
Recording from an Identified Neuron Efficiently Reveals Hazard for Brain Function in Risk Assessment


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
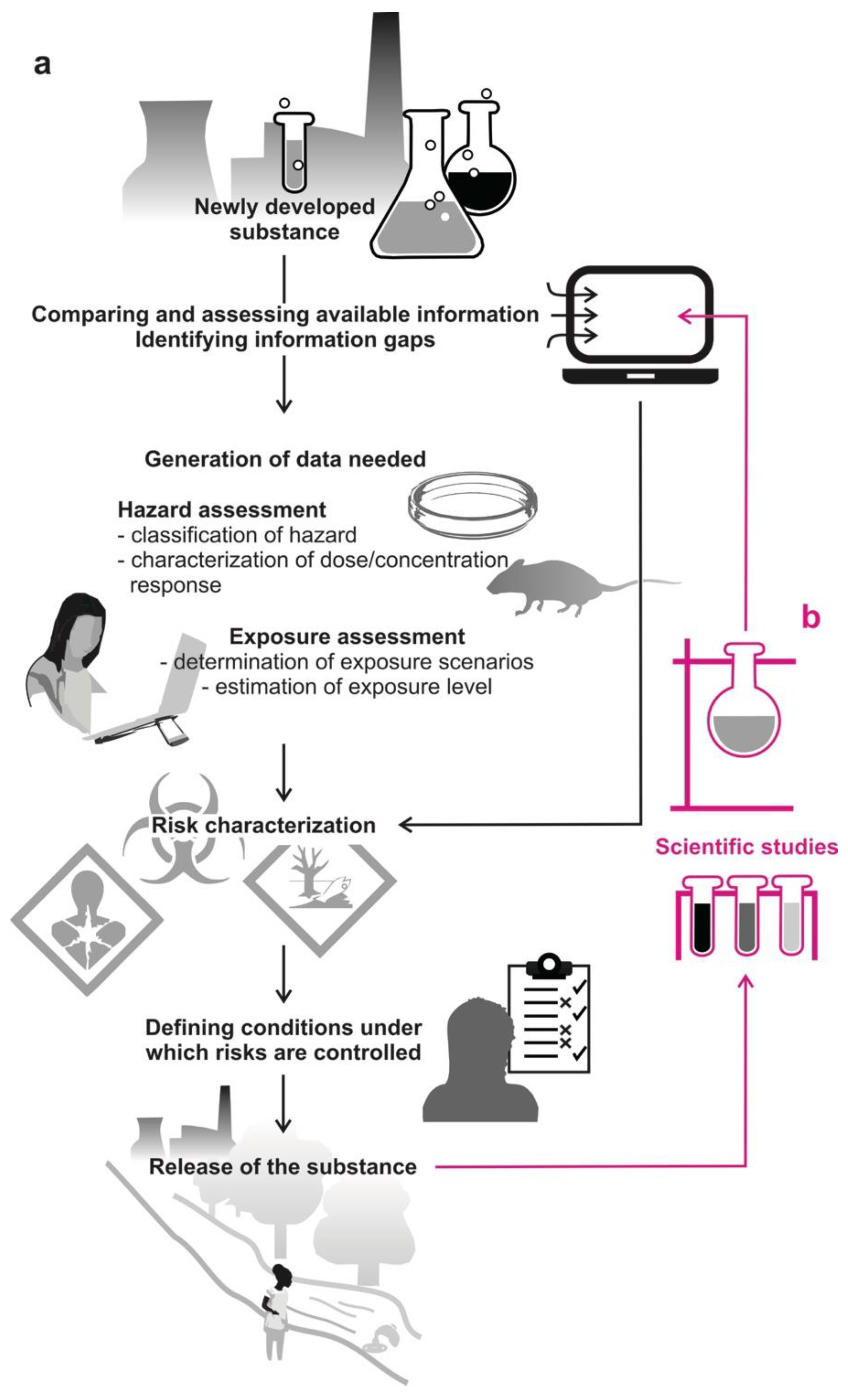
2. Standardized Tests Are Used Globally in Risk Assessment
3. Beyond Toxicity Tests: The Blind Spots in Risk Assessment
4. Opportunities in Chemical Risk Assessment Arising from an Identified Neuron in Fish
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- EPA. Guidelines for Neurotoxicity Risk Assessment. Available online: https://www.epa.gov/sites/default/files/2014-11/documents/neuro_tox.pdf (accessed on 11 October 2021).
