
Lipoic Acid-Modified Oligoethyleneimine-Mediated miR-34a Delivery to Achieve the Anti-Tumor Efficacy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Synthesis and Characterization of LA-OEI
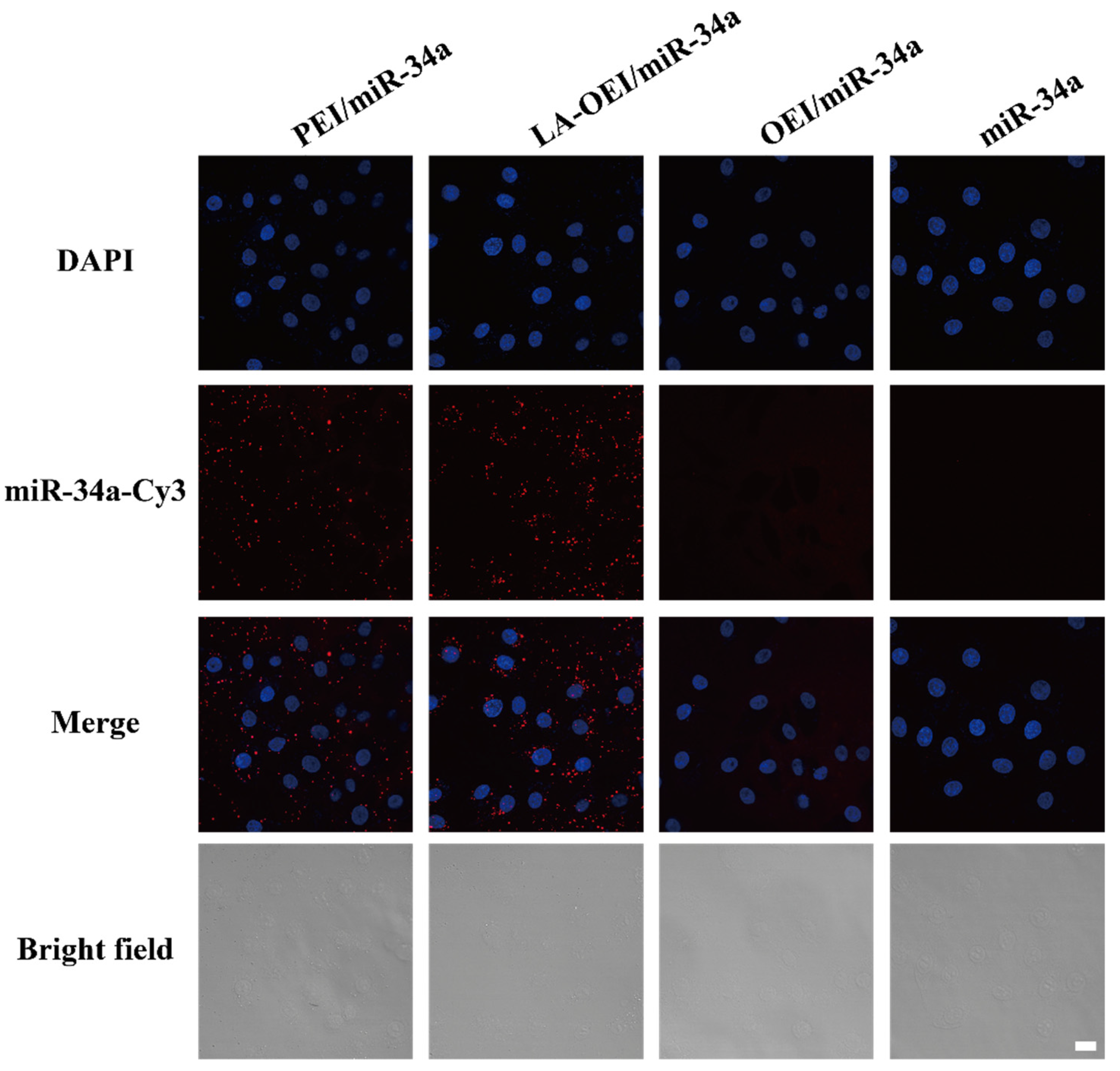
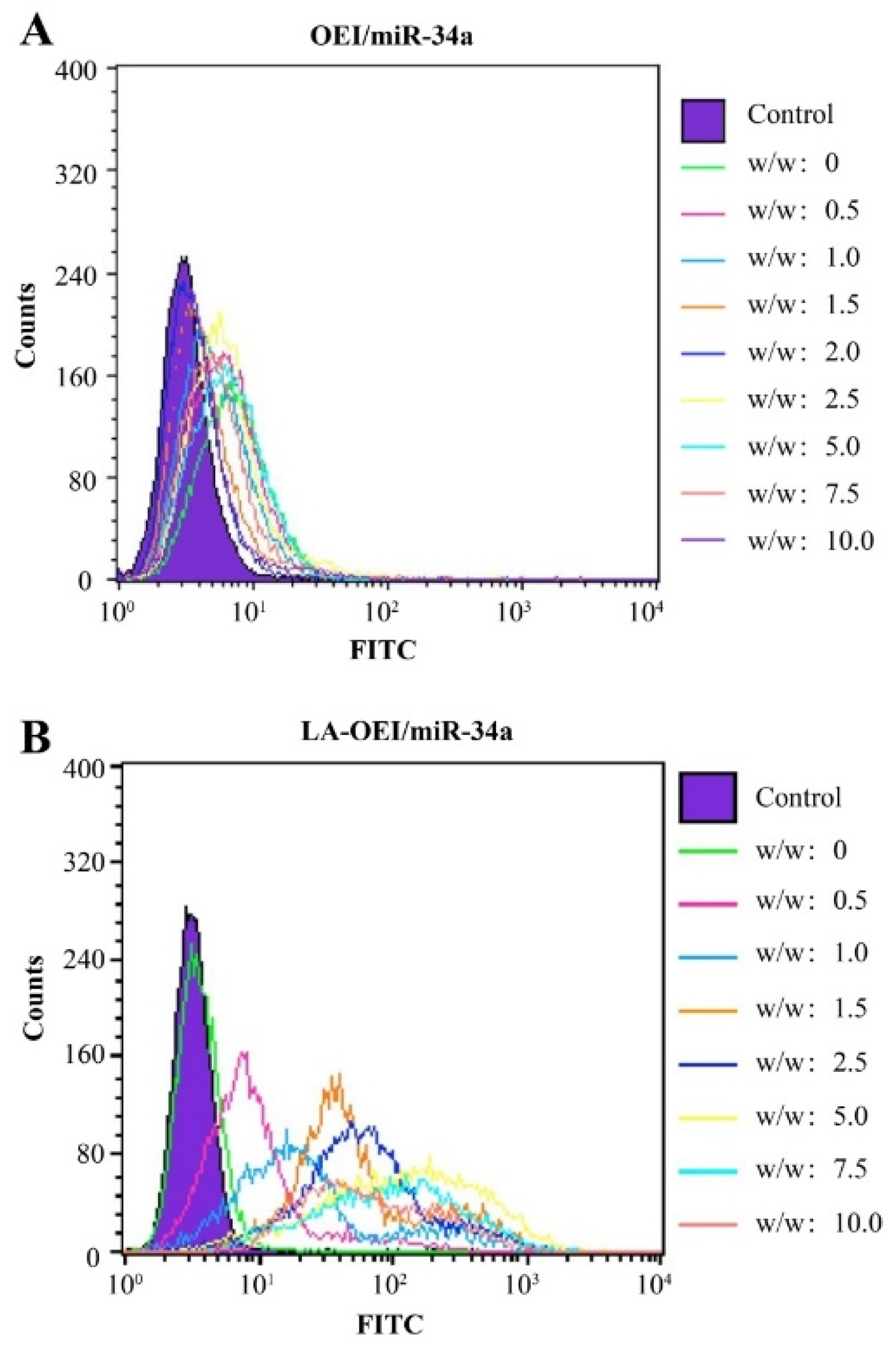
2.2. In Vitro Transfection Efficiency of LA-OEI/miR-34a Nanoparticles
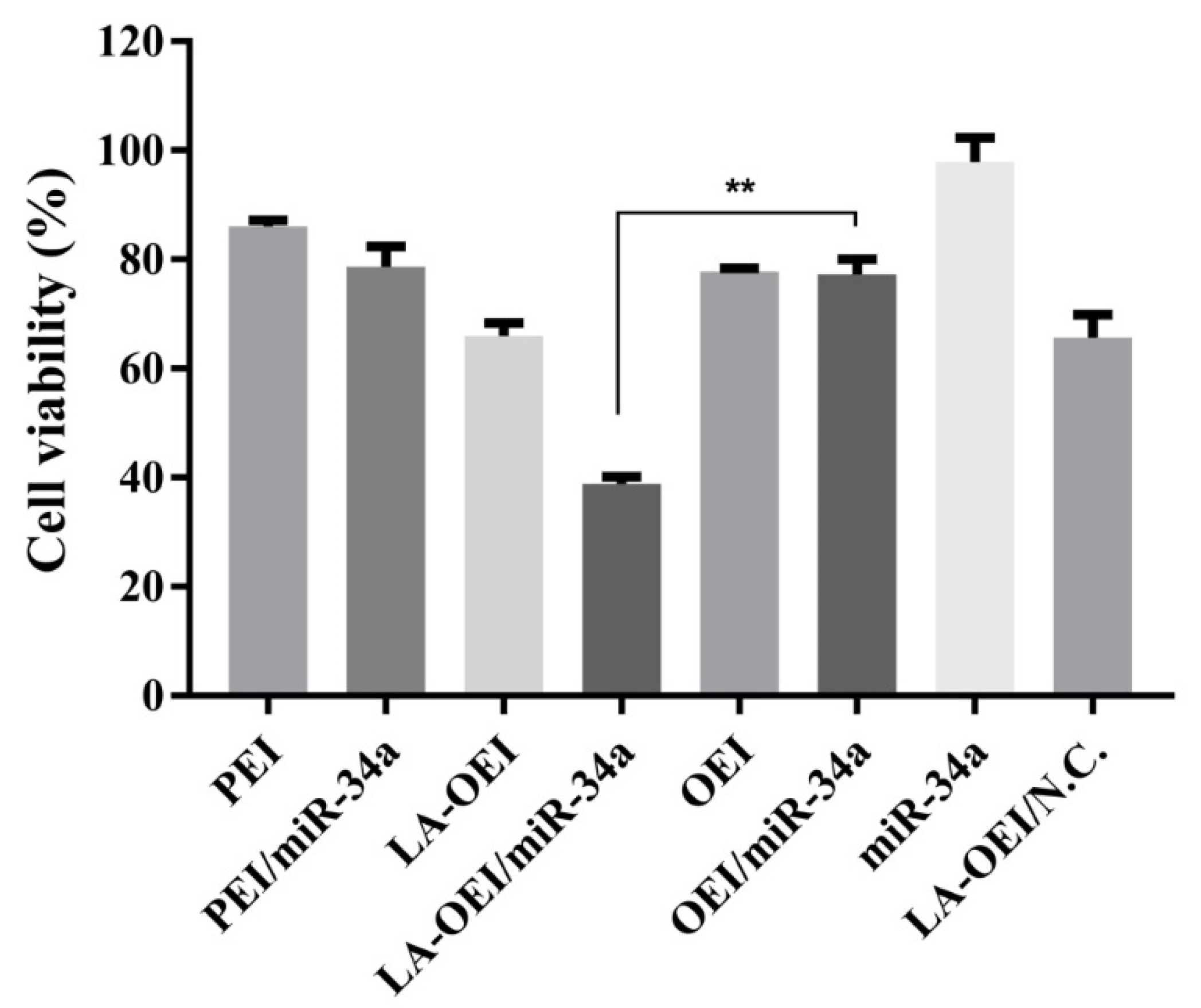
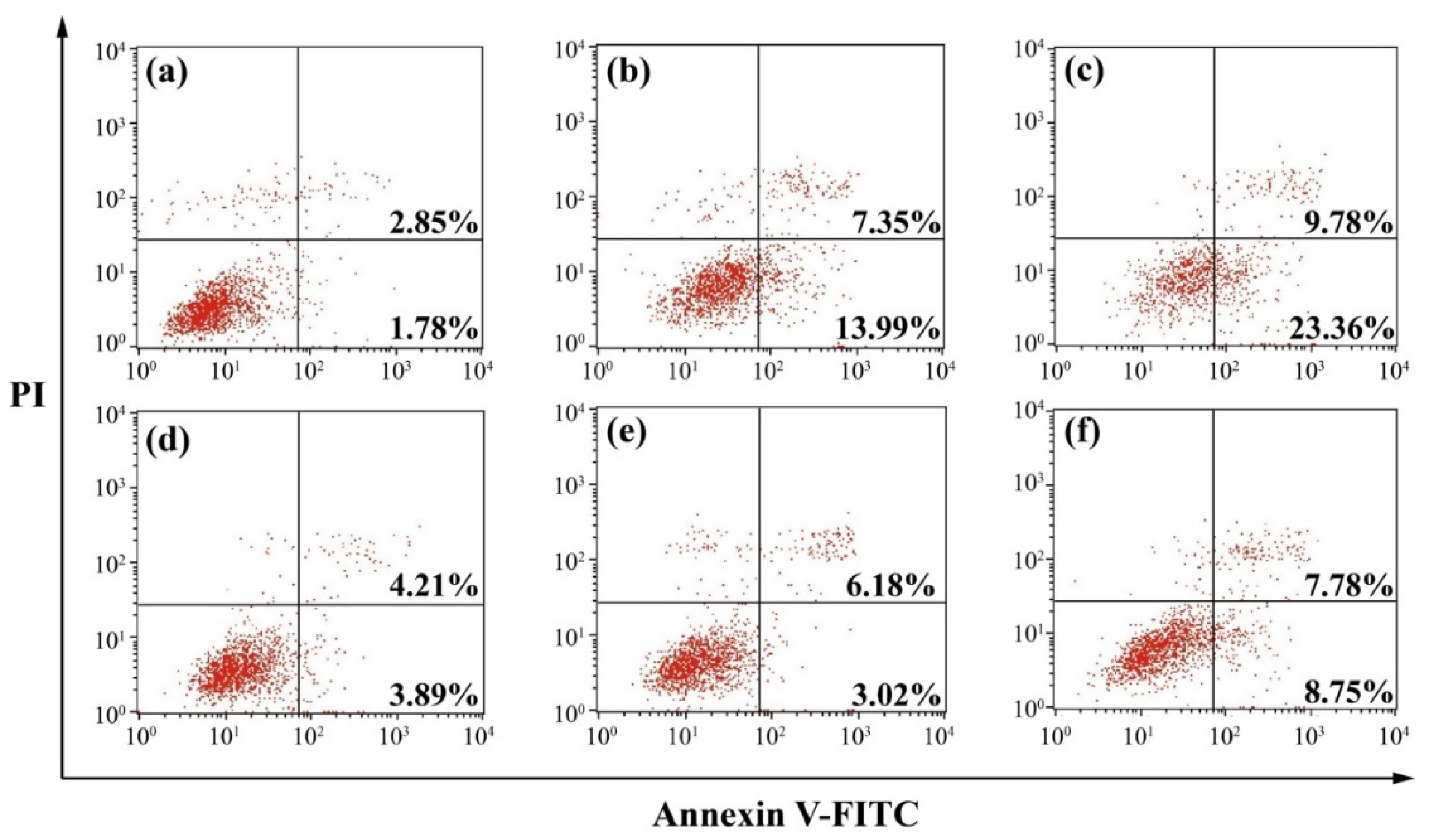
2.3. Anti-Proliferative Effect of LA-OEI/miR-34a Nanoparticles
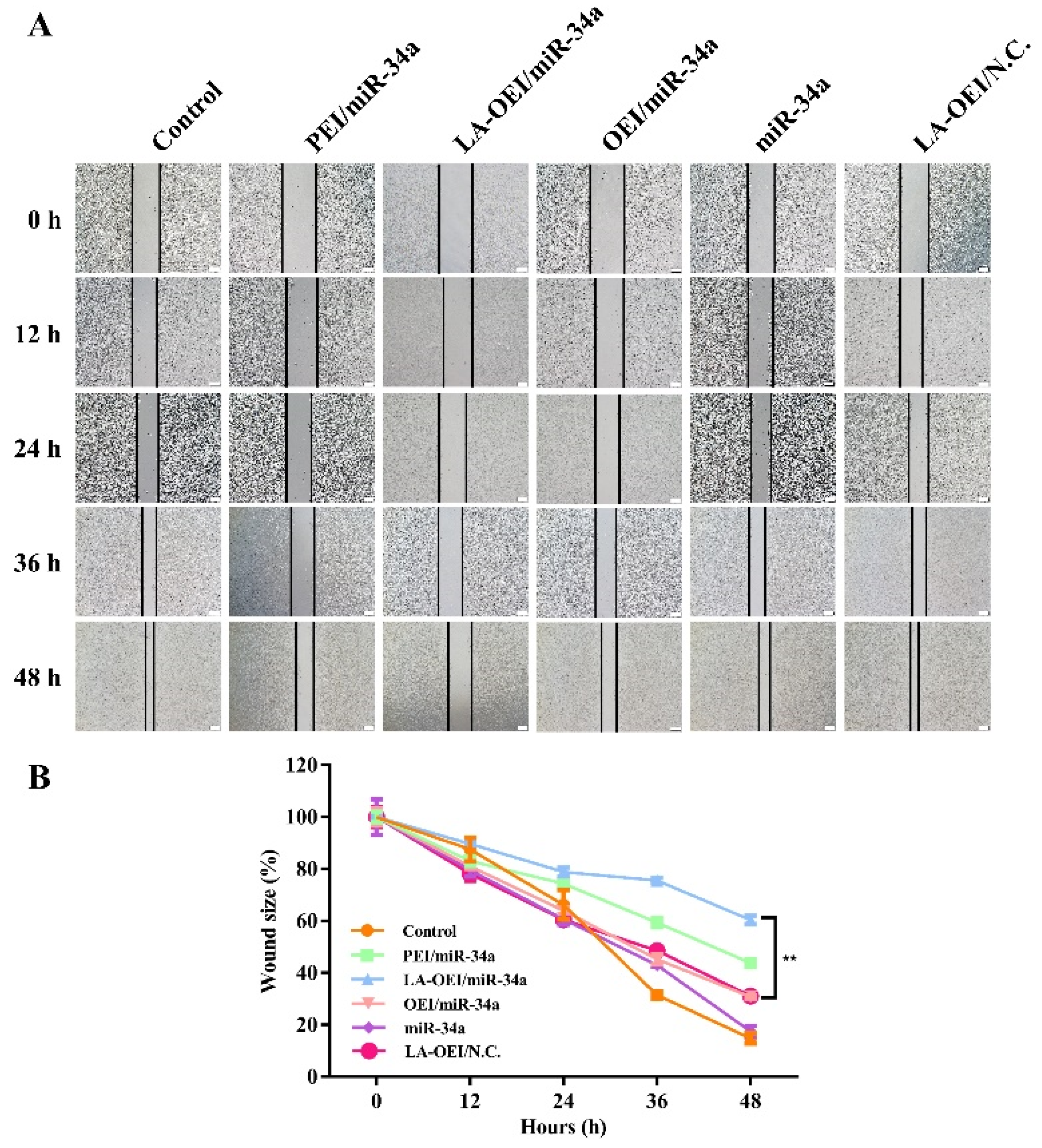
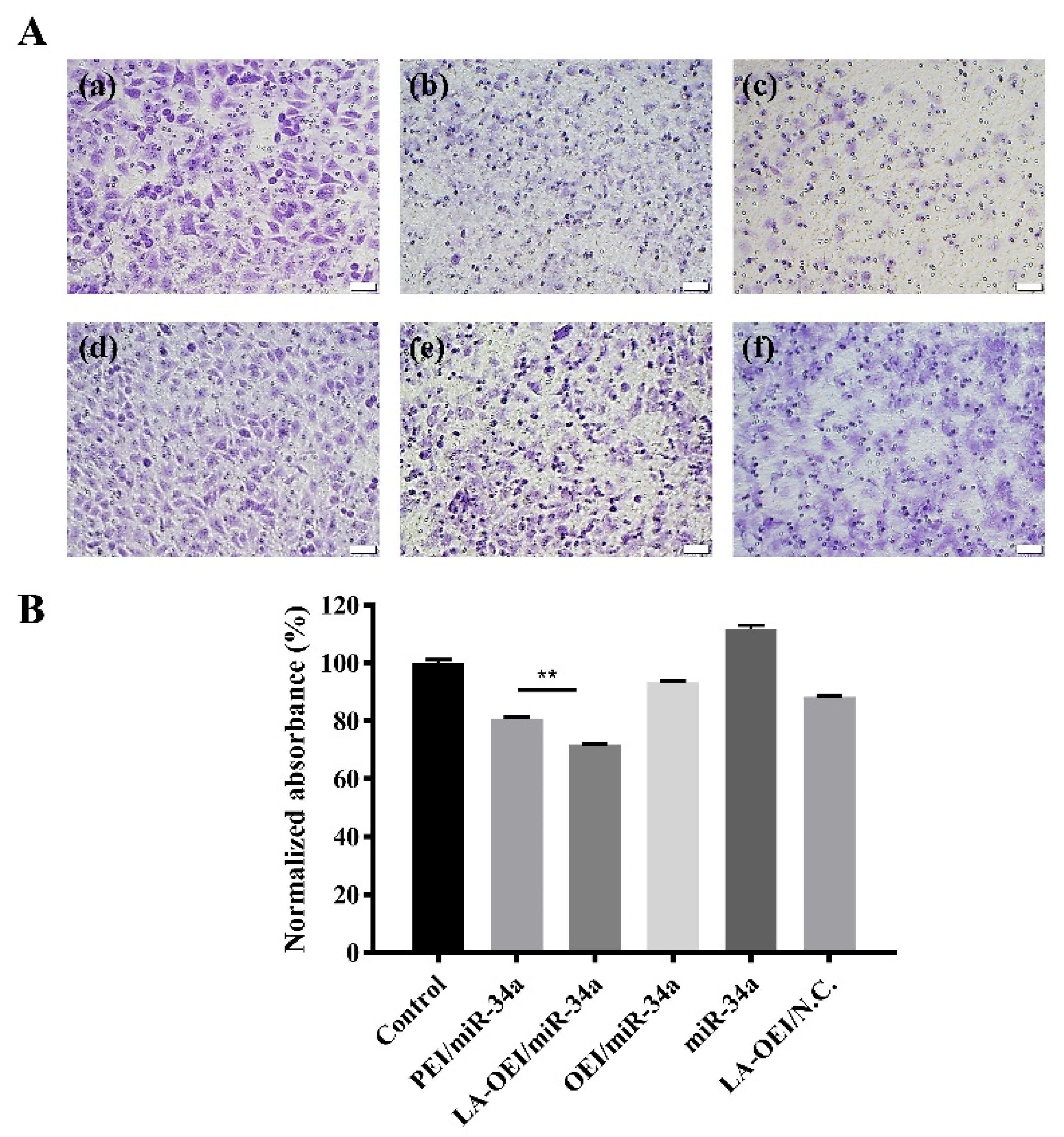
2.4. Anti-Migration Effect of LA-OEI/miR-34a Nanoparticles
3. Materials and Methods
3.1. Materials
3.2. Preparation and Characterization of LA-OEI
3.3. Gel Retardation Assay
3.4. In Vitro miR-34a Transfection
3.5. Anti-Proliferative Effect of miR-34a Transfection
3.6. Cell Apoptosis Analysis
3.7. Live/Dead Cell Staining
3.8. Cell Colony Formation Assay
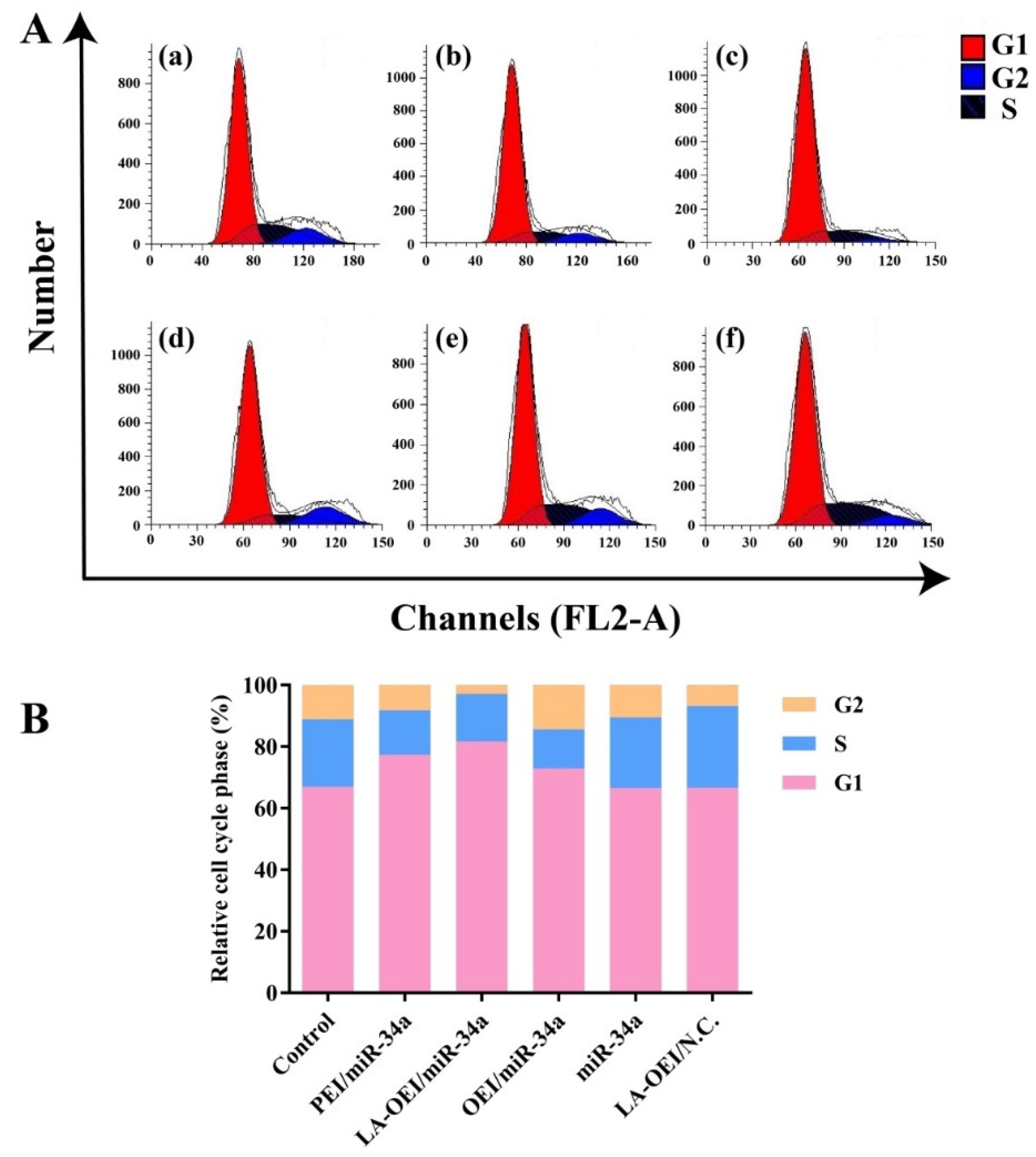
3.9. Cell Cycle Arrest Analysis
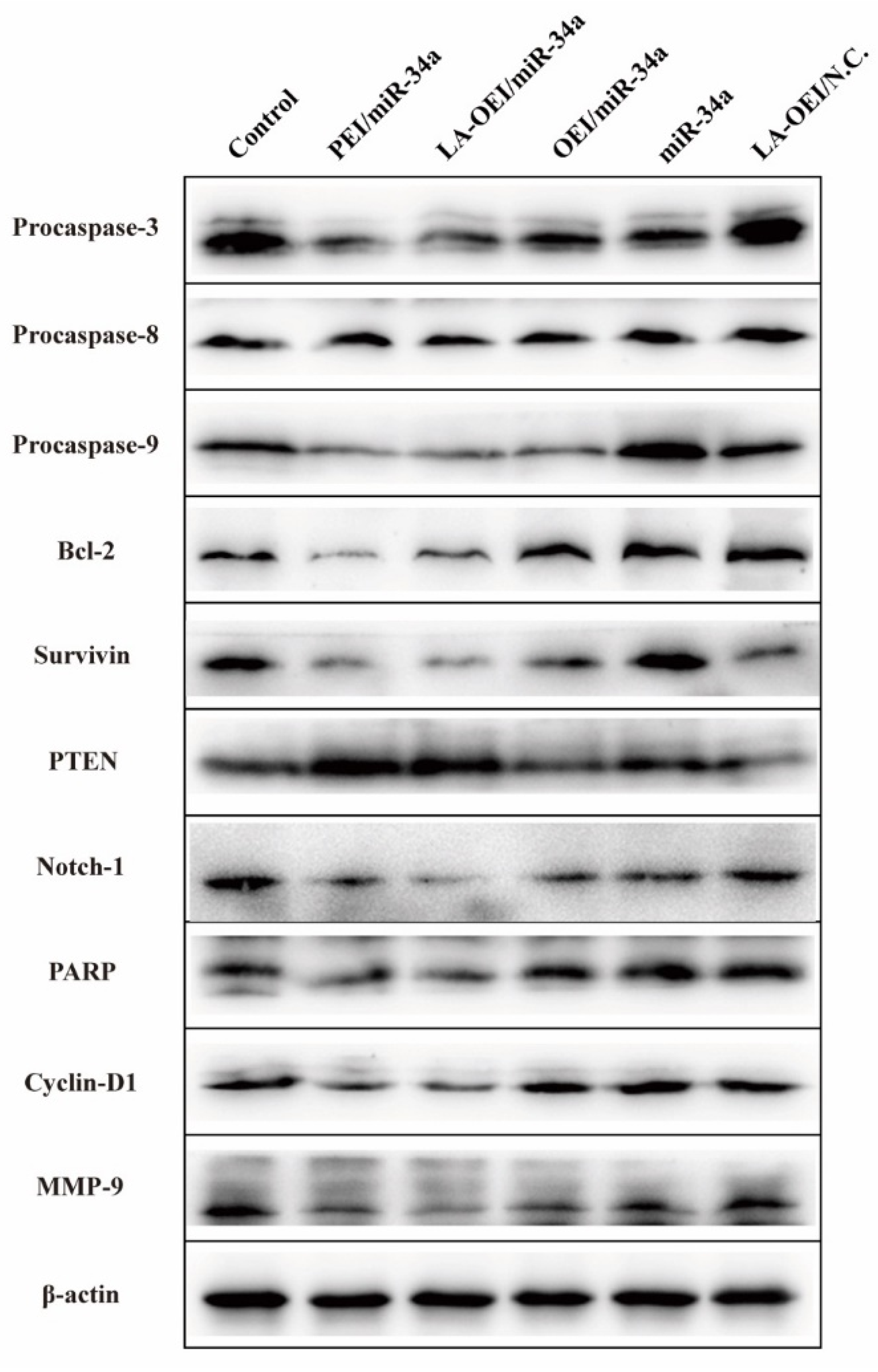
3.10. Western Blotting Assay
3.11. Caspase-3, -8 and -9 Activities Analysis
3.12. Anti-Migration Effect of miR-34a Transfection
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.M.; Wilson, W.R. Exploiting tumour hypoxia in cancer treatment. Nat. Rev. Cancer 2004, 4, 437–447. [Google Scholar] [CrossRef]
