
Potential Effects of Soy Isoflavones on the Prevention of Metabolic Syndrome



Abstract
:
1. Introduction
2. Soybean Components and Isoflavone Characteristics
2.1. Soybean Components
2.2. Dietary Sources of Isoflavones
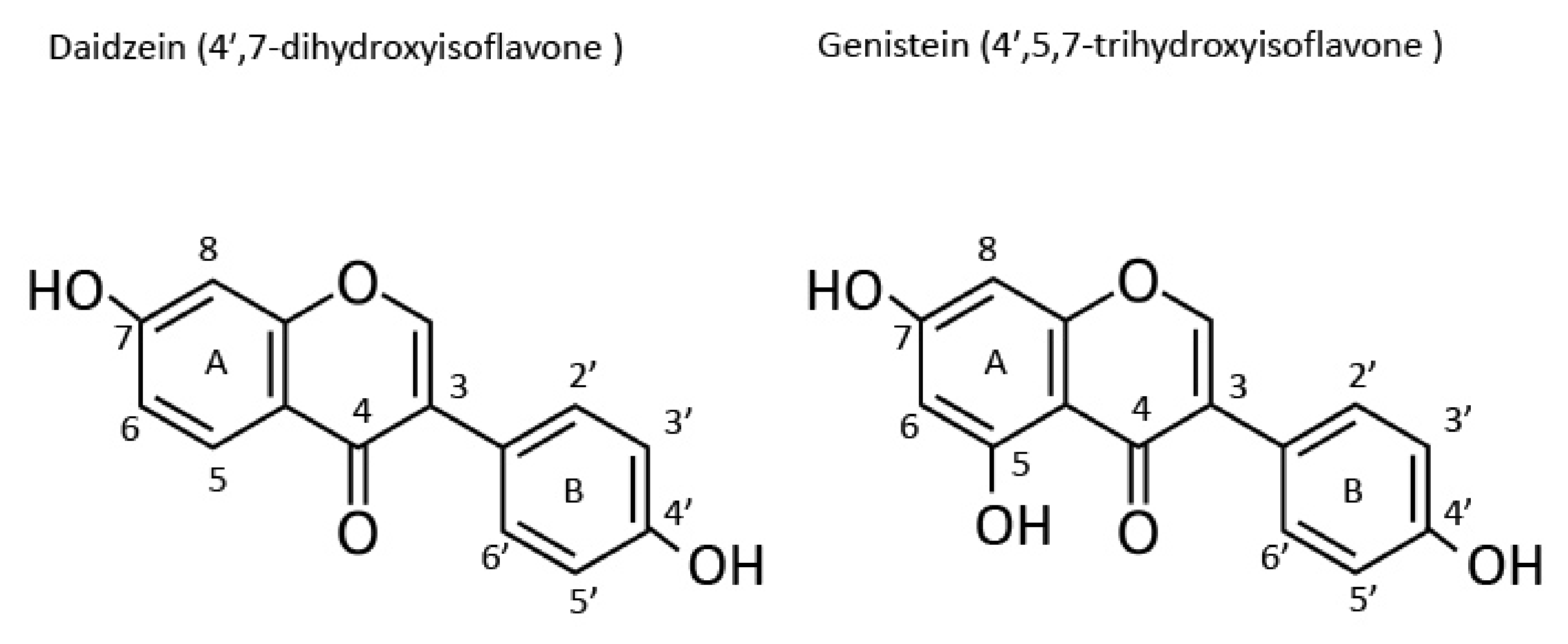
2.3. Isoflavone Types and Structures
2.4. Absorption of Isoflavones in the Human Body
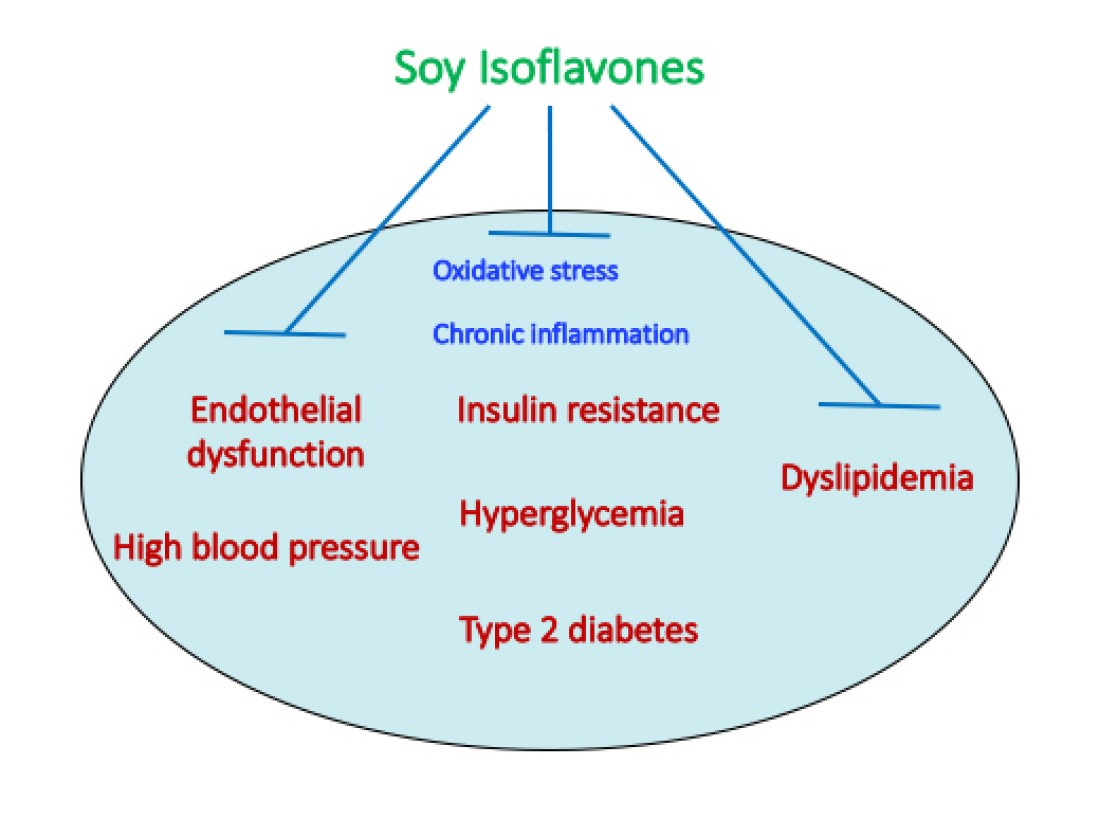
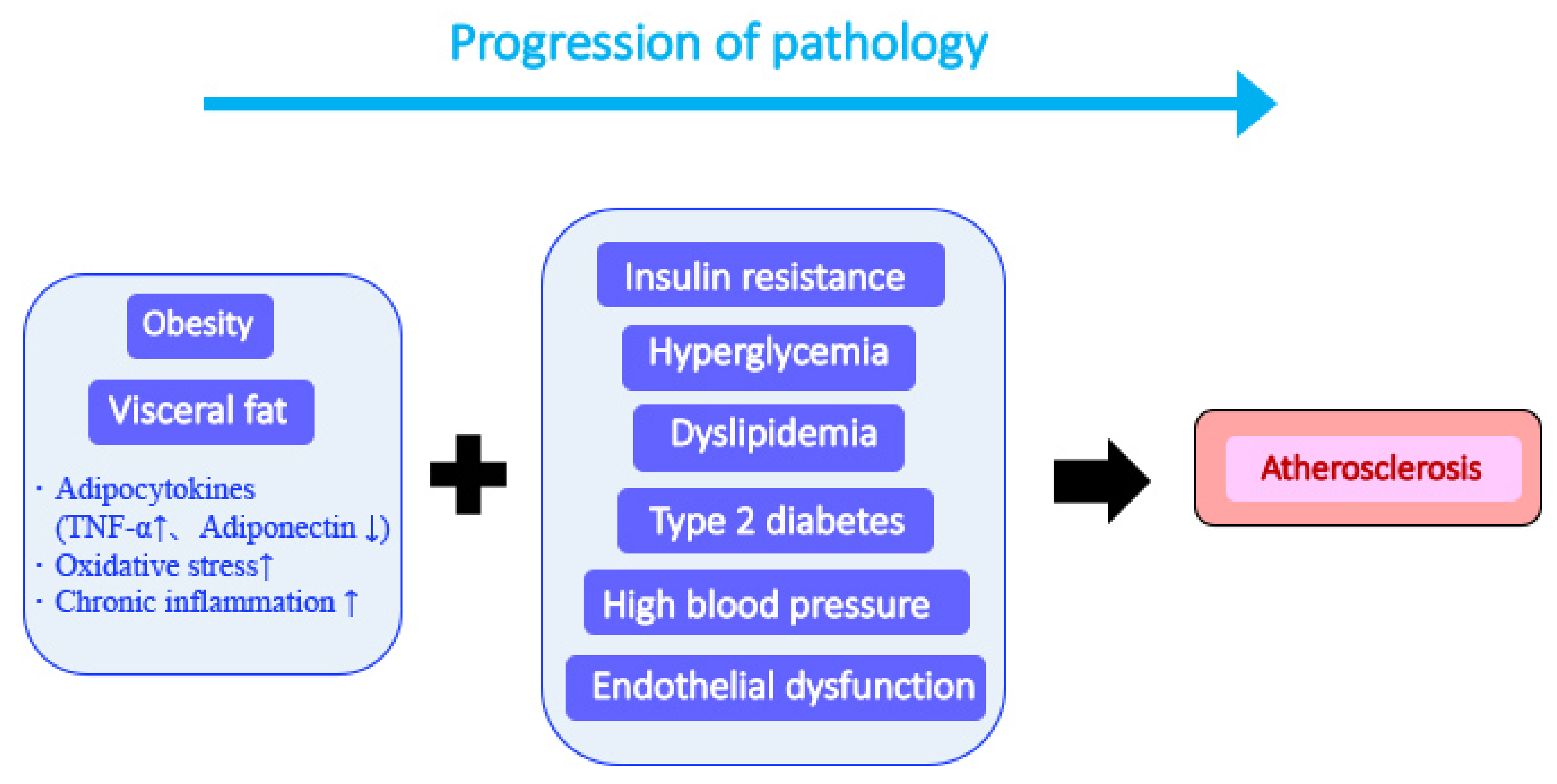
3. Metabolic Syndrome: Definition and Pathological Characteristics
3.1. Definition of Metabolic Syndrome
3.2. Pathological Characteristics of Metabolic Syndrome
3.2.1. Chronic Inflammation Induction and Persistence
3.2.2. Visceral Fat Formation
3.2.3. Induction of Insulin Resistance
3.2.4. Induction of Vascular Endothelial Cell Damage and Development of Arteriosclerosis
4. Effect of Soybean and Isoflavones on Metabolic Syndrome: Epidemiological Studies
5. Effects of Isoflavones on Metabolic Syndrome and Its Constituent Diseases
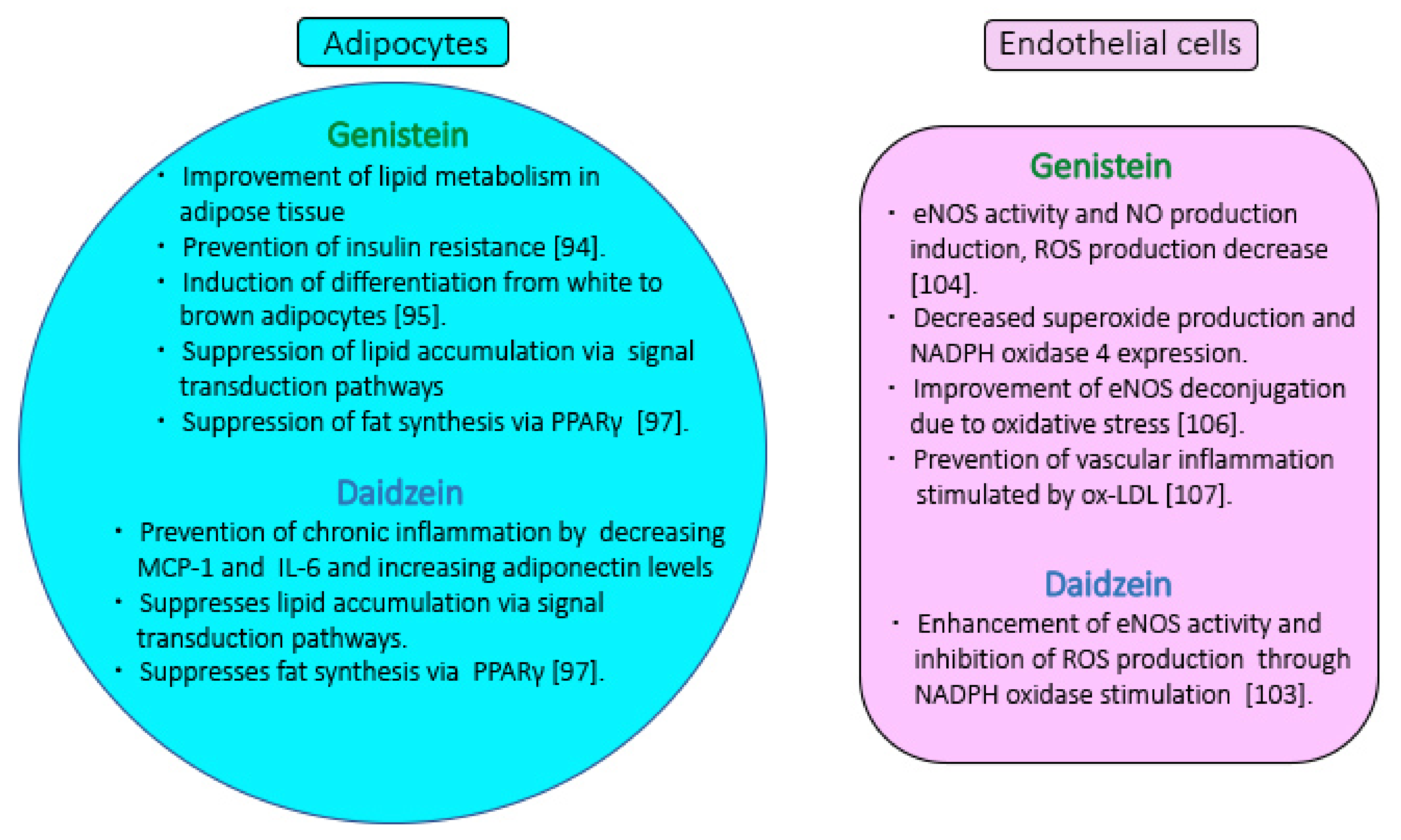
5.1. Genistein and Daidzein Effects on Adipocytes
5.2. Effect of Genistein and Daidzein on Vascular Function
5.3. Effect of Genistein and Daidzein on Type 2 Diabetes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isoflavone | Study Types | Daidzein and/or Genistein Considered | Study Design and Conditions | Evidence | Relation * | Reference |
|---|---|---|---|---|---|---|
| Genistein | Epidemiological | The genistein group (n = 28.54 mg/capsule) and the placebo group (n = 26) took two capsules daily for 12 weeks. | 54 postmenopausal women with type 2 diabetes between the ages of 47 and 69. | It improved genistein intake, T2DM postmenopausal women’s fasting blood glucose, glycated hemoglobin, serum TG, total antioxidant capacity and MDA. | + | [110] |
| Randomly assigned to two. (1) Placebo; (2) Genistein 54 mg. | 120 postmenopausal women with metabolic syndrome (placebo, n = 60; genistein 60) | Genistein intake improved the risk of T2DM and CVD in postmenopausal women with metabolic syndrome. | + | [113] | ||
| Genistein 250 mg (n = 41), placebo (n = 41) taken daily for 8 weeks. | 82 NAFLD patients | Improved fat metabolism, insulin resistance, oxidation and inflammatory index reduction in NAFLD patients. | + | [111] | ||
| Animal | Genistein (20 and 40 mg/kg), 8 weeks forced intake. | High-fat diet/streptozotocin injection in C57BL/6J mice. | Metabolic disorders of glucose and lipids, improved dysfunction of liver and colon. Changed intestinal flora and improved inflammation and insulin resistance. | + | [116] | |
| Genistein (20 mg/kg/day) | 30 Alloxan-induced diabetic rats. | Improves the harmful effects of alloxans on the pancreas and intestines. GLP-1 secretory stimulation. | + | [117] | ||
