
Characterization of the Synergistic Effect between Ligands of Opioid and Free Fatty Acid Receptors in the Mouse Model of Colitis

 , , , , ,
, , , , ,  and
and 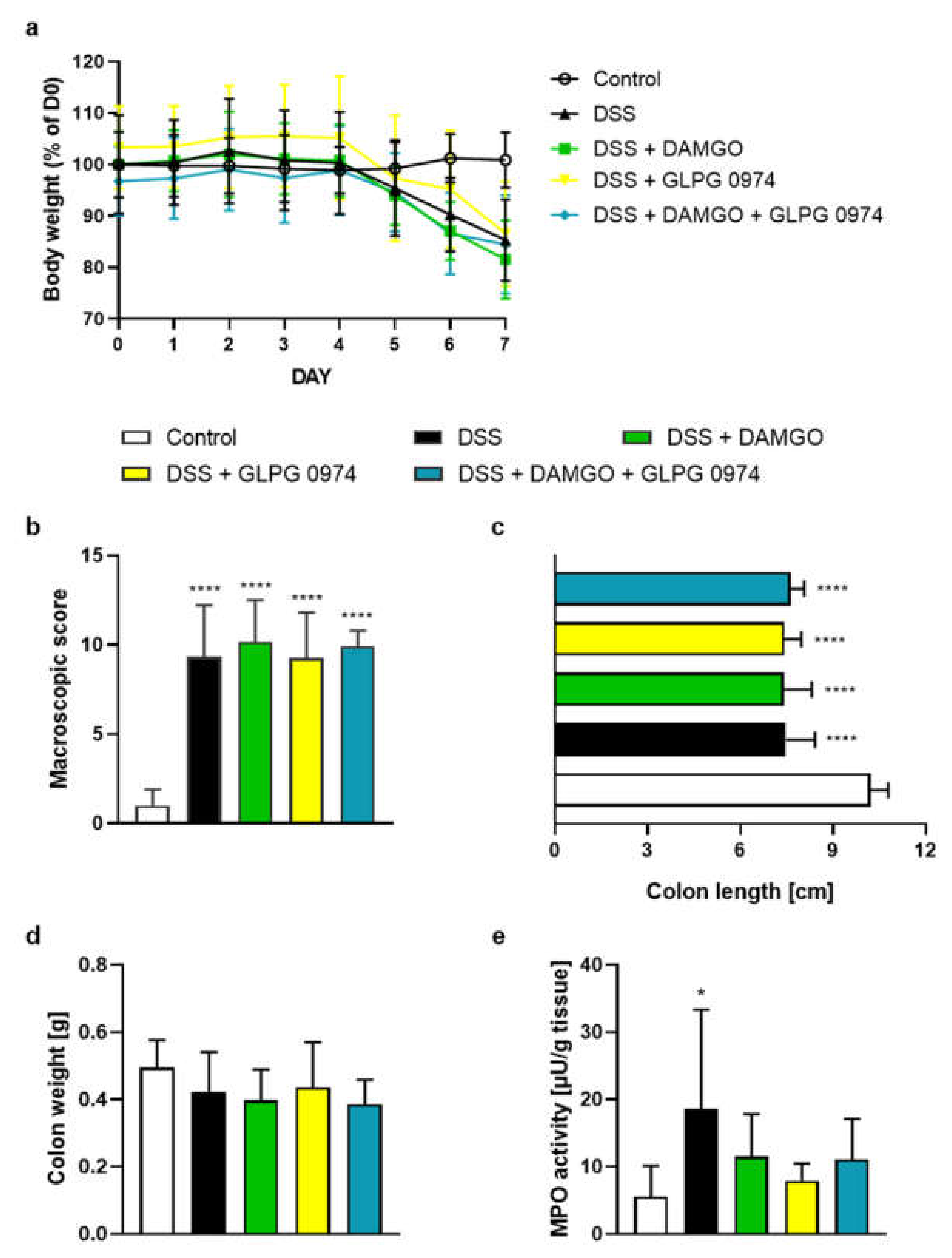{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
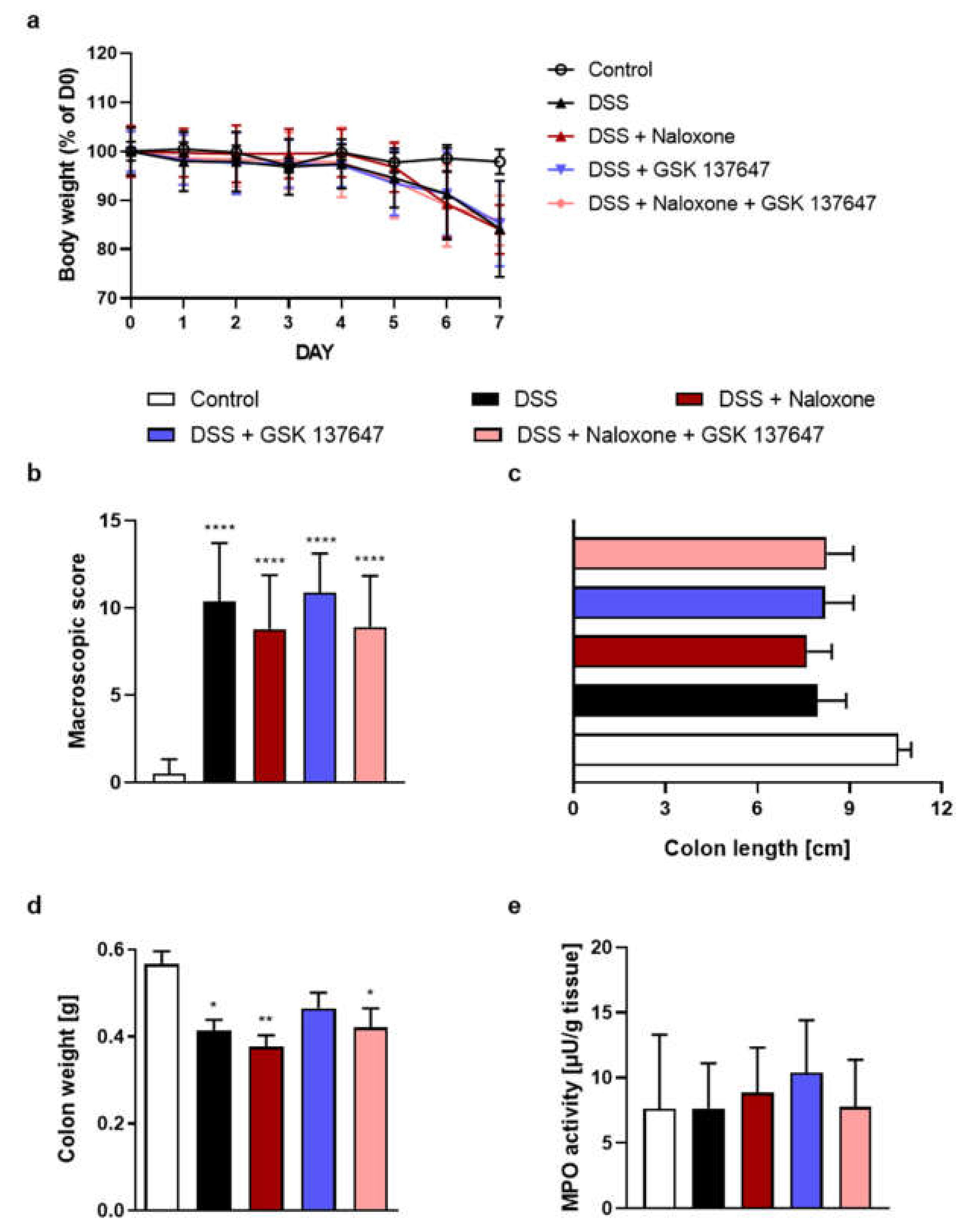
2.1. FFAR2 Antagonist and MOR Agonist Decreased MPO Activity, but Did Not Display Synergistic Anti-Inflammatory Effect
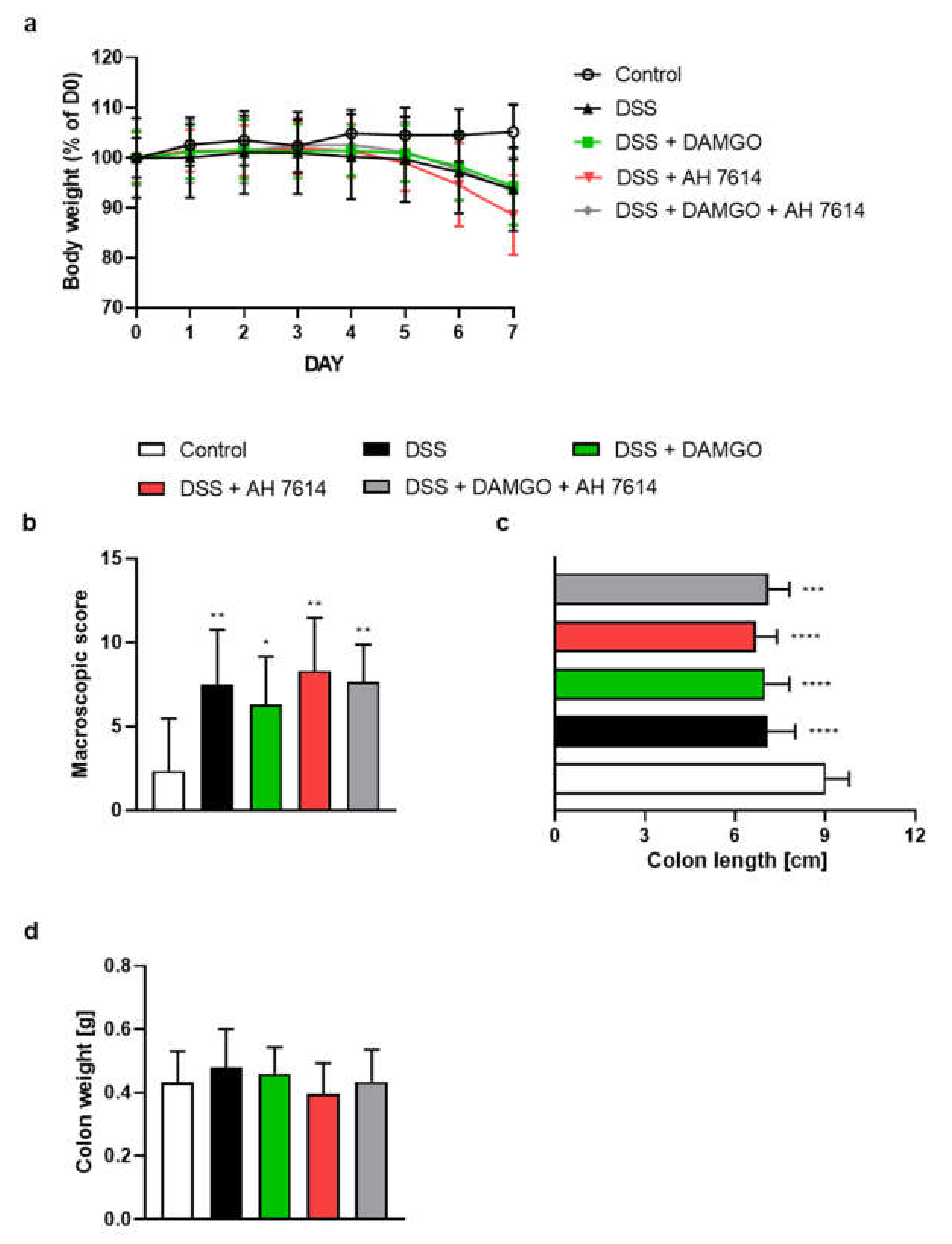
2.2. FFAR4 Antagonist Did Not Influence the Effect of MOR Agonist
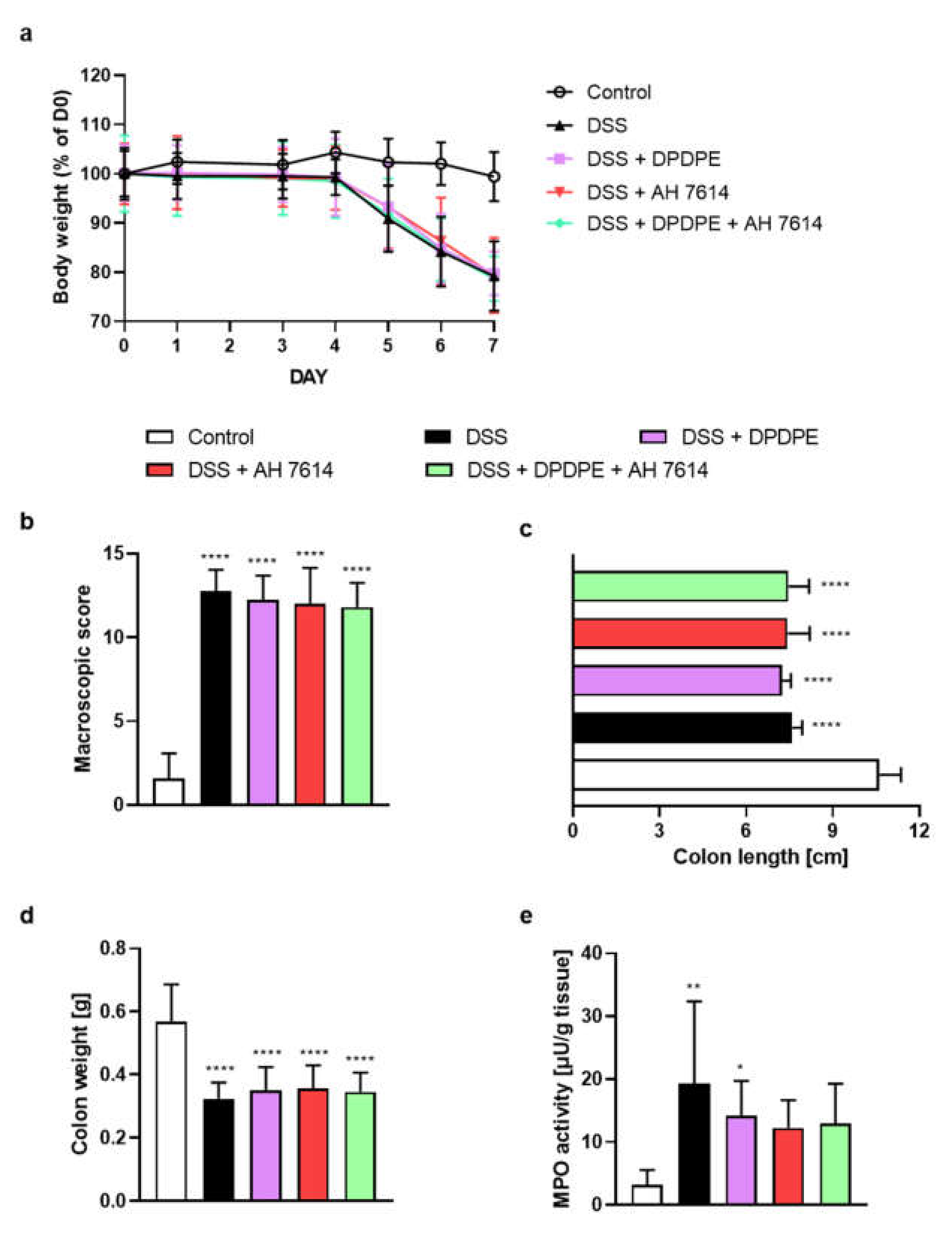
2.3. FFAR4 Antagonist Did Not Influence the Effect of DOR Agonist
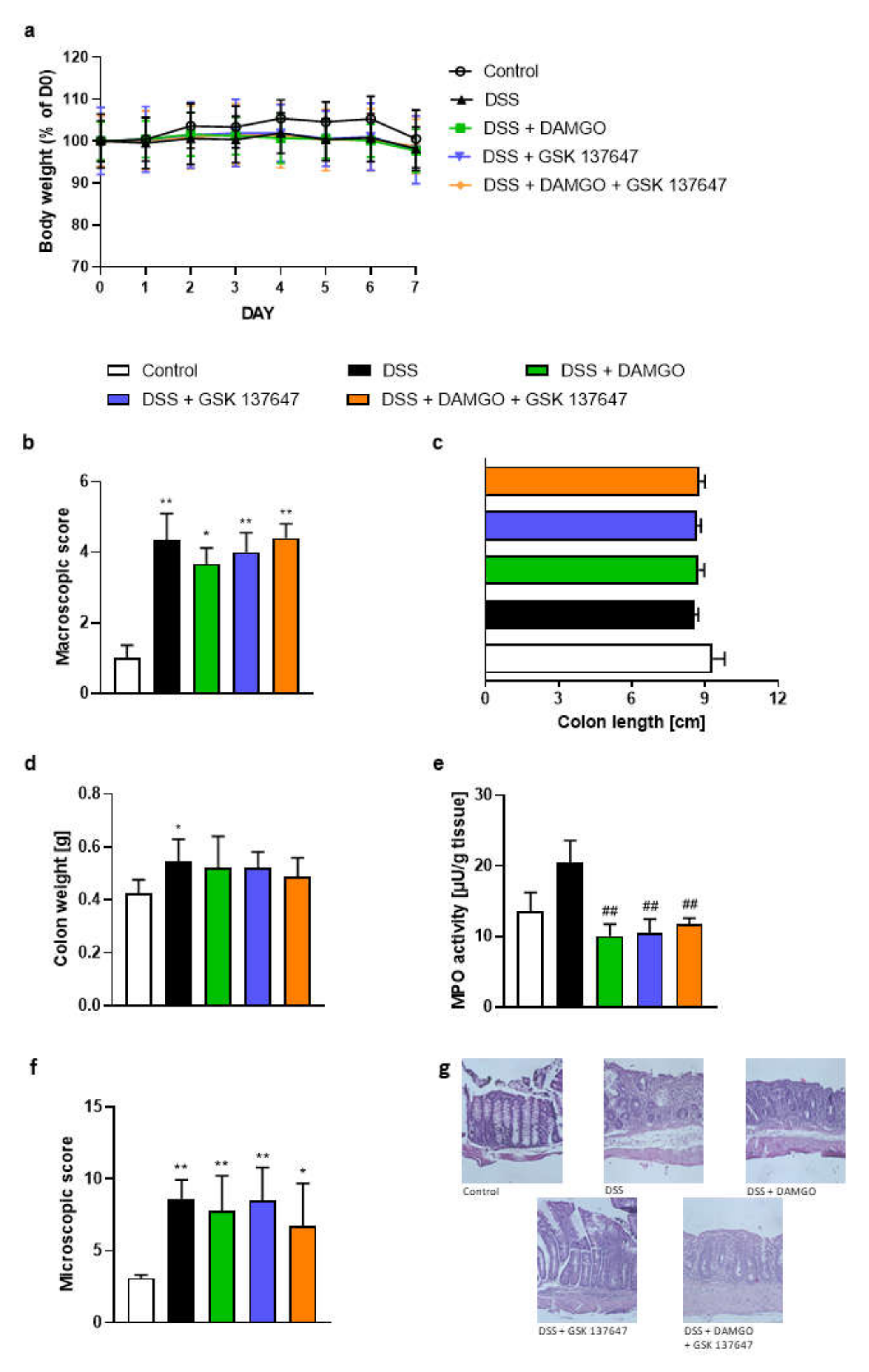
2.4. FFAR4 and MOR Agonists Decreased MPO Activity, but Did Not Display Any Synergistic Effect
2.5. FFAR4 Agonist Did Not Influence the Effect of Opioid Antagonist
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Induction of Colitis
4.4. Pharmacological Treatments
4.5. Evaluation of Colonic Damage
4.6. Determination of Tissue Myeloperoxidase Activity
4.7. Histology
4.8. Bootstrap for Hypothesis Testing in R Software
4.9. Statistics
5. Conclusions
6. Study Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sobolewska-Włodarczyk, A.; Włodarczyk, M. Pathogenesis of IBD. In Introduction to Gastrointestinal Diseases; Fichna, J., Ed.; Springer: Cham, Switzerland, 2017; Volume 1, pp. 83–93. [Google Scholar]
