
Antilymphoma Effect of Incomptine A: In Vivo, In Silico, and Toxicological Studies

 ,
,  , ,
, ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Antilymphoma, Lethality, and Toxic Activities of Incomptine A (IA)
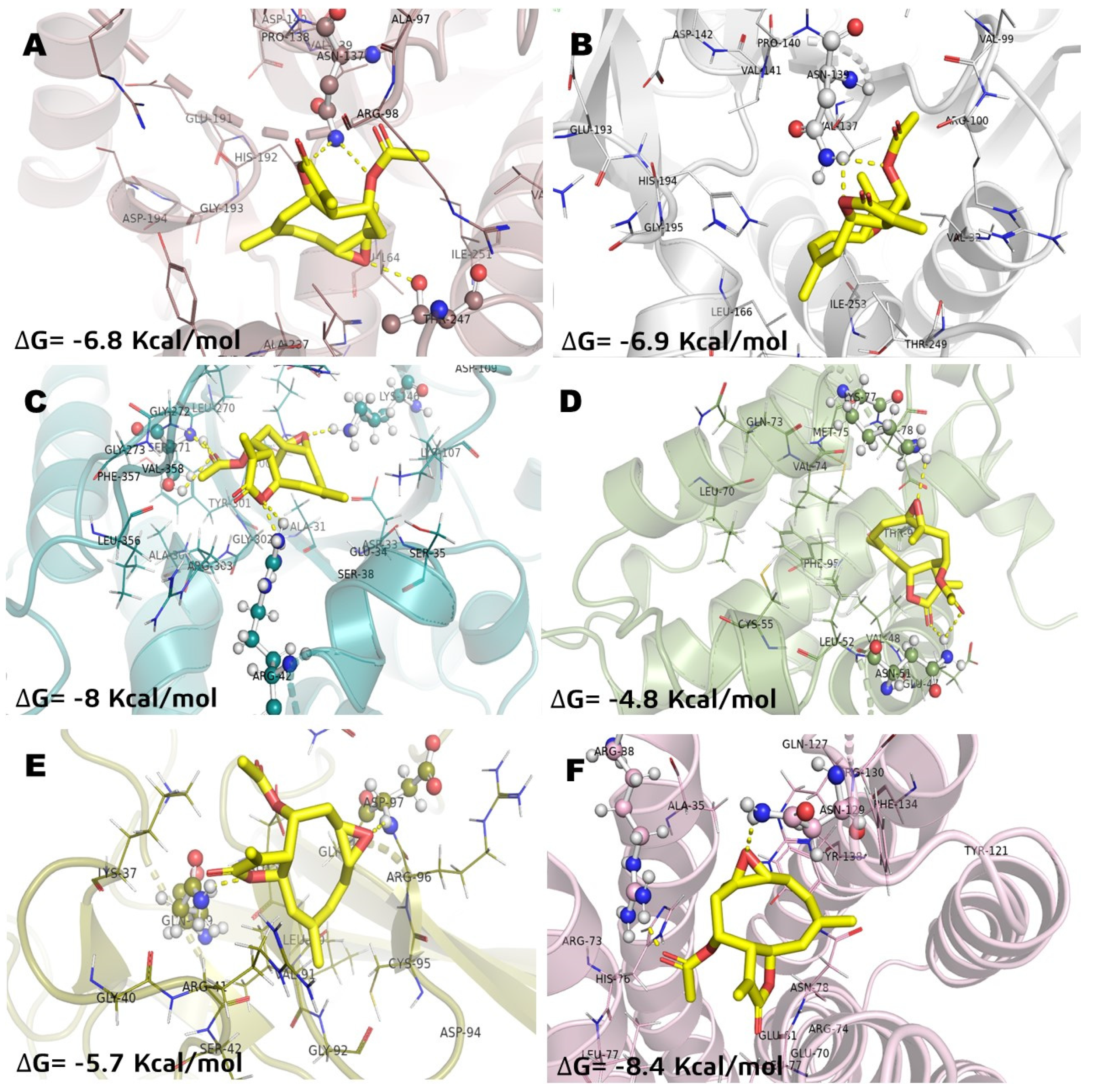
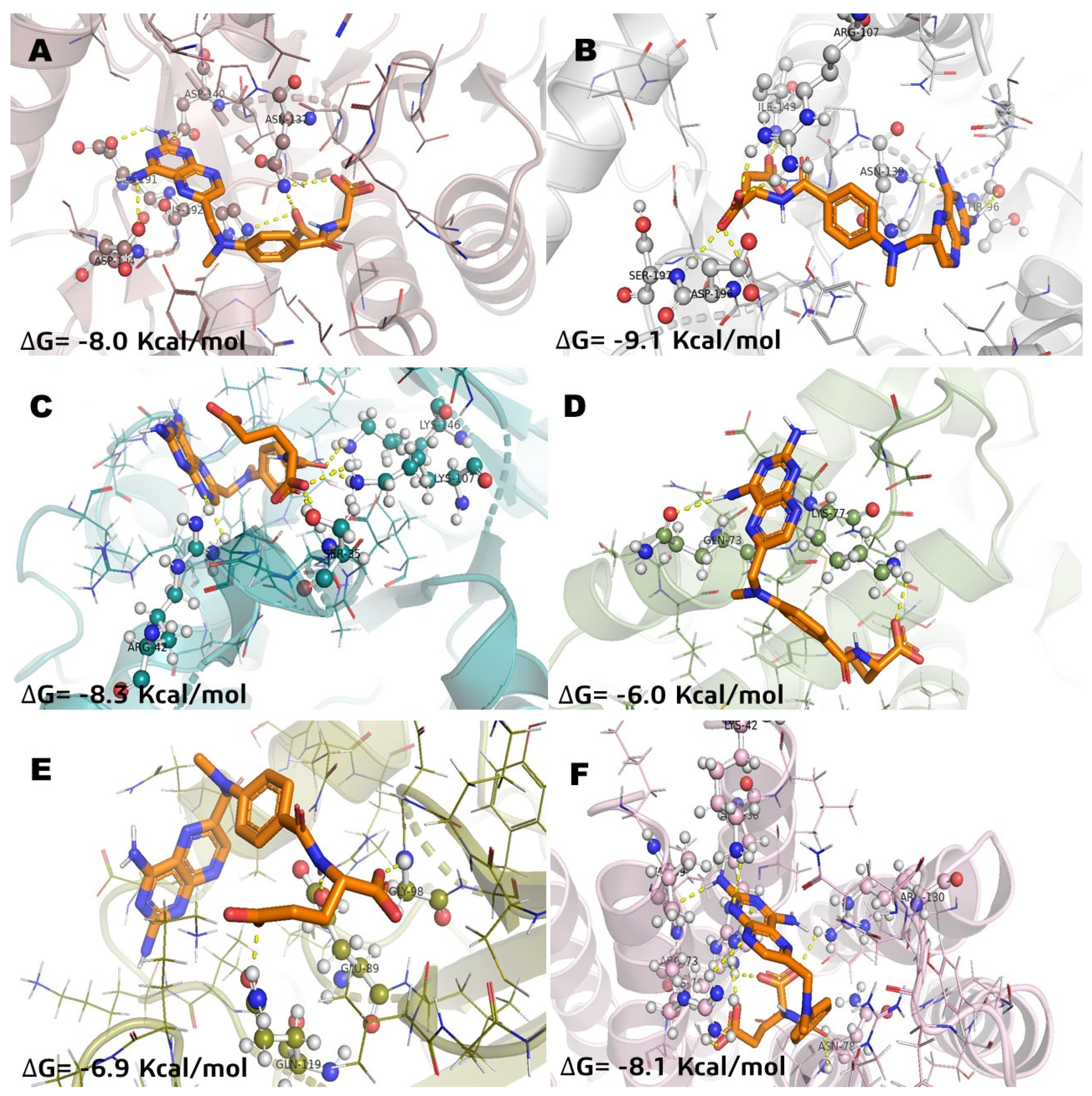
2.2. Molecular Docking Studies of Incomptine A (IA) 29 Several Selected Pharmacological Receptors Associated to Cancer
3. Discussion
4. Materials and Methods
4.1. Collection and Identification of Decachaeta incompta
4.2. Cancer Cell Line
4.3. Animals
4.4. Chemicals
5. Isolation of Incomptine A of the Aerial Parts from D. incompta
5.1. Preparation of the Aerial Parts Extract
Dichloromethane Extract: Isolation and Purification
5.2. Antilymphoma Test
5.3. Brine Shrimp Acute Lethality Test
5.4. Acute Oral Toxicity
5.5. Statistical Analysis
6. Molecular Docking Analysis
6.1. Homology Modeling of LDH B Chain
6.2. Molecular Docking of Incomptine A (IA), Parthenolide (Pa), and Methotrexate (MTX)
6.3. Docking Validation Protocol
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Available Statement
Acknowledgments
Conflicts of Interest
References