- White, B.D.; Duan, C.; Townley, H.E. Nanoparticle activation methods in cancer treatment. Biomolecules 2019, 9, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naldini, L. Gene therapy returns to centre stage. Nature 2015, 526, 351–360. [Google Scholar] [CrossRef]
- Geng, J.; Guo, X.; Wang, L.; Nguyen, R.Q.; Wang, F.; Liu, C.; Wang, H. Intracellular delivery of DNA and protein by a novel cell-permeable peptide derived from DOT1L. Biomolecules 2020, 10, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Shen, X.; Zou, Q.; Wang, S.; Tang, S.; Zhang, G. Biological functions of microRNAs: A review. J. Physiol. Biochem. 2011, 67, 129–139. [Google Scholar] [CrossRef]
- Mizuguchi, Y.; Takizawa, T.; Yoshida, H.; Uchida, E. Dysregulated miRNA in progression of hepatocellular carcinoma: A systematic review. Hepatol. Res. 2016, 46, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, B.; Ramlogan-Steel, C.A.; Steel, J.C. MicroRNA-regulated gene delivery systems for research and therapeutic purposes. Molecules 2018, 23, 1500. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Kelnar, K.; Liu, B.; Chen, X.; Calhoun-Davis, T.; Li, H.; Patrawala, L.; Yan, H.; Jeter, C.; Honorio, S.; et al. The microRNA miR-34a inhibits prostate cancer stem cells and metastasis by directly