- Raine, T. Vedolizumab for inflammatory bowel disease: Changing the game, or more of the same? United Eur. Gastroenterol. J. 2014, 2, 333–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, L.P.; Cross, R.K. Adverse events in IBD: To stop or continue immune suppressant and biologic treatment. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 223–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milligan, G.; Stoddart, L.A.; Brown, A.J. G protein-coupled receptors for free fatty acids. Cell. Signal. 2006, 18, 1360–1365. [Google Scholar] [CrossRef] [PubMed]
- Pongkorpsakol, P.; Moonwiriyakit, A.; Muanprasat, C. Fatty acid and mineral receptors as drug targets for gastrointestinal disorders. Future Med. Chem. 2017, 9, 315–334. [Google Scholar] [CrossRef]
- Calder, P.C. Polyunsaturated fatty acids, inflammatory processes and inflammatory bowel diseases. Mol. Nutr. Food Res. 2008, 52, 885–897. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; MacKay, F.; Artis, D. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Bartoszek, A.; Von Moo, E.; Binienda, A.; Fabisiak, A.; Krajewska, J.B.; Mosińska, P.; Niewinna, K.; Tarasiuk, A.; Martemyanov, K.; Salaga, M.; et al. Free Fatty Acid Receptors as new potential therapeutic target in inflammatory bowel diseases. Pharmacol. Res. 2020, 152, 104604. [Google Scholar] [CrossRef]
- Wang, D.; Tawfik, V.L.; Corder, G.; Low, S.A.; François, A.; Basbaum, A.I.; Scherrer, G. Functional Divergence of Delta and Mu Opioid Receptor Organization in CNS Pain Circuits. Neuron 2018, 98, 90–108.e5. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, K.; Gendron, L. The Delta Opioid Receptor in Pain Control. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2017; pp. 147–177. ISBN 978-3-319-29806-1. [Google Scholar]
