
Insights on the Inhibitory Power of Flavonoids on Tyrosinase Activity: A Survey from 2016 to 2021


 and
and 
Abstract
:1. Introduction
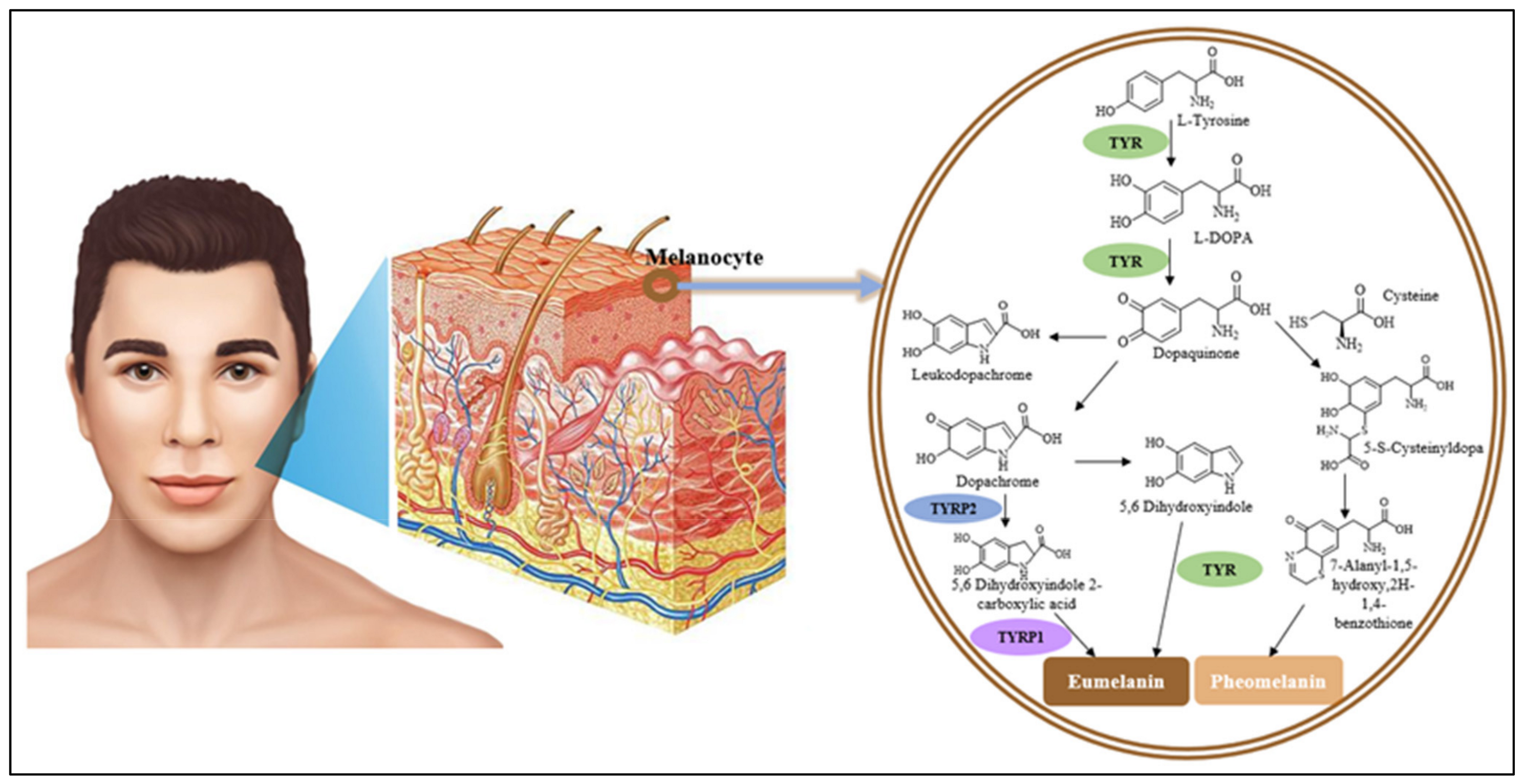
2. The Role of