repressing CD44. Nat. Med. 2011, 17, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Raver-Shapira, N.; Marciano, E.; Meiri, E.; Spector, Y.; Rosenfeld, N.; Moskovits, N.; Bentwich, Z.; Oren, M. Transcriptional activation of miR-34a contributes to p53-mediated apoptosis. Mol. Cell 2007, 26, 731–743. [Google Scholar] [CrossRef]
- Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J. MiR-34a repression of SIRT1 regulates apoptosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13421–13426. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Cao, M.; Zhang, J.; Hu, K.; Yin, Z.; Zhou, Z.; Xiao, X.; Yang, Y.; Sheng, W.; Wu, Y.; et al. Hyaluronic acid-chitosan nanoparticles for co-delivery of MiR-34a and doxorubicin in therapy against triple negative breast cancer. Biomaterials 2014, 35, 4333–4344. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Liang, X.; Wang, Y.; Han, H.; Yang, J.; Fang, X.; Li, Q. Phenylboronic acid-functionalized polyamidoamine-mediated miR-34a delivery for the treatment of gastric cancer. Biomater. Sci. 2019, 7, 1632–1642. [Google Scholar] [CrossRef]
- Young, L.S.; Searle, P.F.; Onion, D.; Mautner, V. Viral gene therapy strategies: From basic science to clinical application. J. Pathol. 2006, 208, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Kay, M.A.; Glorioso, J.C.; Naldini, L. Viral vectors for gene therapy: The art of turning infectious agents into vehicles of therapeutics. Nat. Med. 2001, 7, 33–40. [Google Scholar] [CrossRef]