- EPA. About the TSCA Chemical Substance Inventory. 23 September 2021. Available online: https://www.epa.gov/tsca-inventory/about-tsca-chemical-substance-inventory (accessed on 11 October 2021).
- Vogel, S.A. From the “dose makes the poison” to the “timing makes the poison”: Conceptualizing risk in the synthetic age. Environ. Hist. 2008, 13, 667–673. [Google Scholar]
- Koch, H.M.; Calafat, A.M. Human body burdens of chemicals used in plastic manufacture. Phil. Trans. R. Soc. B 2009, 364, 2063–2078. [Google Scholar] [CrossRef] [PubMed]
- Harry, G.J.; Billingsley, M.; Bruinink, A.; Campbell, I.L.; Classen, W.; Dorman, D.C.; Galli, C.; Ray, D.; Smith, R.A.; Tilson, H.A. In vitro techniques for the assessment of neurotoxicity. Environ. Health Persp. 1998, 106 (Suppl. 1), 131–158. [Google Scholar]
- Coecke, S.; Goldberg, A.M.; Allen, S.; Buzanska, L.; Calamandrei, G.; Crofton, K.; Hareng, L.; Hartung, T.; Knaut, H.; Honegger, P.; et al. Workgroup report: Incorporating in vitro alternative methods for developmental neurotoxicity into international hazard and risk assessment strategies. Environ. Health Persp. 2007, 115, 924–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bal-Price, A.; Pistollato, F.; Sachana, M.; Bopp, S.K.; Munn, S.; Worth, A. Strategies to improve the regulatory assessment of developmental neurotoxicity (DNT) using in vitro methods. Tox. Appl. Pharm. 2018, 354, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Maffini, M.V.; Neltner, T.G. Brain drain: The costs of neglected responsibilities in evaluating cumulative effects of environmental chemicals. J. Epidemiol. Community Health 2015, 69, 496–499. [Google Scholar] [CrossRef]
- ECHA. Guidance on Information Requirement and Chemical Safety Assessment. Part A: Introduction to the Guidance Document. Available online: https://echa.europa.eu/documents/10162/13643/information_requirements_part_a_en.pdf/4d25d209-00a8-4a1b-97e5-5adae231b205 (accessed on 11 October 2021).
- EPA. Human Health Risk Assessment. 2021. Available online: https://www.epa.gov/risk/human-health-risk-assessment (accessed on 11 October 2021).
- Nabholz, J.V. Environmental hazard and risk assessment under the United States toxic substances control act. Sci. Total Environ. 1991, 109, 649–665. [Google Scholar] [CrossRef]
- Eisler, R. Handbook of Chemical Risk Assessment; Lewis Publishers: Boca Raton, FL, USA, 2000; Volume 2, ISBN 1-56670-506-1. [Google Scholar]
- Bodar, C.; de Bruijn, J.; Vermeire, T.; van der Zandt, P. Trends in risk assessment of chemicals in the European Union. Hum. Ecol. Risk Assess. 2002, 8, 1825–1843. [Google Scholar] [CrossRef]
- Van Leeuwen, C.J.; Vermeire, T.G. Risk Assessment of Chemicals: An Introduction; Springer: Dordrecht, The Netherlands, 2007; ISBN 978-1-4020-6101-1. [Google Scholar]
- Kim, M.-U.; Shin, S.; Byeon, S.-H. Comparison of chemical risk assessment methods in South Korea and the United Kingdom. J. Occup. Health 2015, 57, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, T.; Claassen, L.; van Kamp, I.; Timmermans, D.R.M. ‘All chemical substances are harmful.’ public appraisal of uncertain risks of food additives and contaminants. Food Chem. Tox. 2020, 136, 110959. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, K.; Schultz, T.W.; Henry, T.; Diderich, B.; Veith, G.D. Using chemical categories to fill data gaps in hazard assessment. SAR QSAR Environ. Res. 2009, 20, 207–220. [Google Scholar] [CrossRef]
- Schultz, T.W.; Amcoff, P.; Berggren, E.; Gautier, F.; Klaric, M.; Knight, D.J.; Mahony, C.; Schwarz, M.; White, A.; Cronin, M.T.D. A strategy for structuring and reporting a read-across prediction of toxicity. Reg. Tox. Pharm. 2015, 72, 586–601. [Google Scholar] [CrossRef]