| (1) Solid diet + genistein (0.25 mg/kg/day/rat); (2) Solid feed + fructose (20% fructose); (3) Solid feed + fructose (20%) + genistein (0.25 mg/kg/day/rat). | Oxidative stress and inflammation of rat serum and liver due to fructose. | Genistein improved antioxidant, anti inflammatory, HOMA-IR and lipid status in fructose-treated rats. | + | [118] | ||
| Genistein (250 mg/kg meal). | High-fat diet/streptozotocin injection in C57BL/6J mice. | Improves hyperglycemia, glucose tolerance and blood insulin levels. Promotion of islet β-cell survival. | + | [115] | ||
| Daidzein | Epidemiological | Randomly assigned to three. In addition to 10 g of soy protein daily, (1) Placebo; (2) 50 mg of daidzein, and (3) 50 mg of genistein are administered for 24 weeks. | 165 impaired glucose regulation Chinese women aged 30–70. | Daidzein and genistein have no significant effected on glucose control and insulin sensitivity. | − | [114] |
| Animal | Daidzein (50 mg/kg daily) is forced oral administration for 12 weeks. | Ovarian ablation rats (12 weeks old). | Level weight gain, visceral fat accumulation, blood lipid, TNF-α, leptin, IL-6 level reduction.Improved insulin resistance (with HOMA-IR). | + | [119] | |
| Daidzein (0.1% in the diet) | L6 myotubes, db/db mice. KK-Ay mouse | Promotes glucose uptake, AMPK phosphorylation, and GLUT4 translocation. Improvement of gastrocnemius AMPK phosphorylation. | + | [120] | ||
| Daidzein (25, 50, 100 mg/kg), 28 days. | Streptozotocin injection in C57BL/6J mice. | Protects retinal damage due to hyperglycemia. Preventive effects on diabetic retinopathy. | + | [121] |
5.4. Effects of Genistein and Daidzein on Hyperlipidemia
5.5. Effects of Genistein and Daidzein on Hypertension
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Omoni, A.O.; Aluko, R.E. Soybean foods and their benefits: Potential mechanisms of action. Nutr. Rev. 2005, 63, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Imai, S. Soybean and Processed Soy Foods Ingredients, and Their Role in Cardiometabolic Risk Prevention. Recent Pat. Food Nutr. Agric. 2015, 7, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, J.J.; Shin, H.W.; Jung, S.; Ha, J.H. Effect of Soybean and Soybean Koji on Obesity and Dyslipidemia in Rats Fed a High-Fat Diet: A Comparative Study. Int. J. Environ. Res. Public Health 2021, 18, 6032. [Google Scholar] [CrossRef]
- Tatsumi, Y.; Morimoto, A.; Deura, K.; Mizuno, S.; Ohno, Y.; Watanabe, S. Effects of soybean product intake on fasting and postload hyperglycemia and type 2 diabetes in Japanese men with high body mass index: The Saku Study. J. Diabetes Investig. 2013, 4, 626–633. [Google Scholar] [CrossRef]