- Thomas, T.J.; Tajmir-Riahi, H.A.; Pillai, C.K.S. Biodegradable polymers for gene delivery. Molecules 2019, 24, 3744. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.; Klibanov, A. Non-viral gene therapy: Polycation-mediated DNA delivery. Appl. Microbiol. Biotechnol. 2003, 62, 27–34. [Google Scholar] [CrossRef]
- Xing, Z.; Gao, S.; Duan, Y.; Han, H.; Li, L.; Yang, Y.; Li, Q. Delivery of DNAzyme targeting aurora kinase A to inhibit the proliferation and migration of human prostate cancer. Int. J. Nanomed. 2015, 10, 5715–5727. [Google Scholar]
- Zhang, J.; Wu, D.; Xing, Z.; Liang, S.; Han, H.; Shi, H.; Zhang, Y.; Yang, Y.; Li, Q. N-Isopropylacrylamide-modified polyethylenimine-mediated p53 gene delivery to prevent the proliferation of cancer cells. Colloids Surf. B Biointerfaces 2015, 129, 54–62. [Google Scholar] [CrossRef]
- Mainini, F.; Eccles, M.R. Lipid and polymer-based nanoparticle siRNA delivery systems for cancer therapy. Molecules 2020, 25, 2692. [Google Scholar] [CrossRef] [PubMed]
- Wahane, A.; Waghmode, A.; Kapphahn, A.; Dhuri, K.; Gupta, A.; Bahai, R. Role of lipid-based and polymer-based non-viral vectors in nucleic acid delivery for next-generation gene therapy. Molecules 2020, 25, 2866. [Google Scholar] [CrossRef]