- Escher, S.E.; Kamp, H.; Bennekou, S.H.; Bitsch, A.; Fisher, C.; Graepel, R.; Hengstler, J.G.; Herzler, M.; Knight, D.; Leist, M.; et al. Towards grouping concepts on new approach methodologies in chemical hazard assessment: The read-across approach of the EU-ToxRisk project. Arch. Tox. 2019, 93, 3643–3667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ECETOC. Framework for the Integration of Human and Animal Data in Chemical Risk Assessment. Available online: https://www.ecetoc.org/wp-content/uploads/2014/08/ECETOC-TR-104.pdf (accessed on 23 October 2021).
- Embry, M.R.; Belanger, S.E.; Braunbeck, T.A.; Galay-Burgos, M.; Halder, M.; Hinton, D.E.; Léonard, M.A.; Lillicrap, A.; Norberg-King, T.; Whale, G. The fish embryo toxicity test as an animal alternative method in hazard and risk assessment and scientific research. Aqua. Tox. 2010, 97, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ciffroy, P.; Tediosi, A.; Capri, E. Modelling the Fate of Chemicals in the Environment and the Human Body; Springer: Cham, Switzerland, 2018; ISBN 978-3-319-59500-9. [Google Scholar]
- Neltner, T.G.; Alger, H.M.; Leonard, J.E.; Maffini, M.V. Data gaps in toxicity testing of chemicals allowed in food in the United States. Reprod. Tox. 2013, 42, 85–94. [Google Scholar] [CrossRef] [Green Version]
- EPA. Bisphenol A Action Plan. 29 March 2010. Available online: https://www.epa.gov/sites/default/files/2015-09/documents/bpa_action_plan.pdf (accessed on 11 October 2021).
- ECHA. Bisphenol A. Available online: https://www.echa.europa.eu/web/guest/hot-topics/bisphenol-a (accessed on 11 October 2021).
- Berge, T.L.L.; Lygre, G.B.; Jönsson, B.A.G.; Lindh, C.H.; Björkman, L. Bisphenol A concentration in human saliva related to dental polymer-based fillings. Clin. Oral Investig. 2017, 21, 2561–2568. [Google Scholar] [CrossRef]
- Wu, L.-H.; Zhang, X.-M.; Wang, F.; Gao, C.-J.; Chen, D.; Palumbo, J.R.; Guo, Y.; Zeng, E.Y. Occurrence of bisphenol S in the environment and implications for human exposure: A short review. Sci. Total Environ. 2018, 615, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Vilarinho, F.; Sendón, R.; van der Kellen, A.; Vaz, M.F.; Sanches Silva, A. Bisphenol A in food as a result of its migration from food packaging. Trends Food Sci. Technol. 2019, 91, 33–65. [Google Scholar] [CrossRef]
- Kovacic, A.; Gys, C.; Gulin, M.R.; Kosjek, T.; Heath, D.; Covaci, A.; Heath, E. The migration of bisphenols from beverage cans and reusable sports bottles. Food Chem. 2020, 331, 129786. [Google Scholar] [CrossRef]
- Thoene, M.; Dzika, E.; Gonkowski, S.; Wojtkiewicz, J. Bisphenol S in food causes hormonal and obesogenic effects comparable to or worse than bisphenol A: A literature review. Nutrients 2020, 12, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente-Martínez, Y.; Caravaca, M.; Soto-Meca, A. Determination of very low concentrations of bisphenol A in toys and baby pacifiers using dispersive liquid-liquid microextraction by in situ ionic liquid formation and high-performance liquid chromatography. Pharmaceuticals 2020, 13, 301. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S.A. The politics of plastics: The making and unmaking of bisphenol A “safety”. Amer. J. Pub. Health 2009, 99 (Suppl. 3), S559–S566. [Google Scholar] [CrossRef]
- GVR. Bisphenol A Market Size, Share & Trends Analysis Report by Application (Polycarbonates, Epoxy Resins) by Region, and Segment Forecasts, 2018–2025. Available online: https://www.grandviewresearch.com/industry-analysis/bisphenol-a-bpa-market (accessed on 23 September 2021).