- Wei, J.L.; Wang, X.Y.; Liu, F.C.; Chen, J.C.; Cao, J.; Li, J.X.; Hu, D.S.; Shen, C.; Lu, F.H.; Zhao, Y.X.; et al. Associations of soybean products intake with blood pressure changes and hypertension incidence: The China-PAR project. J. Geriatr. Cardiol. 2020, 17, 384–392. [Google Scholar]
- Mohammadifard, N.; Sajjadi, F.; Haghighatdoost, F. Effects of soy consumption on metabolic parameters in patients with metabolic syndrome: A systematic review and meta-analysis. EXCLI J. 2021, 20, 665–685. [Google Scholar]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean Bioactive Peptides and Their Functional Properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [Green Version]
- Krizova, L.; Dadakova, K.; Kasparovska, J.; Kasparovsky, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Setchell, K.D.; Clerici, C. Equol: History, chemistry, and formation. J. Nutr. 2010, 140, 1355S–1362S. [Google Scholar] [CrossRef] [Green Version]
- Bai, W.; Wang, C.; Ren, C. Intakes of total and individual flavonoids by US adults. Int. J. Food Sci. Nutr. 2014, 65, 9–20. [Google Scholar] [CrossRef]
- Dixon, R.A.; Sumner, L.W. Legume natural products: Understanding and manipulating complex pathways for human and animal health. Plant Physiol. 2003, 131, 878–885. [Google Scholar] [CrossRef] [Green Version]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.J.; Park, J.; Kang, S.W. Prevalence of metabolic syndrome and cardiovascular risk level in a vulnerable population. Int. J. Nurs. Pract. 2015, 21, 175–183. [Google Scholar] [CrossRef]
- Tamariz, L.; Hassan, B.; Palacio, A.; Arcement, L.; Horswell, R.; Hebert, K. Metabolic syndrome increases mortality in heart failure. Clin. Cardiol. 2009, 32, 327–331. [Google Scholar] [CrossRef]
- Hu, G.; Qiao, Q.; Tuomilehto, J.; Balkau, B.; Borch-Johnsen, K.; Pyorala, K.; Group, D.S. Prevalence of the metabolic syndrome and its relation to all-cause and cardiovascular mortality in nondiabetic European men and women. Arch. Intern. Med. 2004, 164, 1066–1076. [Google Scholar] [CrossRef]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otani, H. Oxidative stress as pathogenesis of cardiovascular risk associated with metabolic syndrome. Antioxid. Redox Signal. 2011, 15, 1911–1926. [Google Scholar] [CrossRef] [PubMed]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Y.; Xu, W.; Lan, L.; Li, Y.; Wang, L.; Sun, X.; Yang, C.; Jiang, Y.; Feng, R. Dietary isoflavones intake is inversely associated with non-alcoholic fatty liver disease, hyperlipidaemia and hypertension. Int. J. Food Sci. Nutr. 2021, 72, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jungbauer, A.; Medjakovic, S. Phytoestrogens and the metabolic syndrome. J. Steroid Biochem. Mol. Biol. 2014, 139, 277–289. [Google Scholar] [CrossRef]