- Meng, Y.; Wang, S.; Li, C.; Qian, M.; Yan, X.; Yao, S.; Peng, X.; Wang, Y.; Huang, R. Photothermal combined gene therapy achieved by polyethyleneimine-grafted oxidized mesoporous carbon nanospheres. Biomaterials 2016, 100, 134–142. [Google Scholar] [CrossRef]
- Guan, X.; Guo, Z.; Wang, T.; Lin, L.; Chen, J.; Tian, H.; Chen, X. A pH-responsive detachable PEG shielding strategy for gene delivery system in cancer therapy. Biomacromolecules 2017, 18, 1342–1349. [Google Scholar] [CrossRef]
- Dong, X.; Tian, H.; Chen, L.; Chen, J.; Chen, X. Biodegradable mPEG-b-P(MCC-g-OEI) copolymers for efficient gene delivery. J. Control. Release 2011, 152, 135–142. [Google Scholar] [CrossRef]
- Gosselin, M.A.; Guo, W.; Lee, R.J. Efficient gene transfer using reversibly cross-linked low molecular weight polyethylenimine. Bioconjug. Chem. 2001, 12, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, C.; Li, H.; Wang, X.; Goh, S.H.; Ding, J.L.; Wang, D.Y.; Leong, K.W. Cationic supramolecules composed of multiple oligoethylenimine-grafted β-cyclodextrins threaded on a polymer chain for efficient gene delivery. Adv. Mater. 2006, 18, 2969–2974. [Google Scholar] [CrossRef]
