, 1–7. [Google Scholar] [CrossRef]
- Zaitsu, K.; Hayashi, Y.; Kusano, M.; Tsuchihashi, H.; Ishii, A. Application of metabolomics to toxicology of drugs of abuse: A mini review of metabolomics approach to acute and chronic toxicity studies. Drug Metab. Pharmacokinet. 2016, 31, 21–26. [Google Scholar] [CrossRef]
- Alves, M.N.R.; Zanchetti, G.; Piccinotti, A.; Tameni, S.; De Martinis, B.S.; Polettini, A. Determination of cocaine and metabolites in hair by column-switching LC-MS-MS analysis. Anal. Bioanal. Chem. 2013, 405, 6299–6306. [Google Scholar] [CrossRef]
- Janicka, M.; Kot-Wasik, A.; Namiesnik, J. Analytical procedures for determination of cocaine and its metabolites in biological samples. TrAC Trends Anal. Chem. 2010, 29, 209–224. [Google Scholar] [CrossRef]
- Dulaurent, S.; EL Balkhi, S.; Poncelet, L.; Gaulier, J.-M.; Marquet, P.; Saint-Marcoux, F. QuEChERS sample preparation prior to LC-MS/MS determination of opiates, amphetamines, and cocaine metabolites in whole blood. Anal. Bioanal. Chem. 2016, 408, 1467–1474. [Google Scholar] [CrossRef]
- Barroso, M.; Gallardo, E.; Queiroz, J.; Queiroz, J. Bioanalytical methods for the determination of cocaine and metabolites in human biological samples. Bioanalysis 2009, 1, 977–1000. [Google Scholar] [CrossRef]
- Pego, A.; Roveri, F.; Kuninari, R.; Leyton, V.; Miziara, I.; Yonamine, M. Determination of cocaine and its derivatives in hair samples by liquid phase microextraction (LPME) and gas chromatography–mass spectrometry (GC–MS). Forensic Sci. Int. 2017, 274, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Orfanidis, A.; Gika, H.; Mastrogianni, O.; Krokos, A.; Theodoridis, G.; Zaggelidou, E.; Raikos, N. Determination of drugs of abuse and pharmaceuticals in skeletal tissue by UHPLC–MS/MS. Forensic Sci. Int. 2018, 290, 137–145. [Google Scholar] [CrossRef]
- Orfanidis, A.; Mastrogianni, O.; Koukou, A.; Psarros, G.; Gika, H.; Theodoridis, G.; Raikos, N. A GC–MS method for the detection and quantitation of ten major drugs of abuse in human hair samples. J. Chromatogr. B 2017, 1047, 141–150. [Google Scholar] [CrossRef]
- Alvear, E.; von Baer, D.; Mardones, C.; Hitschfeld, A. Determination of cocaine and its major metabolite benzoylecgonine in several matrices obtained from deceased individuals with presumed drug consumption prior to death. J. Forensic Leg. Med. 2014, 23, 37–43. [Google Scholar] [CrossRef]
- Marcos, A.; Moreno, M.; Orihuel, J.; Ucha, M.; De Paz, A.M.; Higuera-Matas, A.; Capellán, R.; Crego, A.L.; Martínez-Larrañaga, M.-R.; Ambrosio, E.; et al. The effects of combined intravenous cocaine and ethanol self-administration on the behavioral and amino acid profile of young adult rats. PLoS ONE 2020, 15, e0227044. [Google Scholar] [CrossRef]