- Kienzl, M.; Storr, M.; Schicho, R. Cannabinoids and Opioids in the Treatment of Inflammatory Bowel Diseases. Clin. Transl. Gastroenterol. 2020, 11, e00120. [Google Scholar] [CrossRef]
- Valdez-Morales, E.; Guerrero-Alba, R.; Ochoa-Cortes, F.; Benson, J.; Spreadbury, I.; Hurlbut, D.; Miranda-Morales, M.; Lomax, A.E.; Vanner, S. Release of endogenous opioids during a chronic IBD model suppresses the excitability of colonic DRG neurons. Neurogastroenterol. Motil. 2013, 25, 5–7. [Google Scholar] [CrossRef]
- Stein, C. Targeting pain and inflammation by peripherally acting opioids. Front. Pharmacol. 2013, 4, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Philippe, D.; Dubuquoy, L.; Groux, H.; Brun, V.; Van Chuoï-Mariot, M.T.; Gaveriaux-Ruff, C.; Colombel, J.F.; Kieffer, B.L.; Desreumaux, P. Anti-inflammatory properties of the μ opioid receptor support its use in the treatment of colon inflammation. J. Clin. Investig. 2003, 111, 1329–1338. [Google Scholar] [CrossRef] [Green Version]
- DiCello, J.J.; Saito, A.; Rajasekhar, P.; Eriksson, E.M.; McQuade, R.M.; Nowell, C.J.; Sebastian, B.W.; Fichna, J.; Veldhuis, N.A.; Canals, M.; et al. Inflammation-associated changes in DOR expression and function in the mouse colon. Am. J. Physiol. Liver Physiol. 2018, 315, G544–G559. [Google Scholar] [CrossRef]