- Le, H.H.; Carlson, E.M.; Chua, J.P.; Belcher, S.M. Bisphenol A is released from polycarbonate drinking bottles and mimics the neurotoxic actions of estrogen in developing cerebellar neurons. Tox. Lett. 2008, 176, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, J.E.; Kendig, E.L.; Belcher, S.M. Assessment of bisphenol A released from reusable plastic, aluminum and stainless steel water bottles. Chemosphere 2011, 85, 943–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoekstra, E.J.; Simoneau, C. Release of bisphenol A from polycarbonate—A review. Crit. Rev. Food Nutr. 2013, 53, 386–402. [Google Scholar] [CrossRef]
- Giulivo, M.; de Alda, M.L.; Capri, E.; Barceló, D. Human exposure to endocrine disrupting compounds: Their role in reproductive systems, metabolic syndrome and breast cancer. A review. Environ. Res. 2016, 151, 251–264. [Google Scholar] [CrossRef] [PubMed]
- FDA. Bisphenol A (BPA): Use in Food Contac Application. Available online: https://www.fda.gov/food/food-additives-petitions/bisphenol-bpa-use-food-contact-application (accessed on 11 October 2021).
- Arnold, S.M.; Clark, K.E.; Staples, C.A.; Klecka, G.M.; Dimond, S.S.; Caspers, N.; Hentges, S.G. Relevance of drinking water as a source of human exposure to bisphenol A. J. Expo. Sci. Environ. Epidemiol. 2013, 23, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Michalowicz, J. Bisphenol A—Sources, toxicity and biotransformation. Environ. Tox. Pharmacol. 2014, 37, 738–758. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Gao, Y.; Wu, X.; Xu, X.; Sarmah, A.K.; Bolan, N.; Gao, B.; Shaheen, S.M.; Rinklebe, J.; Ok, Y.S.; et al. A critical review on remediation of bisphenol S (BPS) contaminated water: Efficiency and mechanisms. Crit. Rev. Environ. Sci. Technol. 2020, 50, 1629802. [Google Scholar] [CrossRef]
- Catenza, C.J.; Farooq, A.; Shubear, N.S.; Donkor, K.K. A targeted review on fate, occurrence, risk and health implications of bisphenol analogues. Chemosphere 2021, 268, 129273. [Google Scholar] [CrossRef]
- Cobellis, L.; Colacurci, N.; Trabucco, E.; Carpentiero, C.; Grumetto, L. Measurement of bisphenol A and bisphenol B levels in human blood sera from healthy and endometriotic women. Biomed. Chromat. 2009, 23, 1186–1190. [Google Scholar] [CrossRef]
- Zhou, R.; Bai, Y.; Yang, R.; Zhu, Y.; Chi, X.; Li, L.; Chen, L.; Sokabe, M.; Chen, L. Abnormal synaptic plasticity in basolateral amygdala may account for hyperactivity and attention-deficit in male rat exposed perinatally to low-dose bisphenol-A. Neuropharmacology 2011, 60, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Flint, S.; Markle, T.; Thompson, S.; Wallace, E. Bisphenol A exposure, effects, and policy: A wildlife perspective. J. Environ. Manag. 2012, 104, 19–34. [Google Scholar] [CrossRef]
- Kajta, M.; Wójtowicz, A.K. Impact of endocrine-disrupting chemicals on neural development and the onset of neurological disorders. Pharm. Rep. 2013, 65, 1632–1639. [Google Scholar] [CrossRef]
- Rochester, J.R. Bisphenol A and human health: A review of the literature. Repro. Tox. 2013, 42, 132–155. [Google Scholar] [CrossRef]
- Heindel, J.J.; Newbold, R.; Schug, T.T. Endocrine disruptors and obesity. Nat. Rev. Endocrinol. 2015, 11, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Zhao, Y.; Yang, M.; Farajzadeh, M.; Pan, C.; Wayne, N.L. Actions of bisphenol A and bisphenol S on the reproductive neuroendocrine system during early development in zebrafish. Endocrinology 2016, 157, 636–647. [Google Scholar] [CrossRef] [Green Version]