- Rizzo, G.; Baroni, L. Soy, Soy Foods and Their Role in Vegetarian Diets. Nutrients 2018, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, J.R.; Falvo, M.J. Protein—Which is Best? J. Sports Sci. Med. 2004, 3, 118–130. [Google Scholar]
- Mateos-Aparicio, I.; Redondo Cuenca, A.; Villanueva-Suarez, M.J.; Zapata-Revilla, M.A. Soybean, a promising health source. Nutr. Hosp. 2008, 23, 305–312. [Google Scholar]
- Messina, M.J. Legumes and soybeans: Overview of their nutritional profiles and health effects. Am. J. Clin. Nutr. 1999, 70 (Suppl. S3), 439S–450S. [Google Scholar] [CrossRef] [Green Version]
- Marlett, J.A. Content and composition of dietary fiber in 117 frequently consumed foods. J. Am. Diet. Assoc. 1992, 92, 175–186. [Google Scholar] [CrossRef]
- Weaver, C.M.; Plawecki, K.L. Dietary calcium: Adequacy of a vegetarian diet. Am. J. Clin. Nutr. 1994, 59 (Suppl. 5), 1238S–1241S. [Google Scholar] [CrossRef]
- Erdman, J.W., Jr.; Badger, T.M.; Lampe, J.W.; Setchell, K.D.; Messina, M. Not all soy products are created equal: Caution needed in interpretation of research results. J. Nutr. 2004, 134, 1229S–1233S. [Google Scholar] [CrossRef] [Green Version]
- Vitale, D.C.; Piazza, C.; Melilli, B.; Drago, F.; Salomone, S. Isoflavones: Estrogenic activity, biological effect and bioavailability. Eur. J. Drug Metab. Pharmacokinet 2013, 38, 15–25. [Google Scholar] [CrossRef]
- Biggers, J.D.; Curnow, D.H. Oestrogenic activity of subterranean clover. I. The oestrogenic activity of genistein. Biochem. J. 1954, 58, 278–282. [Google Scholar] [CrossRef] [Green Version]
- Kano, M.; Takayanagi, T.; Harada, K.; Sawada, S.; Ishikawa, F. Bioavailability of isoflavones after ingestion of soy beverages in healthy adults. J. Nutr. 2006, 136, 2291–2296. [Google Scholar] [CrossRef]
- Rufer, C.E.; Bub, A.; Moseneder, J.; Winterhalter, P.; Sturtz, M.; Kulling, S.E. Pharmacokinetics of the soybean isoflavone daidzein in its aglycone and glucoside form: A randomized, double-blind, crossover study. Am. J. Clin. Nutr. 2008, 87, 1314–1323. [Google Scholar] [CrossRef] [Green Version]
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Johnson, L.W.; Weinstock, R.S. The metabolic syndrome: Concepts and controversy. Mayo Clin. Proc. 2006, 81, 1615–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberti, K.G.; Zimmet, P.; Shaw, J. Metabolic syndrome—A new world-wide definition. A Consensus Statement from the International Diabetes Federation. Diabet. Med. 2006, 23, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Expert Panel on Detection, Evaluation Treatment of High Blood Cholesterol in, Adults, Executive Summary of The Third Report of The National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, And, Treatment of High Blood Cholesterol In Adults (Adult Treatment Panel III). JAMA 2001, 285, 2486–2497.