- Ghanbari, R.; Sumner, S. Using Metabolomics to Investigate Biomarkers of Drug Addiction. Trends Mol. Med. 2018, 24, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Theodoridis, G.; Gika, H.G.; Wilson, I.D. LC-MS-based methodology for global metabolite profiling in metabonomics/metabolomics. TrAC Trends Anal. Chem. 2008, 27, 251–260. [Google Scholar] [CrossRef]
- Theodoridis, G.A.; Gika, H.G.; Want, E.J.; Wilson, I. Liquid chromatography–mass spectrometry based global metabolite profiling: A review. Anal. Chim. Acta 2012, 711, 7–16. [Google Scholar] [CrossRef]
- Theodoridis, G.; Gika, H.G.; Wilson, I. Mass spectrometry-based holistic analytical approaches for metabolite profiling in systems biology studies. Mass Spectrom. Rev. 2011, 30, 884–906. [Google Scholar] [CrossRef]
- Heard, K.; Palmer, R.; Zahniser, N.R. Mechanisms of Acute Cocaine Toxicity. Open Pharmacol. J. 2008, 2, 70–78. [Google Scholar] [CrossRef]
- Zaitsu, K.; Miyawaki, I.; Bando, K.; Horie, H.; Shima, N.; Katagi, M.; Tatsuno, M.; Bamba, T.; Sato, T.; Ishii, A.; et al. Metabolic profiling of urine and blood plasma in rat models of drug addiction on the basis of morphine, methamphetamine, and cocaine-induced conditioned place preference. Anal. Bioanal. Chem. 2013, 406, 1339–1354. [Google Scholar] [CrossRef]
- Patkar, A.A.; Rozen, S.G.; Mannelli, P.; Matson, W.; Pae, C.-U.; Krishnan, K.R.; Kaddurah-Daouk, R. Alterations in tryptophan and purine metabolism in cocaine addiction: A metabolomic study. Psychopharmacology 2009, 206, 479–489. [Google Scholar] [CrossRef]
- Li, Y.; Yan, G.-Y.; Zhou, J.-Q.; Bu, Q.; Deng, P.-C.; Yang, Y.-Z.; Lv, L.; Deng, Y.; Zhao, J.-X.; Shao, X.; et al. 1H NMR-based metabonomics in brain nucleus accumbens and striatum following repeated cocaine treatment in rats. Neuroscience 2012, 218, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Yao, D.; Gosnell, B.A.; Chen, C. Lipidomic profiling reveals protective function of fatty acid oxidation in cocaine-induced hepatotoxicity. J. Lipid Res. 2012, 53, 2318–2330. [Google Scholar] [CrossRef] [Green Version]
- Virgiliou, C.; Sampsonidis, I.; Gika, H.G.; Raikos, N.; Theodoridis, G.A. Development and validation of a HILIC-MS/MS multitargeted method for metabolomics applications. Electrophoresis 2015, 36, 2215–2225. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Li, J.; Li, W.; Altura, B.T.; Altura, B. Cocaine induces apoptosis in cerebral vascular muscle cells: Potential roles in strokes and brain damage. Eur. J. Pharmacol. 2003, 482, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Falk, P.; Sabater, R.; Carballo, D. Response of the human hepatic tissue cultures HEP-G2 and WRL-68 to cocaine. J. Pharmacol. Toxicol. Methods 1995, 33, 113–120. [Google Scholar] [CrossRef]