- Ren, Y.; Yu, J.; Kinghorn, A.D. Development of anticancer agents from plant-derived sesquiterpene lactones. Curr. Med. Chem. 2016, 23, 2397–2420. [Google Scholar] [CrossRef] [PubMed]
- Babaei, G.; Aliarab, A.; Abroon, S.; Rasmi, Y.; Gholizadeh-Ghaleh Aziz, S. Application of sesquiterpene lactone: A new promising way for cancer therapy based on anticancer activity. Biomed. Pharm. 2018, 106, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Calzada, F.; Yepez-Mulia, L.; Tapia-Contreras, A.; Ortega, A. Antiprotozoal and antibacterial properties of Decachaeta incompta. Rev. Lat. Qui. 2009, 37, 97–103. [Google Scholar]
- Calzada, F.; Valdés, M.; Barbosa, E.; Velázquez, C.; Bautista, E. Evaluation of antipropulsive activity of Decachaeta incompta (DC) King and Robinson and its sesquiterpene lactones on induced hyperperistalsis in rats. Phcog. Mag. 2020, 16, S272–S275. [Google Scholar] [CrossRef]
- Escandón-Rivera, S.; González-Adrade, M.; Bye, R.; Linares, E.; Navarrete, A.; Mata, R. α-glucosidase inhibitors from Brickellia cavanillesii. J. Nat. Prod. 2012, 75, 968–974. [Google Scholar] [CrossRef]
- Sepulveda-Robles, O.; Espinoza-Gutiérrez, B.; Gómez-Verjan, J.C.; Guzman-Gutierrez, S.L.; De Ita, M.; Silva-Miranda, M.; Espitia-Pinzón, C.I.; Fernandez-Ramirez, F.; Herrera-Salazar, A.; Mata-Rocha, M.; et al. Trypanocidal and toxicological assessment in vitro and in silico of three sesquiterpene lactones from Asteraceae plant species. Food Chem. Toxicol. 2019, 125, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Tassorelli, C.; Greco, R.; Morazzoni, P.; Riva, A.; Sandrini, G.; Nappi, G. Parthenolide is the component of Tanacetum parthenium that inhibits nitroglycerin-induced Fos activation: Studies in an animal model of migraine. Cephalalgia 2005, 25, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Won, Y.K.; Ong, C.W.; Shen, H.M. Anti-cancer potential of sesquiterpene lactones: Bioactivity and molecular mechanisms. Curr. Med. Chem. 2005, 5, 239–249. [Google Scholar] [CrossRef] [PubMed]
- GBD 2015 Disease and injury incidence and prevalence, collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2015, 388, 1545–1602. [Google Scholar]
- Armitage, J.O.; Gascoyne, R.D.; Lunning, M.A.; Cavalli, F. Non-Hodgkin lymphoma. Lancet 2017, 390, 298–310. [Google Scholar] [CrossRef]
- INEGI. Estadística a Propósito del día Mundial Contra el Cáncer (4 de Febrero) Datos Nacionales. 2018. Available online: https://www.inegi.org.mx/contenidos/saladeprensa/aproposito/2021/cancer2021_Nal.pdf (accessed on 21 April 2021).