- Wu, X.R.; Zhang, J.; Zhang, J.H.; Xiao, Y.P.; He, X.; Liu, Y.H.; Yu, X.Q. Amino acid-linked low molecular weight polyethylenimine for improved gene delivery and biocompatibility. Molecules 2020, 25, 975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knorr, V.; Russ, V.; Allmendinger, L.; Ogris, M.; Wagner, E. Acetal linked oligoethylenimines for use as pH-sensitive gene carriers. Bioconjug. Chem. 2008, 19, 1625–1634. [Google Scholar] [CrossRef]
- Philipp, A.; Zhao, X.; Tarcha, P.; Wagner, E.; Zintchenko, A. Hydrophobically modified oligoethylenimines as highly efficient transfection agents for siRNA delivery. Bioconj. Chem. 2009, 20, 2055–2061. [Google Scholar] [CrossRef]
- Xia, J.; Tian, H.; Chen, L.; Lin, L.; Guo, Z.; Chen, J.; Chen, X. Oligoethylenimines grafted to PEGylated poly(β-amino ester)s for gene delivery. Biomacromolecules 2011, 12, 1024–1031. [Google Scholar] [CrossRef]
- Russ, V.; Gunther, M.; Halama, A.; Ogris, M.; Wagner, E. Oligoethylenimine-grafted polypropylenimine dendrimers as degradable and biocompatible synthetic vectors for gene delivery. J. Control. Release 2008, 132, 131–140. [Google Scholar] [CrossRef]
- Tang, X.; Li, Q.; Liang, X.; Liu, Z.; Li, Q. Inhibition of proliferation and migration of tumor cells through lipoic acid-modified oligoethylenimine-mediated p53 gene delivery. New J. Chem. 2019, 43, 2758–2765. [Google Scholar] [CrossRef]