- Deda, O.; Virgiliou, C.; Orfanidis, A.; Gika, H.G. Study of Fecal and Urinary Metabolite Perturbations Induced by Chronic Ethanol Treatment in Mice by UHPLC-MS/MS Targeted Profiling. Metabolites 2019, 9, 232. [Google Scholar] [CrossRef] [Green Version]
- Deda, O.; Virgiliou, C.; Armitage, E.G.; Orfanidis, A.; Taitzoglou, I.; Wilson, I.D.; Loftus, N.; Gika, H.G. Metabolic Phenotyping Study of Mouse Brains Following Acute or Chronic Exposures to Ethanol. J. Proteome Res. 2020, 19, 4071–4081. [Google Scholar] [CrossRef]
- Jover, R.; Ponsoda, X.; Castell, J.V.; Gómez-Lechón, M.J. Hepatotoxicity of Opiates and Cocaine on Different Hepatic Cellular Systems. Altern. Lab. Anim. 1990, 17, 240–245. [Google Scholar] [CrossRef]
- Cunha-Oliveira, T.; Rego, A.C.; Cardoso, S.M.; Borges, F.; Swerdlow, R.H.; Macedo, T.; De Oliveira, C.R. Mitochondrial dysfunction and caspase activation in rat cortical neurons treated with cocaine or amphetamine. Brain Res. 2006, 1089, 44–54. [Google Scholar] [CrossRef]
- Badisa, R.B.; Batton, C.S.; Mazzio, E.; Grant, S.C.; Goodman, C.B. Identification of biochemical and cytotoxic markers in cocaine treated PC12 cells. Sci. Rep. 2018, 8, 2710. [Google Scholar] [CrossRef]
- Badisa, R.B.; Darling-Reed, S.F.; Goodman, C.B. Cocaine Induces Alterations in Mitochondrial Membrane Potential and Dual Cell Cycle Arrest in Rat C6 Astroglioma Cells. Neurochem. Res. 2009, 35, 288–297. [Google Scholar] [CrossRef] [Green Version]
- Curry, S.H.; Coombs, S.E. Benzoic acid is not the only important product of accelerated metabolism of cocaine. Proc. Natl. Acad. Sci. USA 2016, 113, E2101. [Google Scholar] [CrossRef] [Green Version]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, K.A.; Chiu, V.M.; Lukus, P.A.; Zhang, X.; Siems, W.F.; Schenk, J.O.; Hill, H.H. Neuronal metabolomics by ion mobility mass spectrometry: Cocaine effects on glucose and selected biogenic amine metabolites in the frontal cortex, striatum, and thalamus of the rat. Anal. Bioanal. Chem. 2013, 405, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, E.; Marcos, A.; Ambrosio, E.; Mayboroda, O.; Marina, M.L.; Crego, A.L. Investigation on the combined effect of cocaine and ethanol administration through a liquid chromatography–mass spectrometry metabolomics approach. J. Pharm. Biomed. Anal. 2017, 140, 313–321. [Google Scholar] [CrossRef]
- Yablonsky-Alter, E.; Agovic, M.S.; Gashi, E.; Lidsky, T.I.; Friedman, E.; Banerjee, S.P. Cocaine challenge enhances release of neuroprotective amino acid taurine in the striatum of chronic cocaine treated rats: A microdialysis study. Brain Res. Bull. 2009, 79, 215–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotantaki, P.; Mosialos, G. The expression of tumor suppressor gene Cyldis upregulated by histone deacetylace inhibitors in human hepatocellular carcinoma cell lines. Cell Biochem. Funct. 2016, 34, 465–468. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (μg/mL) | Cocaine (μg/mL) | p-Value | Log2FC (Cocaine vs. Control) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | Compound | 3 h | 24 h | 48 h | 3 h | 24 h | 48 h | 3 h | 24 h | 48 h | 3 h | 24 h | 48 h |