- Zare-Shahneh, F.; Valiyari, S.; Barandaran, B.; Abdolalizadeh, J.; Bandehagh, A.; Azadmehr, A.; Hajiaghaee, R. Inhibitory and cytotoxic activities of Salvia officinalis L. extract on human lymphoma and leukemia cells by induction of apoptosis. Adv. Pharm. Bull. 2013, 3, 51–55. [Google Scholar]
- Bautista, E.; Calzada, F.; Yepez-Mulia, L.; Chavez-Soto, M.; Ortega, A. Incomptines C and D, two heliangolides from Decachaeta incompta and their antiprotozoal activity. Planta Med. 2012, 78, 1698–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, E.; Calzada, F.; López-Huerta, F.A.; Yepez-Mulia, L. Antiprotozoal activity of 8-acyl and 8-alkyl incomptine A analogs. Bioorg. Med. Chem. 2014, 24, 3260–3262. [Google Scholar]
- Pina-Jimenez, E.; Calzada, F.; Bautista, E.; Ordoñez-Razo, R.M.; Velazquez, C.; Barbosa, E.; Garcia-Hernandez, N. Incomptine a induces apoptosis, Ros protduction and a differential protein expression on non-Hodgkin’s lymphoma cells. Int. J. Mol. Sci. 2021, 22, 10516. [Google Scholar] [CrossRef] [PubMed]
- Casulo, C.; Rich, L.; The Leukemia & Lymphoma Society. Non-Hodgkin Lymphoma. 2013. Available online: www.LLS.org (accessed on 6 October 2021).
- Calzada, F.; Solares-Pascasio, J.I.; Valdes, M.; Garcia-Hernandez, N.; Velazquez, C.; Ordoñez-Razo, R.M.; Barbosa, E. Antilymphoma potential of the ethanol extract and rutin obtained of the leaves from Schinus molle linn. Phcog. Res. 2018, 10, 119–123. [Google Scholar] [CrossRef]
- Calzada, F.; Ramirez-Santos, J.; Valdes, M.; Garcia-Hernandez, N.; Pina-Jimenez, E.; Ordoñez-Razo, R.M. Evaluation of acute oral toxicity, brine shrimp lethality, and antilymphoma activity of geranylgeraniol and Annona macroprophyllata leaf extracts. Rev. Bras. Farm. 2020, 30, 301–304. [Google Scholar] [CrossRef]
- Meyer, B.N.; Ferrigni, N.R.; Putman, J.E.; Jacobsen, L.B.; Nichols, D.J.; Mc Laughlin, J.L. Brine shrimp: A convenient general bioassay for active plant constituents. Planta Med. 1982, 45, 31–54. [Google Scholar] [CrossRef]
- OECD. Guideline for Testing of Chemicals 423. Acute Oral Toxicity-Acute Toxic Class Method. Organización para la Cooperación y el Desarrollo Economicos, OECD/OCDE. 2001. Available online: https://www.oecd.org/chemicalsafety/risk-assessment/1948378.pdf (accessed on 21 April 2021).
- Jing, W.; Kyung-Ran, Y.; So-Youn, L.; Chang-Ho, S.; Dae-Ghon, K. Anticancer effect of the sesquiterpene lactone parthenolide. J. Biol. Chem. 2002, 277, 38954–38964. [Google Scholar]
- Ghantous, A.; Sinjab, A.; Herceg, Z.; Darwiche, N. Pharthenolide: From plant to cancer roots. Drug Dicovery Today 2013, 18, 894–905. [Google Scholar] [CrossRef]
- Norma Oficial Mexicana. NOM-062-ZOO-1999: Especificaciones Técnicas Para la Producción, Cuidado y uso de los Animales de Laboratorio. 1999. Available online: https://www.fmvz.unam.mx/fmvz/principal/archivos/062ZOO.PDF (accessed on 21 April 2021).
- Consortium, T.U. Reorganizing the protein space at the Universal Protein Resource (UniProt). Nucleic Acids Res. 2012, 40, D71–D75. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using Modeler. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti-Renom, M.A.; Stuart, A.; Fiser, A.; Sánchez, R.; Melo, F.; Sali, A. Comparative protein structure modeling of genes and genomes. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 291–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molecular Operating Environment (MOE). 2019.01. Chemical Computing Group ULC, 1010 Sherbooke St. West, Suite #910, Montreal, QC, Canada, H3A 2R7. 2021. Available online: https://www.chemcomp.com/Research-Citing_MOE.htm (accessed on 28 October 2021).