- Chiva-Blanch, G.; Badimon, L. Effects of polyphenol intake on metabolic syndrome: Current evidences from human trials. Oxid. Med. Cell Longev. 2017, 2017, 5812401. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Pang, D.; Luo, Q.; Chen, X.; Gao, Q.; Shi, L.; Liu, W.; Zou, Y.; Li, L.; Chen, Z. Soy Isoflavones Regulate Lipid Metabolism through an AKT/mTORC1 Pathway in Diet-Induced Obesity (DIO) Male Rats. Molecules 2016, 21, 586. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C.; et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P. Inflammation and the metabolic syndrome. Indian J. Clin. Biochem. 2011, 26, 317–318. [Google Scholar] [CrossRef] [Green Version]
- Festa, A.; D’Agostino, R., Jr.; Howard, G.; Mykkanen, L.; Tracy, R.P.; Haffner, S.M. Chronic subclinical inflammation as part of the insulin resistance syndrome: The insulin resistance atherosclerosis study (IRAS). Circulation 2000, 102, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Nagasawa, S.Y.; Ohkubo, T.; Masaki, K.; Barinas-Mitchell, E.; Miura, K.; Seto, T.; El-Saed, A.; Kadowaki, T.; Willcox, B.J.; Edmundowicz, D.; et al. Associations between inflammatory markers and subclinical atherosclerosis in middle-aged white, Japanese-American and Japanese men: The ERA-JUMP study. J. Atheroscler. Thromb. 2015, 22, 590–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aso, Y.; Wakabayashi, S.; Yamamoto, R.; Matsutomo, R.; Takebayashi, K.; Inukai, T. Metabolic syndrome accompanied by hypercholesterolemia is strongly associated with proinflammatory state and impairment of fibrinolysis in patients with type 2 diabetes: Synergistic effects of plasminogen activator inhibitor-1 and thrombin-activatable fibrinolysis inhibitor. Diabetes Care 2005, 28, 2211–2216. [Google Scholar] [PubMed] [Green Version]
- Kuo, J.Z.; Guo, X.; Klein, R.; Klein, B.E.; Genter, P.; Roll, K.; Hai, Y.; Goodarzi, M.O.; Rotter, J.I.; Chen, Y.D.; et al. Adiponectin, insulin sensitivity and diabetic retinopathy in Latinos with Type 2 diabetes. J. Clin. Endocrinol. Metab. 2015, 100, 3348–3355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, M.T.; Pagliassotti, M.J. Metabolic alterations following visceral fat removal and expansion: Beyond anatomic location. Adipocyte 2012, 1, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Smith, U. Abdominal obesity: A marker of ectopic fat accumulation. J. Clin. Investig. 2015, 125, 1790–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhassan, S.; Kiazand, A.; Balise, R.R.; King, A.C.; Reaven, G.M.; Gardner, C.D. Metabolic syndrome: Do clinical criteria identify similar individuals among overweight premenopausal women? Metabolism 2008, 57, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Abate, N. Body fat distribution and insulin resistance. Nutrients 2013, 5, 2019–2027. [Google Scholar] [CrossRef]
- Despres, J.P. Cardiovascular disease under the influence of excess visceral fat. Crit. Pathw. Cardiol. 2007, 6, 51–59. [Google Scholar] [CrossRef]
- Jung, S.H.; Ha, K.H.; Kim, D.J. Visceral fat mass has stronger associations with diabetes and prediabetes than other anthropometric obesity indicators among Korean adults. Yonsei Med. J. 2016, 57, 674–680. [Google Scholar] [CrossRef]
- Reaven, G.; Abbasi, F.; McLaughlin, T. Obesity, insulin resistance, and cardiovascular disease. Recent Prog. Horm. Res. 2004, 59, 207–223. [Google Scholar] [CrossRef]
- Le Marchand, L.; Wilkens, L.R.; Castelfranco, A.M.; Monroe, K.R.; Kristal, B.S.; Cheng, I.; Maskarinec, G.; Hullar, M.A.; Lampe, J.W.; Shepherd, J.A.; et al. Circulating biomarker score for visceral fat and risks of incident colorectal and postmenopausal breast cancer: The multiethnic cohort adiposity phenotype study. Cancer Epidemiol. Biomark. Prev. 2020, 29, 966–973. [Google Scholar] [CrossRef] [Green Version]
- Kranendonk, M.E.; van Herwaarden, J.A.; Stupkova, T.; de Jager, W.; Vink, A.; Moll, F.L.; Kalkhoven, E.; Visseren, F.L. Inflammatory characteristics of distinct abdominal adipose tissue depots relate differently to metabolic risk factors for cardiovascular disease: Distinct fat depots and vascular risk factors. Atherosclerosis 2015, 239, 419–427. [Google Scholar] [CrossRef]
- Bagyura, Z.; Kiss, L.; Lux, A.; Csobay-Novak, C.; Jermendy, A.L.; Polgar, L.; Szelid, Z.; Soos, P.; Merkely, B. Association between coronary atherosclerosis and visceral adiposity index. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 796–803. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Badimon, L.; Vilahur, G. Thrombosis formation on atherosclerotic lesions and plaque rupture. J. Intern. Med. 2014, 276, 618–632. [Google Scholar] [CrossRef]
- Otero, Y.F.; Stafford, J.M.; McGuinness, O.P. Pathway-selective insulin resistance and metabolic disease: The importance of nutrient flux. J. Biol. Chem. 2014, 289, 20462–20469. [Google Scholar] [CrossRef] [Green Version]
- Ginsberg, H.N. Insulin resistance and cardiovascular disease. J. Clin. Investig. 2000, 106, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Arcidiacono, B.; Iiritano, S.; Nocera, A.; Possidente, K.; Nevolo, M.T.; Ventura, V.; Foti, D.; Chiefari, E.; Brunetti, A. Insulin resistance and cancer risk: An overview of the pathogenetic mechanisms. Exp. Diabetes Res. 2012, 2012, 789174. [Google Scholar] [CrossRef] [Green Version]
- Bugianesi, E.; Moscatiello, S.; Ciaravella, M.F.; Marchesini, G. Insulin resistance in nonalcoholic fatty liver disease. Curr. Pharm. Des. 2010, 16, 1941–1951. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E. Insulin resistance in PCOS. Endocrine 2006, 30, 13–17. [Google Scholar] [CrossRef]
- Watson, G.S.; Craft, S. Insulin resistance, inflammation, and cognition in Alzheimer’s Disease: Lessons for multiple sclerosis. J. Neurol. Sci. 2006, 245, 21–33. [Google Scholar] [CrossRef]