| 1 | Betaine | 0.129 | 0.368 | 0.768 | 0.090 | 0.096 | 0.175 | 3 × 10−2 | 4 10−2 | 7 × 10−5 | −0.51 | −1.93 | −2.14 |
| 2 | Hypotaurine | 6.619 | 11.14 | 12.59 | 2.592 | 5.821 | 3.796 | 8 × 10−2 | 10−1 | 6 × 10−3 | −1.35 | −0.94 | −1.73 |
| 3 | Acetylcarnitine | 0.041 | 0.068 | 0.107 | 0.032 | 0.004 | 0.006 | 9 × 10−2 | 7 × 10−3 | 6 × 10−3 | −0.33 | −4.08 | −4.12 |
| 4 | Creatine | 0.560 | 1.107 | 1.994 | 0.419 | 0.314 | 0.763 | 9 × 10−3 | 10−2 | 7 × 10−3 | −0.42 | −1.82 | −1.39 |
| 5 | Acetyl aspartate | 0.826 | 2.946 | 3.929 | 0.678 | 0.794 | 0.940 | 7 × 10−1 | 2 × 10−1 | 10−2 | −0.28 | −1.89 | −2.06 |
| 6 | Serine | 1.886 | 2.469 | 4.012 | 1.449 | 3.025 | 0.584 | 6 × 10−1 | 8 × 10−1 | 10−2 | −0.38 | 0.29 | −2.78 |
| 7 | Glutamine | 11.02 | 16.29 | 20.88 | 11.44 | 8.056 | 8.182 | 8 × 10−1 | 2 × 10−2 | 2 ×10−2 | 0.05 | −1.02 | −1.35 |
| 8 | Taurine | 1.064 | 1.717 | 2.619 | 0.699 | 1.103 | 1.220 | 5 × 10−2 | 5 × 10−2 | 3 × 10−2 | −0.61 | −0.64 | −1.10 |
| 9 | Alanine | 1.534 | 2.447 | 3.840 | 1.599 | 1.846 | 1.649 | 8 × 10−1 | 2 × 10−1 | 3 × 10−2 | 0.06 | −0.41 | −1.22 |
| 10 | Proline | 1.010 | 1.661 | 1.972 | 0.632 | 0.712 | 0.818 | 3 × 10−1 | 3 × 10−2 | 4 × 10−2 | −0.68 | −1.22 | −1.27 |
| 11 | Anthranilic acid | 0.440 | 1.527 | 1.959 | 0.212 | 0.552 | 0.314 | 5 × 10−1 | 2 × 10−1 | 4 × 10−2 | −1.06 | −1.47 | −2.64 |
| 12 | Creatinine | 0.060 | 0.099 | 0.116 | 0.054 | 0.064 | 0.064 | 2 × 10−1 | 3 × 10−2 | 4 × 10−2 | −0.16 | −0.63 | −0.86 |
| 13 | Pyridoxine | 0.028 | 0.027 | 0.027 | 0.025 | 0.014 | 0.016 | 4 × 10−2 | 3 × 10−3 | 4 × 10−2 | −0.21 | −0.93 | −0.80 |
| 14 | Glutamic acid | 8.255 | 15.979 | 29.92 | 7.682 | 8.928 | 11.21 | 7 × 10−1 | 10−1 | 4 × 10−2 | −0.10 | −0.84 | −1.42 |
| 15 | Pantothenate | 0.148 | 0.196 | 0.378 | 0.210 | 0.135 | 0.152 | 5 × 10−2 | 8 × 10−2 | 5 × 10−2 | 0.50 | −0.54 | −1.31 |
| 16 | Aspartic acid | 3.073 | 4.953 | 6.476 | 2.935 | 3.503 | 2.213 | 7 × 10−1 | 5 × 10−1 | 6 × 10−2 | −0.07 | −0.50 | −1.55 |
| 17 | Uracil | 0.029 | 0.043 | 0.046 | 0.015 | 0.027 | 0.021 | 8 × 10−2 | 8 × 10−2 | 8 × 10−2 | −0.93 | −0.63 | −1.11 |
| 18 | Choline | 0.003 | 0.009 | 0.007 | 0.006 | 0.005 | 0.004 | 4 × 10−3 | 10−1 | 8 × 10−2 | 1.25 | −0.79 | −0.92 |
| 19 | Isoleucine | 0.315 | 0.858 | 1.148 | 0.384 | 0.416 | 0.612 | 8 × 10−1 | 9 × 10−2 | 8 × 10−2 | 0.28 | −1.04 | −0.91 |
| 20 | Benzoic acid | 0.417 | 0.462 | 0.428 | 0.386 | 0.339 | 0.604 | 8 × 10−1 | 3 × 10−1 | 9 × 10−2 | −0.11 | −0.45 | 0.50 |