- Tang, X.; Liang, X.; Wen, K.; Chen, Y.; Han, H.; Li, Q. Dual ATP/reduction-responsive polyplex to achieve the co-delivery of doxorubicin and miR-23b for the cancer treatment. Colloids Surf. B Biointerfaces 2021, 206, 111955. [Google Scholar] [CrossRef]
- Moura, F.A.; de Andrade, K.Q.; dos Santos, J.C.; Goulart, M.O. Lipoic acid: Its antioxidant and anti-inflammatory role and clinical applications. Curr. Top. Med. Chem. 2015, 15, 458–483. [Google Scholar] [CrossRef]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.J.; Lopez-Chavez, A.; Citrin, D.; Janik, J.E.; Morris, J.C. Impacting tumor cell-fate by targeting the inhibitor of apoptosis protein survivin. Mol. Cancer 2011, 10, 35. [Google Scholar] [CrossRef] [Green Version]
- Malaney, P.; Uversky, V.N.; Dave, V. PTEN proteoforms in biology and disease. Cell. Mol. Life Sci. 2017, 74, 2783–2794. [Google Scholar] [CrossRef]
- Qin, X.; Wang, X.; Wang, Y.; Tang, Z.; Cui, Q.; Xi, J.; Li, Y.S.; Chien, S.; Wang, N. MicroRNA-19a mediates the suppressive effect of laminar flow on cyclin D1 expression in human umbilical vein endothelial cells. Proc. Natl. Acad. Sci. USA 2010, 107, 3240–3244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, B.; Koul, S.; Petersen, J.; Khandrika, L.; Hwa, J.S.; Meacham, R.B.; Wilson, S.; Koul, H.K. p38 