- Ford, E.S. Risks for all-cause mortality, cardiovascular disease, and diabetes associated with the metabolic syndrome: A summary of the evidence. Diabetes Care 2005, 28, 1769–1778. [Google Scholar] [CrossRef] [Green Version]
- Kocelak, P.; Chudek, J.; Olszanecka-Glinianowicz, M. Prevalence of metabolic syndrome and insulin resistance in overweight and obese women according to the different diagnostic criteria. Minerva Endocrinol. 2012, 37, 247–254. [Google Scholar]
- Pearson, E.R. Type 2 diabetes: A multifaceted disease. Diabetologia 2019, 62, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Ostenson, C.G. The pathophysiology of type 2 diabetes mellitus: An overview. Acta Physiol. Scand. 2001, 171, 241–247. [Google Scholar] [CrossRef]
- Alzamil, H. Elevated Serum TNF-alpha Is Related to Obesity in Type 2 Diabetes Mellitus and Is Associated with Glycemic Control and Insulin Resistance. J. Obes. 2020, 2020, 5076858. [Google Scholar] [CrossRef] [Green Version]
- Boden, G. Effects of free fatty acids (FFA) on glucose metabolism: Significance for insulin resistance and type 2 diabetes. Exp. Clin. Endocrinol. Diabetes 2003, 111, 121–124. [Google Scholar] [CrossRef]
- Belfort, R.; Mandarino, L.; Kashyap, S.; Wirfel, K.; Pratipanawatr, T.; Berria, R.; Defronzo, R.A.; Cusi, K. Dose-response effect of elevated plasma free fatty acid on insulin signaling. Diabetes 2005, 54, 1640–1648. [Google Scholar] [CrossRef] [Green Version]
- Tam, C.S.; Viardot, A.; Clement, K.; Tordjman, J.; Tonks, K.; Greenfield, J.R.; Campbell, L.V.; Samocha-Bonet, D.; Heilbronn, L.K. Short-term overfeeding may induce peripheral insulin resistance without altering subcutaneous adipose tissue macrophages in humans. Diabetes 2010, 59, 2164–2170. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, B.; Lamendola, C. Insulin resistance syndrome. J. Cardiovasc. Nurs. 2004, 19, 339–345. [Google Scholar] [CrossRef]
- McCracken, E.; Monaghan, M.; Sreenivasan, S. Pathophysiology of the metabolic syndrome. Clin. Dermatol. 2018, 36, 14–20. [Google Scholar] [CrossRef]
- Grassi, G.; Seravalle, G.; Scopelliti, F.; Dell’Oro, R.; Fattori, L.; Quarti-Trevano, F.; Brambilla, G.; Schiffrin, E.L.; Mancia, G. Structural and functional alterations of subcutaneous small resistance arteries in severe human obesity. Obesity (Silver Spring) 2010, 18, 92–98. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef]
- Fuster, J.J.; Ouchi, N.; Gokce, N.; Walsh, K. Obesity-induced changes in adipose tissue microenvironment and their impact on cardiovascular disease. Circ. Res. 2016, 118, 1786–1807. [Google Scholar] [CrossRef] [Green Version]
- Kwaifa, I.K.; Bahari, H.; Yong, Y.K.; Noor, S.M. Endothelial dysfunction in obesity-induced inflammation: Molecular mechanisms and clinical implications. Biomolecules 2020, 10, 291. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Morioka, T.; Hayashi, M.; Kakutani, Y.; Yamazaki, Y.; Kurajoh, M.; Mori, K.; Fukumoto, S.; Shioi, A.; Shoji, T.; et al. Plasma omentin levels are inversely associated with atherosclerosis in type 2 diabetes patients with increased plasma adiponectin levels: A cross-sectional study. Cardiovasc. Diabetol. 2019, 18, 167. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Fang, S.; Liu, X.; Li, J.; Wang, X.; Cui, J.; Chen, T.; Li, Z.; Yang, F.; Tian, J.; et al. Omentin-1 protects against high glucose-induced endothelial dysfunction via the AMPK/PPARdelta signaling pathway. Biochem. Pharmacol. 2020, 174, 113830. [Google Scholar] [CrossRef]
- Nakashima, Y.; Raines, E.W.; Plump, A.S.; Breslow, J.L.; Ross, R. Upregulation of VCAM-1 and ICAM-1 at atherosclerosis-prone sites on the endothelium in the ApoE-deficient mouse. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 842–851. [Google Scholar] [CrossRef] [Green Version]
- Forstermann, U. Nitric oxide and oxidative stress in vascular disease. Pflugers Arch. 2010, 459, 923–939. [Google Scholar] [CrossRef]
- Woodman, O.L. Modulation of vasoconstriction by endothelium-derived nitric oxide: The influence of vascular disease. Clin. Exp. Pharmacol. Physiol. 1995, 22, 585–593. [Google Scholar] [CrossRef]
- Laakso, M. Cardiovascular disease in type 2 diabetes from population to man to mechanisms: The Kelly West Award Lecture 2008. Diabetes Care 2010, 33, 442–449. [Google Scholar] [CrossRef] [Green Version]
- Ruscica, M.; Pavanello, C.; Gandini, S.; Gomaraschi, M.; Vitali, C.; Macchi, C.; Morlotti, B.; Aiello, G.; Bosisio, R.; Calabresi, L.; et al. Effect of soy on metabolic syndrome and cardiovascular risk factors: A randomized controlled trial. Eur. J. Nutr. 2018, 57, 499–511. [Google Scholar] [CrossRef]
- Bakhtiari, A.; Hajian-Tilaki, K.; Omidvar, S.; Nasiri-Amiri, F. Clinical and metabolic response to soy administration in older women with metabolic syndrome: A randomized controlled trial. Diabetol. Metab. Syndr. 2019, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Azadbakht, L.; Kimiagar, M.; Mehrabi, Y.; Esmaillzadeh, A.; Hu, F.B.; Willett, W.C. Soy consumption, markers of inflammation, and endothelial function: A cross-over study in postmenopausal women with the metabolic syndrome. Diabetes Care 2007, 30, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Vallance, P.; Hingorani, A. Endothelial nitric oxide in humans in health and disease. Int. J. Exp. Pathol. 1999, 80, 291–303. [Google Scholar] [CrossRef]
- Tousoulis, D.; Kampoli, A.M.; Tentolouris, C.; Papageorgiou, N.; Stefanadis, C. The role of nitric oxide on endothelial function. Curr. Vasc. Pharmacol. 2012, 10, 4–18. [Google Scholar] [CrossRef]
- Woo, H.W.; Kim, M.K.; Lee, Y.H.; Shin, D.H.; Shin, M.H.; Choi, B.Y. Habitual consumption of soy protein and isoflavones and risk of metabolic syndrome in adults ≥ 40 years old: A prospective analysis of the Korean Multi-Rural Communities Cohort Study (MRCohort). Eur. J. Nutr. 2019, 58, 2835–2850. [Google Scholar] [CrossRef]