| 21 | leucine | 0.577 | 1.283 | 1.466 | 0.665 | 0.861 | 0.887 | 6 × 10−1 | 3 × 10−1 | 10−1 | 0.20 | −0.58 | −0.73 |
| 22 | Threonine | 1.818 | 2.626 | 3.498 | 1.901 | 2.246 | 2.115 | 8 × 10−1 | 7 × 10−1 | 10−1 | 0.06 | −0.23 | −0.73 |
| 23 | Cytosine | 0.047 | 0.047 | 0.046 | 0.046 | 0.046 | 0.046 | 5 × 10−1 | 4 × 10−1 | 10−1 | −0.03 | −0.03 | −0.01 |
| 24 | Adenine | 0.018 | 0.022 | 0.025 | 0.021 | 0.021 | 0.021 | 5 × 10−3 | 7 × 10−1 | 10−1 | 0.18 | −0.06 | −0.26 |
| 25 | Nicotinamide | 0.008 | 0.061 | 0.084 | 0.010 | 0.041 | 0.036 | 5 × 10−1 | 4 × 10−1 | 10−1 | 0.36 | −0.55 | −1.24 |
| 26 | Asparagine | 0.938 | 0.961 | 0.935 | 0.900 | 0.926 | 0.854 | 6 × 10−1 | 7 × 10−1 | 10−1 | −0.06 | −0.05 | −0.13 |
| 27 | Thymidine | 0.007 | 0.009 | 0.007 | 0.004 | 0.003 | 0.004 | 5 × 10−2 | 4 × 10−2 | 10−1 | −0.91 | −1.53 | −0.76 |
| 28 | Tyrosine | 0.958 | 2.024 | 2.663 | 1.326 | 1.542 | 2.146 | 3 × 10−1 | 2 × 10−1 | 2 × 10−1 | 0.47 | −0.39 | −0.31 |
| 29 | Pyroglutamic | 2.749 | 3.503 | 5.955 | 2.114 | 4.744 | 3.479 | 7 × 10−1 | 4 × 10−1 | 2 × 10−1 | −0.38 | 0.44 | −0.78 |
| 30 | Norvaline-Valine | 1.110 | 1.625 | 1.936 | 1.155 | 1.401 | 1.407 | 9 × 10−1 | 4 × 10−1 | 3 × 10−1 | 0.06 | −0.21 | −0.46 |
| 31 | Glycine | 1.307 | 2.570 | 4.963 | 1.350 | 2.064 | 3.606 | 9 × 10−1 | 3 × 10−1 | 3 × 10−1 | 0.05 | −0.32 | −0.46 |
| 32 | Methionine | 0.148 | 0.338 | 0.400 | 0.155 | 0.159 | 0.250 | 9 × 10−1 | 9 × 10−2 | 3 × 10−1 | 0.07 | −1.09 | −0.68 |
| 33 | Phenylalanine | 1.058 | 1.506 | 1.455 | 1.094 | 1.325 | 1.276 | 8 × 10−1 | 3 × 10−1 | 4 × 10−1 | 0.05 | −0.19 | −0.19 |
| 34 | Sorbitol | 1.629 | 2.422 | 3.273 | 2.752 | 1.676 | 1.980 | 4 × 10−1 | 4 × 10−1 | 4 × 10−1 | 0.76 | −0.53 | −0.73 |
| 35 | Lactic acid | 13.46 | 15.17 | 14.10 | 8.273 | 14.62 | 17.44 | 10−1 | 9 × 10−1 | 5 × 10−1 | −0.70 | −0.05 | 0.31 |
| 36 | Lysine | 1.270 | 1.713 | 1.981 | 1.214 | 1.430 | 1.667 | 4 × 10−1 | 3 × 10−1 | 5 × 10−1 | −0.07 | −0.26 | −0.25 |
| 37 | Thiamine | 0.085 | 0.091 | 0.091 | 0.083 | 0.087 | 0.088 | 10−1 | 2 × 10−1 | 5 × 10−1 | −0.03 | −0.06 | −0.05 |
| 38 | Tryptophan | 0.803 | 0.814 | 0.938 | 0.806 | 0.831 | 0.850 | 9 × 10−1 | 6 × 10−1 | 5 × 10−1 | 0.01 | 0.03 | −0.14 |
| 39 | Ornithine | 1.194 | 1.184 | 1.270 | 1.178 | 1.116 | 1.169 | 8 × 10−1 | 9 × 10−1 | 7 × 10−1 | −0.02 | −0.09 | −0.12 |
| 40 | Mannitol | 1.775 | 1.444 | 1.492 | 1.172 | 1.353 | 1.606 | 10−1 | 7 × 10−1 | 7 × 10−1 | −0.60 | −0.09 | 0.11 |
| 41 | Arginine | 0.210 | 0.202 | 0.221 | 0.140 | 0.620 | 0.207 | 3 × 10−1 | 4 × 10−1 | 8 × 10−1 | −0.58 | 1.62 | −0.09 |
| Control (μg/mL) | Cocaine (μg/mL) | p-Value | Log2FC (Cocaine vs. Control) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | Compound | 3 h | 24 h | 48 h | 3 h | 24 h | 48 h | 3 h | 24 h | 48 h | 3 h | 24 h | 48 h |