- Hasegawa, J.-I.; Loh, H.H.; Lee, N.M. Lipid Requirement for μ Opioid Receptor Binding. J. Neurochem. 1987, 49, 1007–1012. [Google Scholar] [CrossRef]
- Faden, A.; Jacobs, T.; Holaday, J. Opiate antagonist improves neurologic recovery after spinal injury. Science 1981, 211, 493–494. [Google Scholar] [CrossRef]
- Hayes, R.L.; Galinat, B.J.; Kulkarne, P.; Becker, D.P. Effects of naloxone on systemic and cerebral responses to experimental concussive brain injury in cats. J. Neurosurg. 1983, 58, 720–728. [Google Scholar] [CrossRef]
- Bakshi, R.; Newman, A.H.; Faden, A.I. Dynorphin A-(1-17) induces alterations in free fatty acids, excitatory amino acids, and motor function through an opiate-receptor-mediated mechanism. J. Neurosci. 1990, 10, 3793–800. [Google Scholar] [CrossRef] [Green Version]
- Abot, A.; Wemelle, E.; Laurens, C.; Paquot, A.; Pomie, N.; Carper, D.; Bessac, A.; Mas Orea, X.; Fremez, C.; Fontanie, M.; et al. Identification of new enterosynes using prebiotics: Roles of bioactive lipids and mu-opioid receptor signalling in humans and mice. Gut 2021, 70, 1078–1087. [Google Scholar] [CrossRef]
- Stein, C.; Kuchler, S. Non-Analgesic Effects of Opioids: Peripheral Opioid Effects on Inflammation and Wound Healing. Curr. Pharm. Des. 2012, 18, 6053–6069. [Google Scholar] [CrossRef]
- Kovarik, J.J.; Tillinger, W.; Hofer, J.; Hölzl, M.A.; Heinzl, H.; Saemann, M.D.; Zlabinger, G.J. Impaired anti-inflammatory efficacy of n-butyrate in patients with IBD. Eur. J. Clin. Investig. 2011, 41, 291–298. [Google Scholar] [CrossRef]