- Guevara-Cruz, M.; Godinez-Salas, E.T.; Sanchez-Tapia, M.; Torres-Villalobos, G.; Pichardo-Ontiveros, E.; Guizar-Heredia, R.; Arteaga-Sanchez, L.; Gamba, G.; Mojica-Espinosa, R.; Schcolnik-Cabrera, A.; et al. Genistein stimulates insulin sensitivity through gut microbiota reshaping and skeletal muscle AMPK activation in obese subjects. BMJ Open Diabetes Res. Care 2020, 8, e000948. [Google Scholar] [CrossRef] [Green Version]
- Gan, M.; Shen, L.; Wang, S.; Guo, Z.; Zheng, T.; Tan, Y.; Fan, Y.; Liu, L.; Chen, L.; Jiang, A.; et al. Genistein inhibits high fat diet-induced obesity through miR-222 by targeting BTG2 and adipor1. Food Funct. 2020, 11, 2418–2426. [Google Scholar] [CrossRef]
- Aziz, S.A.; Wakeling, L.A.; Miwa, S.; Alberdi, G.; Hesketh, J.E.; Ford, D. Metabolic programming of a beige adipocyte phenotype by genistein. Mol. Nutr. Food Res. 2017, 61, 1600574. [Google Scholar] [CrossRef]
- Grossini, E.; Farruggio, S.; Raina, G.; Mary, D.; Deiro, G.; Gentilli, S. Effects of genistein on differentiation and viability of human visceral adipocytes. Nutrients 2018, 10, 978. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, Y.; Kanatsu, J.; Toh, M.; Naka, A.; Kondo, K.; Iida, K. The dietary isoflavone daidzein reduces expression of pro-inflammatory genes through PPARα/γ and JNK pathways in adipocyte and macrophage co-cultures. PLoS ONE 2016, 11, e0149676. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Choi, Y.R.; Shim, J.; Kim, M.J. Genistin: A novel potent anti-adipogenic and anti-lipogenic agent. Molecules 2020, 25, 2042. [Google Scholar] [CrossRef]
- Squadrito, F.; Altavilla, D.; Morabito, N.; Crisafulli, A.; D’Anna, R.; Corrado, F.; Ruggeri, P.; Campo, G.M.; Calapai, G.; Caputi, A.P.; et al. The effect of the phytoestrogen genistein on plasma nitric oxide concentrations, endothelin-1 levels and endothelium dependent vasodilation in postmenopausal women. Atherosclerosis 2002, 163, 339–347. [Google Scholar] [CrossRef]
- Irace, C.; Marini, H.; Bitto, A.; Altavilla, D.; Polito, F.; Adamo, E.B.; Arcoraci, V.; Minutoli, L.; Di Benedetto, A.; Di Vieste, G.; et al. Genistein and endothelial function in postmenopausal women with metabolic syndrome. Eur. J. Clin. Investig. 2013, 43, 1025–1031. [Google Scholar] [CrossRef]
- Barsky, L.; Cook-Wiens, G.; Doyle, M.; Shufelt, C.; Rogers, W.; Reis, S.; Pepine, C.J.; Noel Bairey Merz, C. Phytoestrogen blood levels and adverse outcomes in women with suspected ischemic heart disease. Eur. J. Clin. Nutr. 2021, 75, 829–835. [Google Scholar] [CrossRef]
- Vera, R.; Galisteo, M.; Villar, I.C.; Sanchez, M.; Zarzuelo, A.; Perez-Vizcaino, F.; Duarte, J. Soy isoflavones improve endothelial function in spontaneously hypertensive rats in an estrogen-independent manner: Role of nitric-oxide synthase, superoxide, and cyclooxygenase metabolites. J. Pharmacol. Exp. Ther. 2005, 314, 1300–1309. [Google Scholar] [CrossRef]
- Si, H.; Yu, J.; Jiang, H.; Lum, H.; Liu, D. Phytoestrogen genistein up-regulates endothelial nitric oxide synthase expression via activation of cAMP response element-binding protein in human aortic endothelial cells. Endocrinology 2012, 153, 3190–3198. [Google Scholar] [CrossRef] [Green Version]
- Gawrys, J.; Gajecki, D.; Szahidewicz-Krupska, E.; Doroszko, A. Intraplatelet L-arginine-nitric oxide metabolic pathway: From discovery to clinical implications in prevention and treatment of cardiovascular disorders. Oxid. Med. Cell Longev. 2020, 2020, 1015908. [Google Scholar] [CrossRef]
- Zhang, H.P.; Zhao, J.H.; Yu, H.X.; Guo, D.X. Genistein ameliorated endothelial nitric oxidase synthase uncoupling by stimulating sirtuin-1 pathway in ox-LDL-injured HUVECs. Environ. Toxicol. Pharmacol. 2016, 42, 118–124. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Z.; Pang, X.; Yang, J.; Yu, H.; Zhang, Y.; Zhou, H.; Zhao, J. Genistein Protects Against Ox-LDL-Induced Inflammation Through MicroRNA-155/SOCS1-Mediated Repression of NF-kB Signaling Pathway in HUVECs. Inflammation 2017, 40, 1450–1459. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Goel, A.; Mehta, J.L. LOX-1: Regulation, Signaling and Its Role in Atherosclerosis. Antioxidants 2019, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Ding, M.; Pan, A.; Manson, J.E.; Willett, W.C.; Malik, V.; Rosner, B.; Giovannucci, E.; Hu, F.B.; Sun, Q. Consumption of soy foods and isoflavones and risk of type 2 diabetes: A pooled analysis of three US cohorts. Eur. J. Clin. Nutr. 2016, 70, 1381–1387. [Google Scholar] [CrossRef] [Green Version]
- Braxas, H.; Rafraf, M.; Karimi Hasanabad, S.; Asghari Jafarabadi, M. Effectiveness of Genistein Supplementation on Metabolic Factors and Antioxidant Status in Postmenopausal Women With Type 2 Diabetes Mellitus. Can. J. Diabetes 2019, 43, 490–497. [Google Scholar] [CrossRef]
- Amanat, S.; Eftekhari, M.H.; Fararouei, M.; Bagheri Lankarani, K.; Massoumi, S.J. Genistein supplementation improves insulin resistance and inflammatory state in non-alcoholic fatty liver patients: A randomized, controlled trial. Clin. Nutr. 2018, 37, 1210–1215. [Google Scholar] [CrossRef]
- Liu, Y.; Li, J.; Wang, T.; Wang, Y.; Zhao, L.; Fang, Y. The effect of genistein on glucose control and insulin sensitivity in postmenopausal women: A meta-analysis. Maturitas 2017, 97, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Marini, H.; Bitto, A.; Altavilla, D.; Polito, F.; Adamo, E.B.; D’Anna, R.; Arcoraci, V.; Burnett, B.P.; Minutoli, L.; et al. Genistein in the metabolic syndrome: Results of a randomized clinical trial. J. Clin. Endocrinol. Metab. 2013, 98, 3366–3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.B.; Chen, A.L.; Lu, W.; Zhuo, S.Y.; Liu, J.; Guan, J.H.; Deng, W.P.; Fang, S.; Li, Y.B.; Chen, Y.M. Daidzein and genistein fail to improve glycemic control and insulin sensitivity in Chinese women with impaired glucose regulation: A double-blind, randomized, placebo-controlled trial. Mol. Nutr. Food Res. 2015, 59, 240–249. [Google Scholar] [CrossRef]