| 1 | Benzoic acid | 0.190 | 0.215 | 0.254 | 7.662 | 14.26 | 18.63 | 10−3 | 2 × 10−7 | 3 × 10−4 | 5.33 | 6.05 | 6.19 |
| 2 | Choline | 0.124 | 0.072 | 0.032 | 0.189 | 0.198 | 0.165 | 3 × 10−3 | 10−8 | 4 × 10−4 | 0.61 | 1.46 | 2.37 |
| 3 | Aspartic acid | 2.039 | 1.965 | 2.770 | 2.379 | 5.397 | 6.420 | 4 × 10−1 | 5 × 10−4 | 2 × 10−3 | 0.22 | 1.46 | 1.21 |
| 4 | Malonic acid | 5.296 | 5.028 | 4.955 | 5.234 | 3.444 | 5.380 | 8 × 10−1 | 6 × 10−1 | 4 × 10−3 | −0.02 | −0.55 | 0.12 |
| 5 | Thymidine | 0.041 | 0.022 | 0.015 | 0.066 | 0.051 | 0.029 | 2 × 10−2 | 10−3 | 8 × 10−3 | 0.69 | 1.24 | 0.93 |
| 6 | Uridine | 0.061 | 0.060 | 0.086 | 0.141 | 0.236 | 0.326 | 10−5 | 5 × 10−3 | 2 × 10−2 | 1.21 | 1.97 | 1.93 |
| 7 | Phenylalanine | 26.10 | 19.32 | 25.03 | 38.31 | 49.78 | 49.09 | 10−1 | 4 × 10−5 | 2 × 10−2 | 0.55 | 1.37 | 0.97 |
| 8 | Hypoxanthine | 0.115 | 0.032 | 0.007 | 0.200 | 0.272 | 0.317 | 2 × 10−3 | 8 × 10−5 | 3 × 10−2 | 0.80 | 3.07 | 5.52 |
| 9 | Hippuric acid | 0.356 | 0.409 | 0.518 | 0.432 | 0.672 | 0.608 | 5 × 10−1 | 10−1 | 3 × 10−2 | 0.28 | 0.72 | 0.23 |
| 10 | Mannitol | 3.006 | 2.391 | 4.760 | 5.075 | 5.086 | 6.759 | 5 × 10−2 | 4 × 10−2 | 4 × 10−2 | 0.76 | 1.09 | 0.51 |
| 11 | Creatine | 1.606 | 1.766 | 2.209 | 2.472 | 3.364 | 3.447 | 3 × 10−2 | 5 × 10−4 | 4 × 10−2 | 0.62 | 0.93 | 0.64 |
| 12 | Glutamic acid | 29.35 | 35.92 | 60.79 | 37.60 | 85.88 | 134.8 | 3 × 10−1 | 3 × 10−4 | 5 × 10−2 | 0.36 | 1.26 | 1.15 |
| 13 | Glycine | 22.47 | 28.00 | 38.30 | 34.20 | 48.31 | 60.63 | 4 × 10−2 | 8 × 10−3 | 5 × 10−2 | 0.61 | 0.79 | 0.66 |
| 14 | Alanine | 10.21 | 19.83 | 35.71 | 13.47 | 38.74 | 48.84 | 10−1 | 10−3 | 8 × 10−2 | 0.40 | 0.97 | 0.45 |
| 15 | Asparagine | 1.623 | 2.150 | 3.106 | 1.985 | 3.849 | 5.670 | 6 × 10−2 | 10−3 | 8 × 10−2 | 0.29 | 0.84 | 0.87 |
| 16 | Arginine | 23.84 | 23.28 | 30.03 | 44.15 | 51.26 | 47.10 | 3 × 10−2 | 5 × 10−6 | 8 × 10−2 | 0.89 | 1.14 | 0.65 |
| 17 | Xanthine | 0.342 | 0.371 | 0.631 | 0.592 | 0.893 | 1.167 | 5 × 10−3 | 3 × 10−3 | 9 × 10−2 | 0.79 | 1.27 | 0.89 |
| 18 | Acetylcarnitine | 0.049 | 0.048 | 0.065 | 0.067 | 0.088 | 0.092 | 9 × 10−2 | 3 × 10−4 | 10−1 | 0.45 | 0.86 | 0.51 |
| 19 | Homocysteine | 14.37 | 21.51 | 31.46 | 14.29 | 42.48 | 50.90 | 100 | 8 × 10−2 | 10−1 | −0.01 | 0.98 | 0.69 |
| 20 | Norvaline-Valine | 40.54 | 35.39 | 47.70 | 55.02 | 66.90 | 67.49 | 10−1 | 8 × 10−5 | 10−1 | 0.44 | 0.92 | 0.50 |
| 21 | Methionine | 13.19 | 10.56 | 14.68 | 18.00 | 21.98 | 21.27 | 7 × 10−2 | 3 × 10−4 | 10−1 | 0.45 | 1.06 | 0.53 |
| 22 | Tryptophan | 6.289 | 4.624 | 7.233 | 8.632 | 10.94 | 11.06 | 2 × 10−1 | 8 × 10−5 | 10−1 | 0.46 | 1.24 | 0.61 |
| 23 | Taurine | 1.001 | 1.032 | 1.016 | 1.453 | 1.484 | 1.309 | 10−1 | 10−1 | 10−1 | 0.54 | 0.52 | 0.37 |