- Carnevali, O.; Notarstefano, V.; Olivotto, I.; Granziano, M.; Gallo, P.; Di Marco Pisciottano, I.; Vaccari, L.; Mandich, A.; Giorgini, E.; Maradonna, F. Dietary administration of EDC mixtures: A focus on fish lipid metabolism. Aqua. Tox. 2017, 185, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Sadoul, B.; Birceanu, O.; Aluru, N.; Thomas, J.K.; Vijayan, M.M. Bisphenol A in eggs causes development-specific liver molecular reprogramming in two generations of rainbow trout. Sci. Rep. 2017, 7, 14131. [Google Scholar] [CrossRef]
- Qiu, W.; Liu, S.; Yang, F.; Dong, P.; Yang, M.; Wong, M.; Zheng, C. Metabolism disruption analysis of zebrafish larvae in response to BPA and BPA analogs based on RNA-Seq techniques. Ecotox. Environ. Safety 2019, 174, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Hwang, K.-S.; Yang, J.Y.; Chae, J.S.; Kim, G.R.; Kan, H.; Jung, M.H.; Lee, H.-Y.; Song, J.S.; Ahn, S.; et al. Neurochemical and behavioral analysis by acute exposure to bisphenol A in zebrafish larvae model. Chemosphere 2020, 239, 124751. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, E.; Schuster, S.; Machnik, P. Bisphenols exert detrimental effects on neuronal signaling in mature vertebrate brains. Comm. Biol. 2021, 4, 465. [Google Scholar] [CrossRef]
- Trullemans, L.; Koelewijn, S.-F.; Scodeller, I.; Hendrickx, T.; van Puyvelde, P.; Sels, B.F. A guide towards safe, functional and renewable BPA alternatives by rational molecular design: Structure-property and structure-toxicity relationships. Polym. Chem. 2021. [Google Scholar] [CrossRef]
- Kang, J.H.; Kondo, F.; Katayama, Y. Human exposure to bisphenol A. Toxicology 2006, 226, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Repro. Tox. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- EC. Commission Directive 2011/8/EU. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2011:026:0011:0014:EN:PDF (accessed on 11 October 2021).
- Ben-Jonathan, N.; Hugo, E.R. Bisphenols come in different flavors: Is “S” better than “A”? Endocrinology 2016, 1, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Zhang, J.; Chen, Y.; Wang, H.; Guo, M.; Wang, L.; Wang, Z.; Wu, S.; Shi, L.; Gu, A.; et al. Neurobehavioral effects of bisphenol S exposure in early life stages of zebrafish larvae (Danio rerio). Chemosphere 2019, 217, 629–635. [Google Scholar] [CrossRef] [PubMed]
- McDonough, C.; Guo, D.J.; Guo, T. Developmental toxicity of bisphenol S in Caenorhabditis elegans and NODEF mice. NeuroToxicology 2021, 87, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Fregert, S.; Rorsman, H. Hypersensitivity to epoxy resins with reference to the role played by bisphenol A. J. Investig. Dermatol. 1962, 39, 471–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaak, J.B.; Sullivan, L.J. Metabolism of bisphenol A in the rat. Toxicol. Appl. Pharmacol. 1966, 8, 175–184. [Google Scholar] [CrossRef]
- Belle, A.M.; Enright, H.A.; Sales, A.P.; Kulp, K.; Osburn, J.; Kuhn, E.A.; Fischer, N.O.; Wheeler, E.K. Evaluation of in vitro neuronal platforms as surrogates for in vivo whole brain systems. Sci. Rep. 2018, 8, 10820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machnik, P.; Schirmer, E.; Glück, L.; Schuster, S. Recordings in an integrating central neuron provide a quick way for identifying appropriate anaesthetic use in fish. Sci. Rep. 2018, 8, 17541. [Google Scholar] [CrossRef]