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New docking methods, expanded force field, and python bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Antilymphoma Activity EC50 (mg/kg) in Female Mice * | BS Lethality Test LD50 (µg/mL) * | Acute Oral Toxicity Test LD50 (mg/kg) in Female Mice * |
|---|---|---|---|
| DEDi | 75.0 ± 1.2 | 90.2 ± 0.77 | 1134.0 ± 0.39 |
| Incomptine A (IA) | 2.4 ± 0.10 | 16.7 ± 0.85 | 149.0 ± 0.50 |
| Methotrexate (MTX) | 1.3 ± 0.26 | 24.6 ± 0.27 | 335.0 ± 0.39 |
| Compound | Incomptine A (IA) | Parthenolide (Pa) | Methotrexate (MTX) | |
|---|---|---|---|---|
| LDHA | ΔG | −6.8 | −6.3 | −8 |
| Ki | 14.4 | 19.68 | 6.82 | |
| aa residues | Asn137, Thr247 | Arg 168, Thr247 | Asn137, Asp140 | |
| Glu191, Asp194, His192 | ||||
| LDHB | ΔG | −6.9 | −6.1 | −9.1 |
| Ki | 13.54 | 22.3 | 3.43 | |
| aa residues | Asn139 | - | Asn139, Thr96, Ile143 | |
| Asp142, Arg107, Ser197 | ||||
| ALDOA | ΔG | −8 | −8.1 | −8.3 |
| Ki | 6.82 | 6.41 | 5.65 | |
| aa residues | Ser271, Arg42 | Ser271, Gly272 | Lys107, Arg42 | |
| Lys146 | Lys146, Ser35 | |||
| NF-kB | ΔG | −5.7 | −5.7 | −6.9 |
| Ki | 28.62 | 28.62 | 13.54 | |
| aa residues | Gln199, Asp97 | Gly98, Asp97 | Gln119, Gly98 | |
| Glu89 | ||||
| BCL-2A1 | ΔG | −4.8 | −5.5 | −6 |
| Ki | 50.16 | 32.42 | 23.73 | |
| aa residues | Lys77, Asn51 | Lys77 | Lys77, Gln73 | |
| MGST1 | ΔG | −8.4 | −8.3 | −8.1 |
| Ki | 5.31 | 5.62 | 6.41 | |
| aa residues | Arg88, Arg130 | Hist76, Arg38 | Hist76, Arg130 | |
| Hist76, Arg38, Lys42 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calzada, F.; Bautista, E.; Hidalgo-Figueroa, S.; García-Hernández, N.; Barbosa, E.; Velázquez, C.; Ordoñez-Razo, R.M.; Arietta-García, A.G. Antilymphoma Effect of Incomptine A: In Vivo, In Silico, and Toxicological Studies. Molecules 2021, 26, 6646. https://doi.org/10.3390/molecules26216646
Calzada F, Bautista E, Hidalgo-Figueroa S, García-Hernández N, Barbosa E, Velázquez C, Ordoñez-Razo RM, Arietta-García AG. Antilymphoma Effect of Incomptine A: In Vivo, In Silico, and Toxicological Studies. Molecules. 2021; 26(21):6646. https://doi.org/10.3390/molecules26216646
Chicago/Turabian StyleCalzada, Fernando, Elihú Bautista, Sergio Hidalgo-Figueroa, Normand García-Hernández, Elizabeth Barbosa, Claudia Velázquez, Rosa María Ordoñez-Razo, and Angel Giovanni Arietta-García. 2021. "Antilymphoma Effect of Incomptine A: In Vivo, In Silico, and Toxicological Studies" Molecules 26, no. 21: 6646. https://doi.org/10.3390/molecules26216646
APA StyleCalzada, F., Bautista, E., Hidalgo-Figueroa, S., García-Hernández, N., Barbosa, E., Velázquez, C., Ordoñez-Razo, R. M., & Arietta-García, A. G. (2021). Antilymphoma Effect of Incomptine A: In Vivo, In Silico, and Toxicological Studies. Molecules, 26(21), 6646. https://doi.org/10.3390/molecules26216646