mitogen-activated protein kinase-driven MAPKAPK2 regulates invasion of bladder cancer by modulation of MMP-2 and MMP-9 activity. Cancer Res. 2010, 70, 832–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, S.L.; Kunnimalaiyaan, M.; Drenzek, J.; Seiler, N. Notch 1 signaling is active in ovarian cancer. Gynecol. Oncol. 2010, 117, 130–133. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Wang, L.; Chen, Y.; Han, H.; Li, Q. Lipoic Acid-Modified Oligoethyleneimine-Mediated miR-34a Delivery to Achieve the Anti-Tumor Efficacy. Molecules 2021, 26, 4827. https://doi.org/10.3390/molecules26164827
Huang Y, Wang L, Chen Y, Han H, Li Q. Lipoic Acid-Modified Oligoethyleneimine-Mediated miR-34a Delivery to Achieve the Anti-Tumor Efficacy. Molecules. 2021; 26(16):4827. https://doi.org/10.3390/molecules26164827
Chicago/Turabian StyleHuang, Yu, Longxiang Wang, Yingxuan Chen, Haobo Han, and Quanshun Li. 2021. "Lipoic Acid-Modified Oligoethyleneimine-Mediated miR-34a Delivery to Achieve the Anti-Tumor Efficacy" Molecules 26, no. 16: 4827. https://doi.org/10.3390/molecules26164827
APA StyleHuang, Y., Wang, L., Chen, Y., Han, H., & Li, Q. (2021). Lipoic Acid-Modified Oligoethyleneimine-Mediated miR-34a Delivery to Achieve the Anti-Tumor Efficacy. Molecules, 26(16), 4827. https://doi.org/10.3390/molecules26164827