- Pizzonero, M.; Dupont, S.; Babel, M.; Beaumont, S.; Bienvenu, N.; Blanqué, R.; Cherel, L.; Christophe, T.; Crescenzi, B.; De Lemos, E.; et al. Discovery and optimization of an azetidine chemical series as a free fatty acid receptor 2 (FFA2) antagonist: From hit to clinic. J. Med. Chem. 2014, 57, 10044–10057. [Google Scholar] [CrossRef] [PubMed]
- Namour, F.; Galien, R.; Van Kaem, T.; Van der Aa, A.; Vanhoutte, F.; Beetens, J.; van’t Klooster, G. Safety, pharmacokinetics and pharmacodynamics of GLPG0974, a potent and selective FFA2 antagonist, in healthy male subjects. Br. J. Clin. Pharmacol. 2016, 82, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Vasu, R.; Zhang, H. The Role of Long-Chain Fatty Acids in Inflammatory Bowel Disease. Mediators Inflamm. 2019, 2019, 8495913. [Google Scholar] [CrossRef] [PubMed]
- Marton, L.T.; Goulart, R.A.; de Carvalho, A.C.A.; Barbalho, S.M. Omega fatty acids and inflammatory bowel diseases: An overview. Int. J. Mol. Sci. 2019, 20, 4851. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Moraes-Vieira, P.M.; Castoldi, A.; Aryal, P.; Yee, E.U.; Vickers, C.; Parnas, O.; Donaldson, C.J.; Saghatelian, A.; Kahn, B.B. Branched fatty acid esters of hydroxy fatty acids (FAHFAs) protect against colitis by regulating gut innate and adaptive immune responses. J. Biol. Chem. 2016, 291, 22207–22217. [Google Scholar] [CrossRef] [Green Version]