| 24 | Lysine | 59.84 | 53.75 | 76.39 | 89.80 | 110.5 | 111.4 | 6 × 10−2 | 6 × 10−5 | 10−1 | 0.59 | 1.04 | 0.54 |
| 25 | Uracil | 0.331 | 0.374 | 0.570 | 0.636 | 0.724 | 0.837 | 4 × 10−2 | 6 × 10−4 | 10−1 | 0.95 | 0.95 | 0.55 |
| 26 | Isoleucine | 57.14 | 46.12 | 62.89 | 84.93 | 102.6 | 94.04 | 7 × 10−2 | 2 × 10−5 | 10−1 | 0.57 | 1.15 | 0.58 |
| 27 | Thymine | 0.030 | 0.062 | 0.095 | 0.052 | 0.092 | 0.133 | 2 × 10−1 | 4 × 10−3 | 10−1 | 0.79 | 0.57 | 0.49 |
| 28 | Adenine | 0.018 | 0.017 | 0.017 | 0.018 | 0.019 | 0.018 | 6 × 10−1 | 7 × 10−3 | 10−1 | 0.07 | 0.16 | 0.11 |
| 29 | leucine | 40.98 | 33.85 | 46.79 | 64.32 | 77.14 | 68.77 | 7 × 10−2 | 5 × 10−5 | 2 × 10−1 | 0.65 | 1.19 | 0.56 |
| 30 | Lactic acid | 476.4 | 612.1 | 879.3 | 552.9 | 1067 | 1230 | 4 × 10−1 | 3 × 10−3 | 2 × 10−1 | 0.21 | 0.80 | 0.48 |
| 31 | Pantothenate | 1.650 | 1.376 | 2.288 | 2.444 | 3.062 | 3.134 | 7 × 10−2 | 2 × 10−4 | 2 × 10−1 | 0.57 | 1.15 | 0.45 |
| 32 | Anthranilic acid | 6.135 | 8.028 | 20.20 | 8.463 | 18.48 | 31.89 | 4 × 10−2 | 5 × 10−3 | 2 × 10−1 | 0.46 | 1.20 | 0.66 |
| 33 | Tyrosine | 32.09 | 31.30 | 52.33 | 55.24 | 61.81 | 78.54 | 2 × 10−2 | 2 × 10−3 | 2 × 10−1 | 0.78 | 0.98 | 0.59 |
| 34 | Creatinine | 1.413 | 1.313 | 1.961 | 1.994 | 2.483 | 2.569 | 6 × 10−2 | 6 × 10−5 | 2 × 10−1 | 0.50 | 0.92 | 0.39 |
| 35 | Allontoin | 1.506 | 1.426 | 2.182 | 2.306 | 2.677 | 2.802 | 5 × 10−2 | 3 × 10−4 | 2 × 10−1 | 0.62 | 0.91 | 0.36 |
| 36 | Threonine | 29.51 | 28.00 | 40.98 | 42.43 | 51.64 | 52.87 | 10−1 | 2 × 10−4 | 2 × 10−1 | 0.52 | 0.88 | 0.37 |
| 37 | Betaine | 1.229 | 1.157 | 1.640 | 1.641 | 1.988 | 2.069 | 6 × 10−2 | 8 × 10−4 | 2 × 10−1 | 0.42 | 0.78 | 0.33 |
| 38 | Folic acid | 2.834 | 3.069 | 3.110 | 3.206 | 4.177 | 4.247 | 6 × 10−1 | 3 × 10−2 | 2 × 10−1 | 0.18 | 0.44 | 0.45 |
| 39 | Sorbitol | 5.067 | 6.675 | 9.881 | 5.629 | 6.356 | 13.84 | 9 × 10−1 | 9 × 10−1 | 3 × 10−1 | 0.15 | −0.07 | 0.49 |
| 40 | Pyroglutamic | 92.66 | 105.46 | 161.7 | 139.1 | 175.1 | 201.1 | 6 × 10−2 | 6 × 10−4 | 3 × 10−1 | 0.59 | 0.73 | 0.31 |
| 41 | Ornithine | 4.183 | 6.162 | 11.27 | 7.507 | 10.65 | 14.07 | 4 × 10−3 | 4 × 10−3 | 3 × 10−1 | 0.84 | 0.79 | 0.32 |
| 42 | Pyruvic acid | 4.507 | 4.636 | 6.377 | 5.944 | 7.868 | 8.150 | 10−1 | 10−3 | 3 × 10−1 | 0.40 | 0.76 | 0.35 |
| 43 | Nicotinamide | 2.225 | 2.097 | 2.910 | 2.624 | 3.322 | 3.548 | 10−1 | 5 × 10−4 | 3 × 10−1 | 0.24 | 0.66 | 0.29 |
| 44 | Fructose | 3.865 | 1.175 | n.d. * | 6.063 | 3.015 | 0.100 | 4 × 10−2 | 3 × 10−3 | 4 × 10−1 | 0.65 | 1.36 | − |
| 45 | Serine | 17.06 | 14.44 | 18.50 | 20.58 | 20.60 | 16.16 | 10−1 | 4 × 10−2 | 5 × 10−1 | 0.27 | 0.51 | −0.20 |
| 46 | Glutamine | 236.3 | 187.8 | 222.6 | 332.1 | 322.4 | 252.6 | 6 × 10−2 | 10−4 | 5 × 10−1 | 0.49 | 0.78 | 0.18 |
| 47 | Pyridoxine | 0.007 | 0.013 | 0.041 | 0.009 | 0.027 | 0.037 | 3 × 10−1 | 2 × 10−3 | 7 × 10−1 | 0.47 | 0.98 | −0.14 |