Tyrosinase in the Pathway of Melanin Biosynthesis
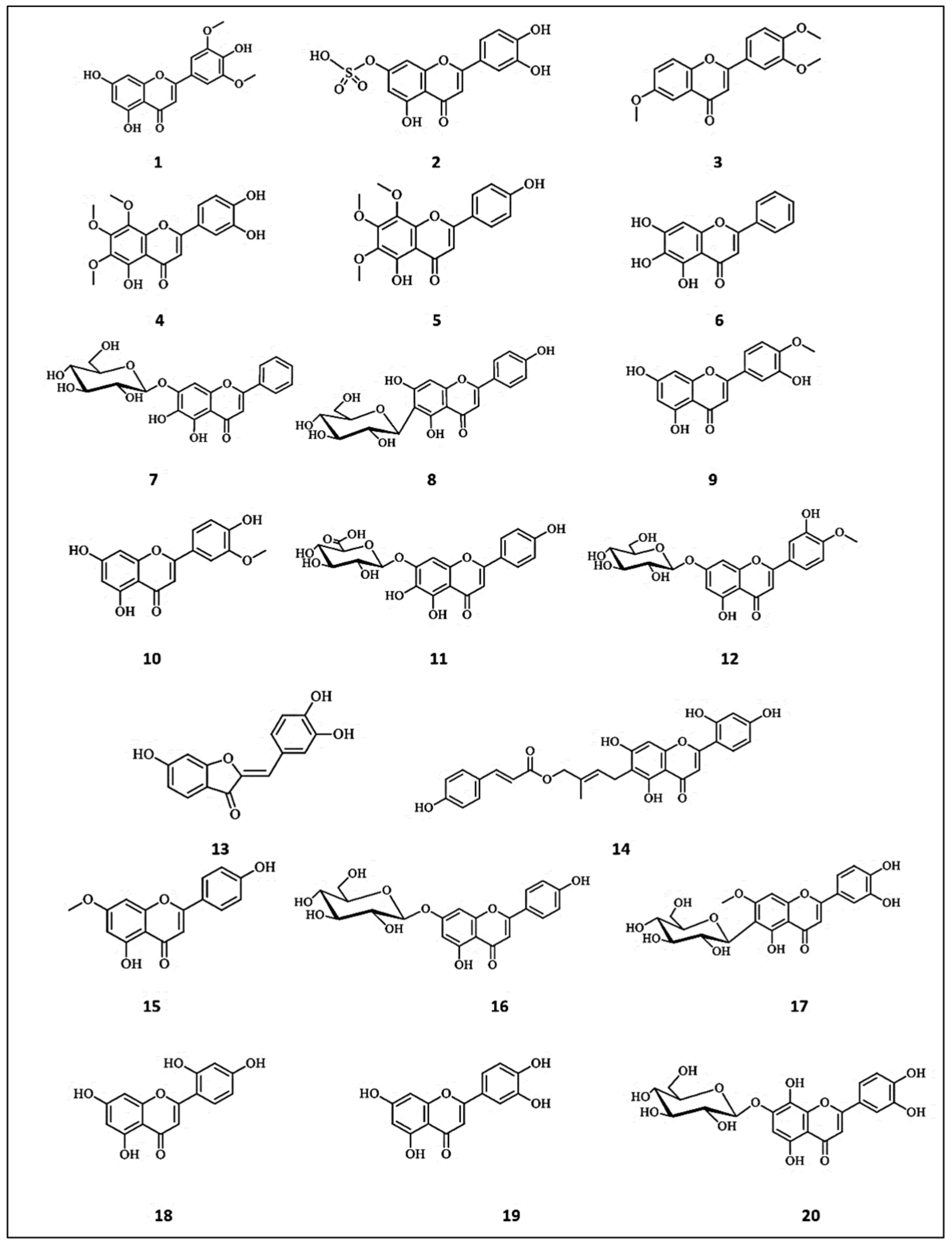