- Davis, G.W. Homeostatic signaling and the stabilization of neuronal function. Neuron 2013, 80, 718–728. [Google Scholar] [CrossRef] [Green Version]
- Keck, T.; Keller, G.B.; Jacobsen, R.I.; Eysel, U.T.; Bonhoeffer, T.; Hübener, M. Synaptic scaling and homeostatic plasticity in the mouse visual cortex in vivo. Neuron 2013, 80, 327–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, F.; Li, T.; Gong, H.; Chen, Z.; Jin, Y.; Xu, G.; Wang, M. Bisphenol A impairs synaptic plasticity by both pre- and postsynaptic mechanisms. Adv. Sci. 2017, 4, 1600493. [Google Scholar] [CrossRef] [Green Version]
- Furshpan, E.J.; Furukawa, T. Intracellular and extracellular responses of the several regions of the Mauthner cell of goldfish. J. Neurophysiol. 1962, 25, 732–771. [Google Scholar] [CrossRef]
- Machnik, P.; Leupolz, K.; Feyl, S.; Schulze, W.; Schuster, S. The Mauthner cell in a fish with top-performance and yet flexibly tuned C-starts. II Physiology. J. Exp. Biol. 2018, 221, jeb175588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecker, A.; Schulze, W.; Oster, J.; Richter, D.O.; Schuster, S. Removing a single neuron in a vertebrate brain forever abolishes an essential behavior. Proc. Natl. Acad. Sci. USA 2020, 117, 3254–3260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, J.D.; Alderton, W.K. Zebrafish: An in vivo model for the study of neurological diseases. Neuropsych. Dis. Treat. 2008, 4, 567–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozol, R.A.; Abrams, A.J.; James, D.M.; Bulgo, E.; Yan, Q.; Dallman, J.E. Function over form: Modeling groups of inherited neurological conditions in zebrafish. Front. Mol. Neurosci. 2016, 9, 55. [Google Scholar] [CrossRef]
- Fontana, B.D.; Mezzomo, N.J.; Kalueff, A.V.; Rosemberg, D.B. The developing utility of zebrafish models of neurological and neuropsychiatric disorders: A critical review. Exp. Neurol. 2018, 299, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Shams, S.; Rihel, J.; Ortiz, J.G.; Gerlai, R. The zebrafish as a promising tool for modeling human brain disorders: A review based upon an IBNS symposium. Neurosci. Biobehav. Rev. 2018, 85, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Zottoli, S.J.; Faber, D.S. The Mauthner cell: What has it taught us? Neuroscientist 2000, 6, 26–38. [Google Scholar] [CrossRef]
- Korn, H.; Faber, D.S. The Mauthner cell half a century later: A neurobiological model for decision-making? Neuron 2005, 47, 13–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machnik, P.; Schuster, S. Recording from an Identified Neuron Efficiently Reveals Hazard for Brain Function in Risk Assessment. Molecules 2021, 26, 6935. https://doi.org/10.3390/molecules26226935
Machnik P, Schuster S. Recording from an Identified Neuron Efficiently Reveals Hazard for Brain Function in Risk Assessment. Molecules. 2021; 26(22):6935. https://doi.org/10.3390/molecules26226935
Chicago/Turabian StyleMachnik, Peter, and Stefan Schuster. 2021. "Recording from an Identified Neuron Efficiently Reveals Hazard for Brain Function in Risk Assessment" Molecules 26, no. 22: 6935. https://doi.org/10.3390/molecules26226935
APA StyleMachnik, P., & Schuster, S. (2021). Recording from an Identified Neuron Efficiently Reveals Hazard for Brain Function in Risk Assessment. Molecules, 26(22), 6935. https://doi.org/10.3390/molecules26226935