- Salaga, M.; Bartoszek, A.; Binienda, A.; Krajewska, J.B.; Fabisiak, A.; Mosińska, P.; Dziedziczak, K.; Niewinna, K.; Talar, M.; Tarasiuk, A.; et al. Activation of free fatty acid receptor 4 affects intestinal inflammation and improves colon permeability in mice. Nutrients 2021, 13, 2716. [Google Scholar] [CrossRef]
- Tsukahara, T.; Watanabe, K.; Watanabe, T.; Yamagami, H.; Sogawa, M.; Tanigawa, T.; Shiba, M.; Tominaga, K.; Fujiwara, Y.; Maeda, K.; et al. Tumor necrosis factor α decreases glucagon-like peptide-2 expression by up-regulating G-protein-coupled receptor 120 in crohn disease. Am. J. Pathol. 2015, 185, 185–196. [Google Scholar] [CrossRef]
- Oh, D.Y.; Walenta, E.; Akiyama, T.E.; Lagakos, W.S.; Lackey, D.; Pessentheiner, A.R.; Sasik, R.; Hah, N.; Chi, T.J.; Cox, J.M.; et al. A Gpr120-selective agonist improves insulin resistance and chronic inflammation in obese mice. Nat. Med. 2014, 20, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Wannick, M.; Bezdek, S.; Guillen, N.; Thieme, M.; Meshrkey, F.; Mousavi, S.; Seeling, M.; Nimmerjahn, F.; Mócsai, A.; Zillikens, D.; et al. Oral administration of the selective GPR120/FFA4 agonist compound A is not effective in alleviating tissue inflammation in mouse models of prototypical autoimmune diseases. Pharmacol. Res. Perspect. 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Stein, C.; Küchler, S. Targeting inflammation and wound healing by opioids. Trends Pharmacol. Sci. 2013, 34, 303–312. [Google Scholar] [CrossRef]
- Ciotu, C.I.; Fischer, M.J.M. Novel Analgesics with Peripheral Targets. Neurotherapeutics 2020, 17, 784–825. [Google Scholar] [CrossRef]