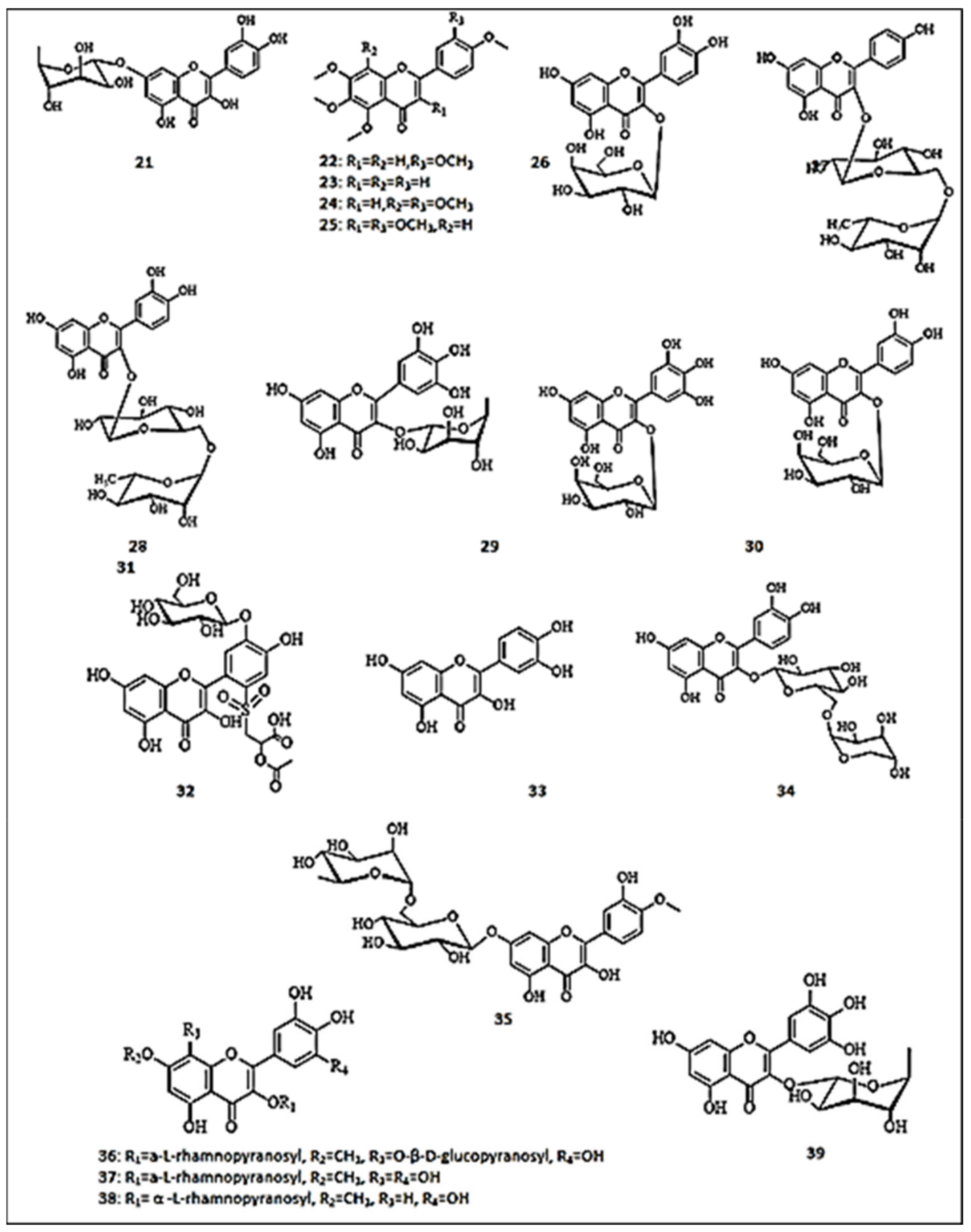
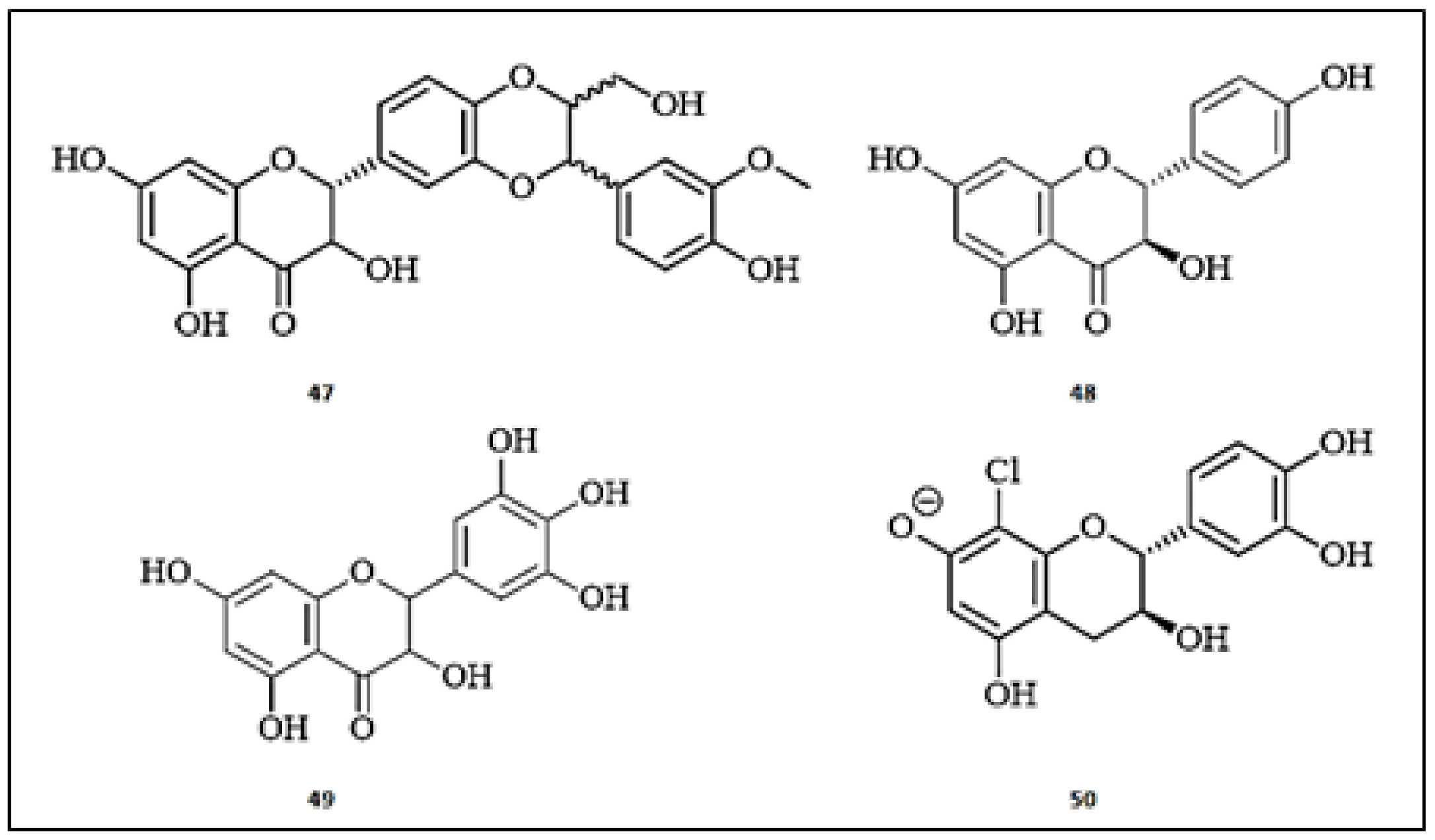
3. Flavonoids with