| 48 | Acetyl aspartate | 0.166 | 0.193 | 0.406 | 0.161 | 0.293 | 0.340 | 8 × 10−1 | 3 × 10−1 | 8 × 10−1 | −0.04 | 0.60 | −0.26 |
| 49 | Inositol | 13.43 | 12.06 | 13.79 | 12.10 | 12.86 | 14.71 | 8 × 10−1 | 9 × 10−1 | 8 × 10−1 | −0.15 | 0.09 | 0.09 |
| 50 | Trimethylamine-n-oxide | 0.050 | 0.041 | 0.056 | 0.058 | 0.065 | 0.058 | 5 × 10−1 | 9 × 10−4 | 8 × 10−1 | 0.20 | 0.65 | 0.05 |
| 51 | Cytosine | 0.049 | 0.048 | 0.050 | 0.049 | 0.049 | 0.051 | 9 × 10−1 | 4 × 10−1 | 8 × 10−1 | 0.01 | 0.05 | 0.02 |
| 52 | Proline | 2.390 | 4.543 | 10.499 | 2.998 | 6.574 | 10.14 | 2 × 10−1 | 5 × 10−3 | 8 × 10−1 | 0.33 | 0.53 | −0.05 |
| 53 | Thiamine | 0.103 | 0.206 | 0.241 | 0.104 | 0.208 | 0.247 | 8 × 10−1 | 8 × 10−1 | 9 × 10−1 | 0.01 | 0.02 | 0.03 |
| 54 | Glucose | 261.21 | 101.07 | 8.194 | 352.0 | 154.1 | 7.714 | 2 × 10−2 | 3 × 10−2 | 100 | 0.43 | 0.61 | −0.09 |
| 55 | Mannose | 48.812 | 20.502 | n.d. * | 75.442 | 35.656 | n.d. * | 9 × 10−3 | 5 × 10−3 | - | 0.63 | 0.80 | − |
| Comparisons | Analysis | R2X | R2Y | Q2Y |
|---|---|---|---|---|
| Control vs. cocaine-treated vs. QC (intracellular material) | PCA | 0.604 | 0.388 | |
| Control vs. cocaine-treated (intracellular material) | PCA | 0.644 | 0.417 | |
| Control vs. cocaine-treated (intracellular material) | OPLS-DA | 0.638 | 0.859 | 0.685 |
| Control vs. cocaine-treated vs. QC (cell medium) | PCA | 0.800 | 0.728 | |
| Control vs. cocaine-treated (cell medium) | PCA | 0.806 | 0.726 | |
| Control vs. cocaine-treated (cell medium) | OPLS-DA | 0.800 | 0.835 | 0.765 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krokos, A.; Deda, O.; Virgiliou, C.; Gika, H.; Raikos, N.; Aggelidou, E.; Kritis, A.; Theodoridis, G. Evaluation of Cocaine Effect on Endogenous Metabolites of HepG2 Cells Using Targeted Metabolomics. Molecules 2021, 26, 4610. https://doi.org/10.3390/molecules26154610
Krokos A, Deda O, Virgiliou C, Gika H, Raikos N, Aggelidou E, Kritis A, Theodoridis G. Evaluation of Cocaine Effect on Endogenous Metabolites of HepG2 Cells Using Targeted Metabolomics. Molecules. 2021; 26(15):4610. https://doi.org/10.3390/molecules26154610
Chicago/Turabian StyleKrokos, Adamantios, Olga Deda, Christina Virgiliou, Helen Gika, Nikolaos Raikos, Eleni Aggelidou, Aristeidis Kritis, and Georgios Theodoridis. 2021. "Evaluation of Cocaine Effect on Endogenous Metabolites of HepG2 Cells Using Targeted Metabolomics" Molecules 26, no. 15: 4610. https://doi.org/10.3390/molecules26154610
APA StyleKrokos, A., Deda, O., Virgiliou, C., Gika, H., Raikos, N., Aggelidou, E., Kritis, A., & Theodoridis, G. (2021). Evaluation of Cocaine Effect on Endogenous Metabolites of HepG2 Cells Using Targeted Metabolomics. Molecules, 26(15), 4610. https://doi.org/10.3390/molecules26154610