- Anselmi, L.; Huynh, J.; Duraffourd, C.; Jaramillo, I.; Vegezzi, G.; Saccani, F.; Boschetti, E.; Brecha, N.C.; De Giorgio, R.; Sternini, C. Activation of μ opioid receptors modulates inflammation in acute experimental colitis. Neurogastroenterol. Motil. 2015, 27, 509–523. [Google Scholar] [CrossRef] [Green Version]
- Leánez, S.; Hervera, A.; Pol, O. Peripheral antinociceptive effects of µ- and δ -opioid receptor agonists in NOS2 and NOS1 knockout mice during chronic in fl ammatory pain. Eur. J. Pharmacol. 2009, 602, 41–49. [Google Scholar] [CrossRef]
- Jiménez, N.; Puig, M.M.; Pol, O. Antiexudative Effects of Opioids and Expression of κ- and δ- Opioid Receptors during Intestinal Inflammation in Mice: Involvement of Nitric Oxide. J. Pharmacol. Exp. Ther. 2006, 316, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Galligan, J.J.; Sternini, C. Insights into the role of opioid receptors in the GI tract: Experimental evidence and therapeutic relevance. Handb. Exp. Pharmacol. 2017, 239, 363–378. [Google Scholar] [CrossRef]
- Sałaga, M.; Lewandowska, U.; Sosnowska, D.; Zakrzewski, P.K.; Cygankiewicz, A.I.; Piechota-Polańczyk, A.; Sobczak, M.; Mosinska, P.; Chen, C.; Krajewska, W.M.; et al. Polyphenol extract from evening primrose pomace alleviates experimental colitis after intracolonic and oral administration in mice. Naunyn. Schmiedebergs. Arch. Pharmacol. 2014, 387, 1069–1078. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binienda, A.; Makaro, A.; Talar, M.; Krajewska, J.B.; Tarasiuk, A.; Bartoszek, A.; Fabisiak, A.; Mosińska, P.; Niewinna, K.; Dziedziczak, K.; et al. Characterization of the Synergistic Effect between Ligands of Opioid and Free Fatty Acid Receptors in the Mouse Model of Colitis. Molecules 2021, 26, 6827. https://doi.org/10.3390/molecules26226827
Binienda A, Makaro A, Talar M, Krajewska JB, Tarasiuk A, Bartoszek A, Fabisiak A, Mosińska P, Niewinna K, Dziedziczak K, et al. Characterization of the Synergistic Effect between Ligands of Opioid and Free Fatty Acid Receptors in the Mouse Model of Colitis. Molecules. 2021; 26(22):6827. https://doi.org/10.3390/molecules26226827
Chicago/Turabian StyleBinienda, Agata, Adam Makaro, Marcin Talar, Julia B. Krajewska, Aleksandra Tarasiuk, Adrian Bartoszek, Adam Fabisiak, Paula Mosińska, Karolina Niewinna, Katarzyna Dziedziczak, and et al. 2021. "Characterization of the Synergistic Effect between Ligands of Opioid and Free Fatty Acid Receptors in the Mouse Model of Colitis" Molecules 26, no. 22: 6827. https://doi.org/10.3390/molecules26226827
APA StyleBinienda, A., Makaro, A., Talar, M., Krajewska, J. B., Tarasiuk, A., Bartoszek, A., Fabisiak, A., Mosińska, P., Niewinna, K., Dziedziczak, K., Świerczyński, M., Kordek, R., Salaga, M., & Fichna, J. (2021). Characterization of the Synergistic Effect between Ligands of Opioid and Free Fatty Acid Receptors in the Mouse Model of Colitis. Molecules, 26(22), 6827. https://doi.org/10.3390/molecules26226827