Promising Anti-Tyrosinase Activities
3.1. Flavones
3.2. Flavonols
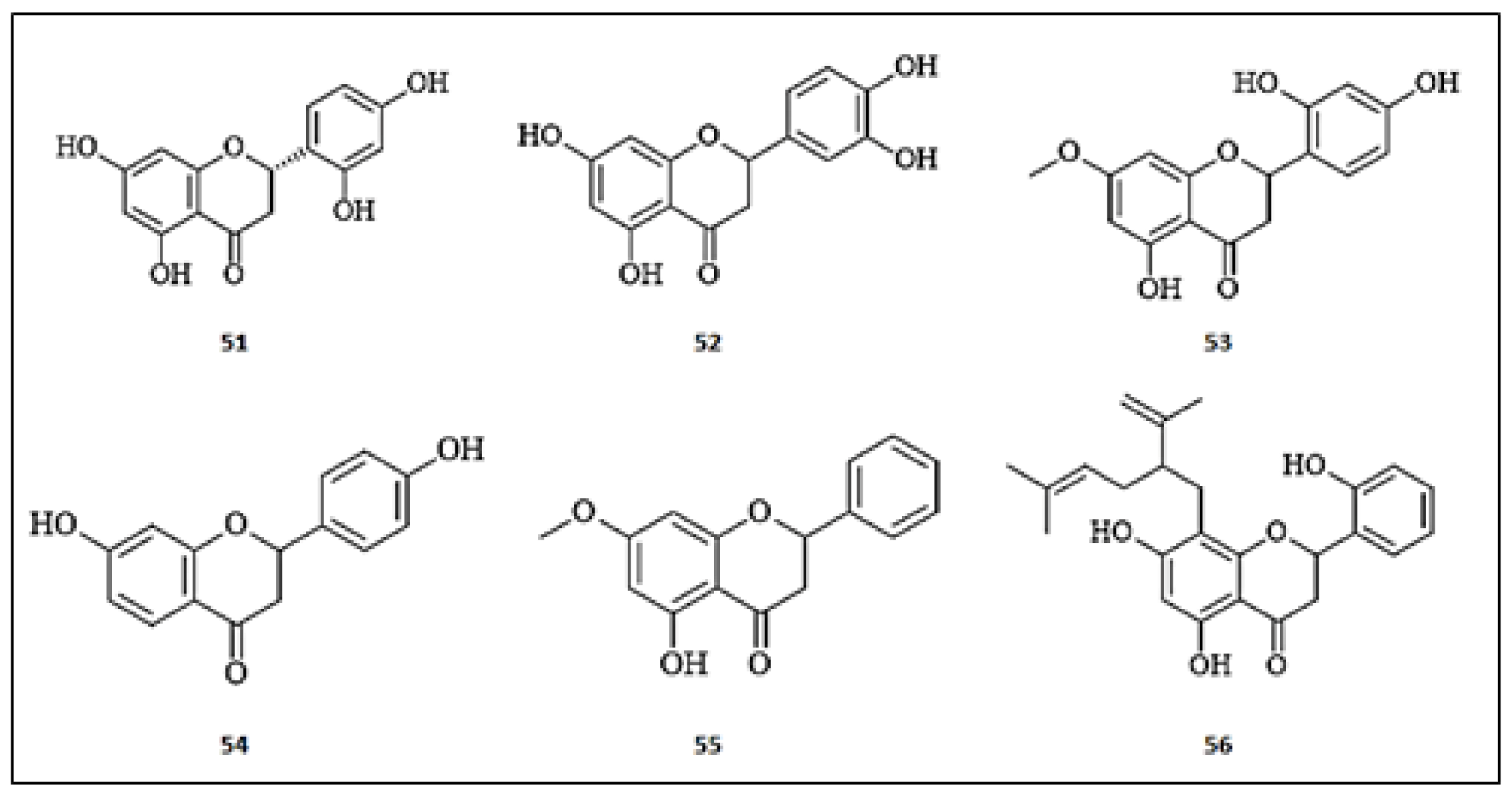