- Fu, Z.; Gilbert, E.R.; Pfeiffer, L.; Zhang, Y.; Fu, Y.; Liu, D. Genistein ameliorates hyperglycemia in a mouse model of nongenetic type 2 diabetes. Appl. Physiol. Nutr. Metab. 2012, 37, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Jia, Q.; Mehmood, S.; Ma, S.; Liu, X. Genistein ameliorates inflammation and insulin resistance through mediation of gut microbiota composition in type 2 diabetic mice. Eur. J. Nutr. 2021, 60, 2155–2168. [Google Scholar] [CrossRef]
- Rehman, K.; Ali, M.B.; Akash, M.S.H. Genistein enhances the secretion of glucagon-like peptide-1 (GLP-1) via downregulation of inflammatory responses. Biomed. Pharmacother. 2019, 112, 108670. [Google Scholar] [CrossRef]
- Incir, S.; Bolayirli, I.M.; Inan, O.; Aydin, M.S.; Bilgin, I.A.; Sayan, I.; Esrefoglu, M.; Seven, A. The effects of genistein supplementation on fructose induced insulin resistance, oxidative stress and inflammation. Life Sci. 2016, 158, 57–62. [Google Scholar] [CrossRef]
- Cao, Y.K.; Zhang, S.F.; Zou, S.E.; Xia, X. Daidzein improves insulin resistance in ovariectomized rats. Climacteric 2013, 16, 111–116. [Google Scholar] [CrossRef]
- Cheong, S.H.; Furuhashi, K.; Ito, K.; Nagaoka, M.; Yonezawa, T.; Miura, Y.; Yagasaki, K. Daidzein promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and improves glucose homeostasis in Type 2 diabetic model mice. J. Nutr. Biochem. 2014, 25, 136–143. [Google Scholar] [CrossRef]
- Laddha, A.P.; Kulkarni, Y.A. Daidzein ameliorates diabetic retinopathy in experimental animals. Life Sci. 2021, 265, 118779. [Google Scholar] [CrossRef]
- Iqbal, J.; Al Qarni, A.; Hawwari, A.; Alghanem, A.F.; Ahmed, G. Metabolic Syndrome, Dyslipidemia and Regulation of Lipoprotein Metabolism. Curr. Diabetes Rev. 2018, 14, 427–433. [Google Scholar] [CrossRef]
- Zhang, T.; Chi, X.X. The effect of genistein on lipid levels and LDLR, LXRalpha and ABCG1 expression in postmenopausal women with hyperlipidemia. Diabetol. Metab. Syndr. 2019, 11, 111. [Google Scholar] [CrossRef]
- Shen, H.H.; Huang, S.Y.; Kung, C.W.; Chen, S.Y.; Chen, Y.F.; Cheng, P.Y.; Lam, K.K.; Lee, Y.M. Genistein ameliorated obesity accompanied with adipose tissue browning and attenuation of hepatic lipogenesis in ovariectomized rats with high-fat diet. J. Nutr. Biochem. 2019, 67, 111–122. [Google Scholar] [CrossRef]
- Lu, R.; Zheng, Z.; Yin, Y.; Jiang, Z. Effect of Genistein on Cholesterol Metabolism-Related Genes in HepG2 Cell. J. Food Sci. 2019, 84, 2330–2336. [Google Scholar] [CrossRef]
- Kitamura, K.; Erlangga, J.S.; Tsukamoto, S.; Sakamoto, Y.; Mabashi-Asazuma, H.; Iida, K. Daidzein promotes the expression of oxidative phosphorylation- and fatty acid oxidation-related genes via an estrogen-related receptor alpha pathway to decrease lipid accumulation in muscle cells. J. Nutr. Biochem. 2020, 77, 108315. [Google Scholar] [CrossRef]
- Bhattarai, K.; Adhikari, S.; Fujitani, M.; Kishida, T. Dietary daidzein, but not genistein, has a hypocholesterolemic effect in non-ovariectomized and ovariectomized female Sprague-Dawley rats on a cholesterol-free diet. Biosci. Biotechnol. Biochem. 2017, 81, 1805–1813. [Google Scholar] [CrossRef] [Green Version]
- Hemati, N.; Asis, M.; Moradi, S.; Mollica, A.; Stefanucci, A.; Nikfar, S.; Mohammadi, E.; Farzaei, M.H.; Abdollahi, M. Effects of genistein on blood pressure: A systematic review and meta-analysis. Food Res. Int. 2020, 128, 108764. [Google Scholar] [CrossRef]
- Silva, H. The Vascular Effects of Isolated Isoflavones-A Focus on the Determinants of Blood Pressure Regulation. Biology 2021, 10, 49. [Google Scholar] [CrossRef]
- Si, H.; Liu, D. Genistein, a soy phytoestrogen, upregulates the expression of human endothelial nitric oxide synthase and lowers blood pressure in spontaneously hypertensive rats. J. Nutr. 2008, 138, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Strilchuk, L.; Cincione, R.I.; Fogacci, F.; Cicero, A.F.G. Dietary interventions in blood pressure lowering: Current evidence in 2020. Kardiol. Pol. 2020, 78, 659–666. [Google Scholar] [CrossRef] [PubMed]



| Soy Beans | Azuki Beans | Chickpeas | Lentils | Pinto Beans | |
|---|---|---|---|---|---|
| Energy (kcal/100 g) | 446 | 329 | 378 | 352 | 347 |
| Protein (g/100 g) | 36.49 | 19.87 | 20.47 | 24.63 | 21.42 |
| Carbohydrate (g/100 g) | 30.16 | 62.90 | 62.95 | 63.35 | 62.55 |
| Fats (g/100 g) | 19.94 | 0.53 | 6.04 | 1.06 | 1.23 |
| Fiber (g/100 g) | 9.3 | 12.7 | 12.2 | 10.7 | 15.5 |
| PUFA (g/100 g) | 11.255 | 0.113 | 2.731 | 0.526 | 0.407 |
| Iron (mg/100 g) | 15.70 | 4.98 | 4.31 | 6.51 | 5.07 |
| Calcium (mg/100 g) | 277 | 66 | 57 | 35 | 113 |
| SOYBEANS and Their Processed Products | mg per 100 g |
|---|---|
| Soy beans, raw | 154.53 |
| Soy beans, roasted | 148.5 |
| Tofu | 13.1~34.78 |
| Soy milk | 0.7~10.73 |
| Miso | 41.45 |
| Natto | 82.29 |
| Shoyu (soy sauce) | 1.18 |
| Edamame | 17.92 |
| Tempeh | 3.82 |
| Okara | 9.39 |
| Soybean oil | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamagata, K.; Yamori, Y. Potential Effects of Soy Isoflavones on the Prevention of Metabolic Syndrome. Molecules 2021, 26, 5863. https://doi.org/10.3390/molecules26195863
Yamagata K, Yamori Y. Potential Effects of Soy Isoflavones on the Prevention of Metabolic Syndrome. Molecules. 2021; 26(19):5863. https://doi.org/10.3390/molecules26195863
Chicago/Turabian StyleYamagata, Kazuo, and Yukio Yamori. 2021. "Potential Effects of Soy Isoflavones on the Prevention of Metabolic Syndrome" Molecules 26, no. 19: 5863. https://doi.org/10.3390/molecules26195863
APA StyleYamagata, K., & Yamori, Y. (2021). Potential Effects of Soy Isoflavones on the Prevention of Metabolic Syndrome. Molecules, 26(19), 5863. https://doi.org/10.3390/molecules26195863