3.3. Isoflavones
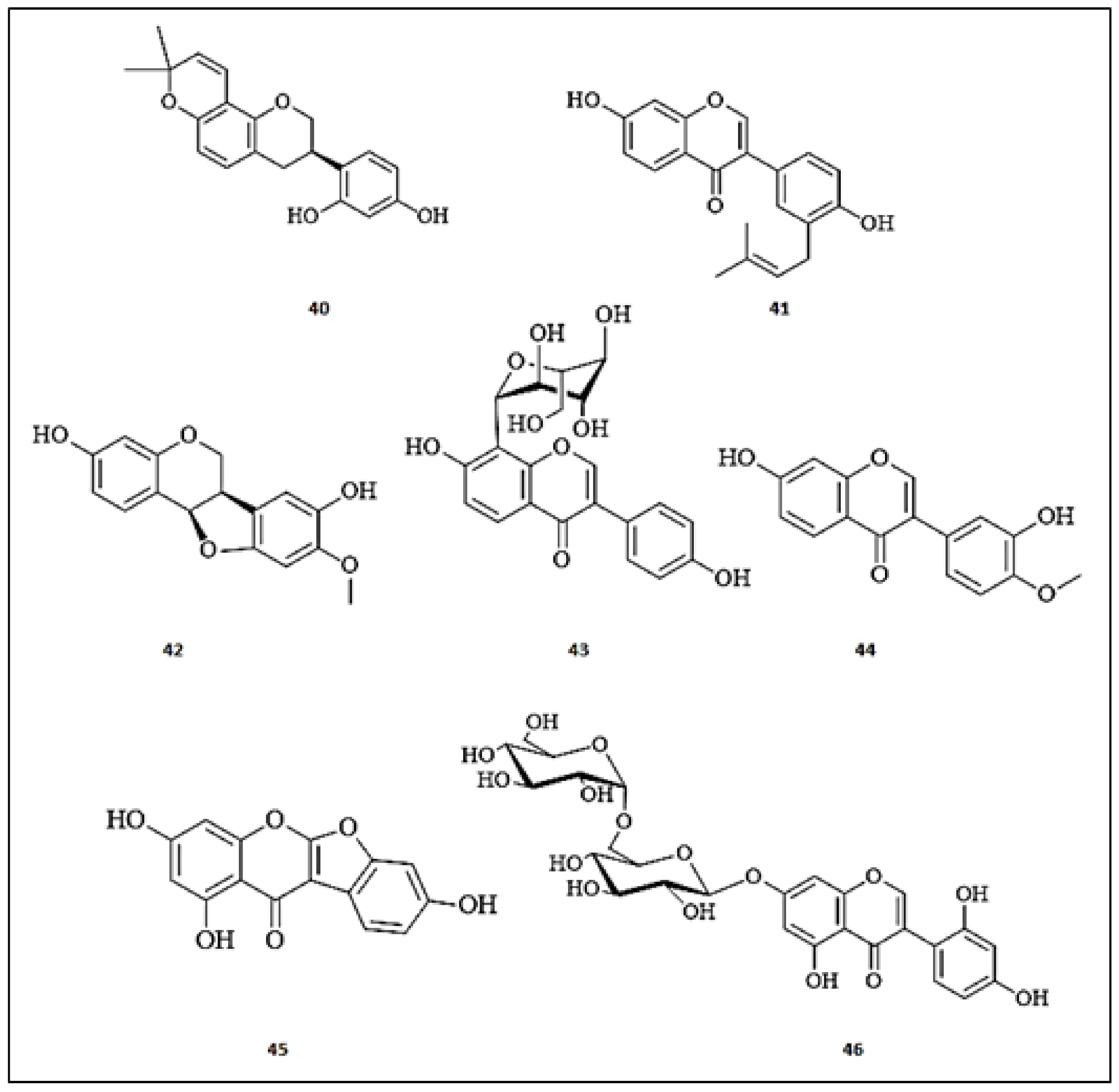
3.4. Flavan-3-ols
3.5. Flavanones
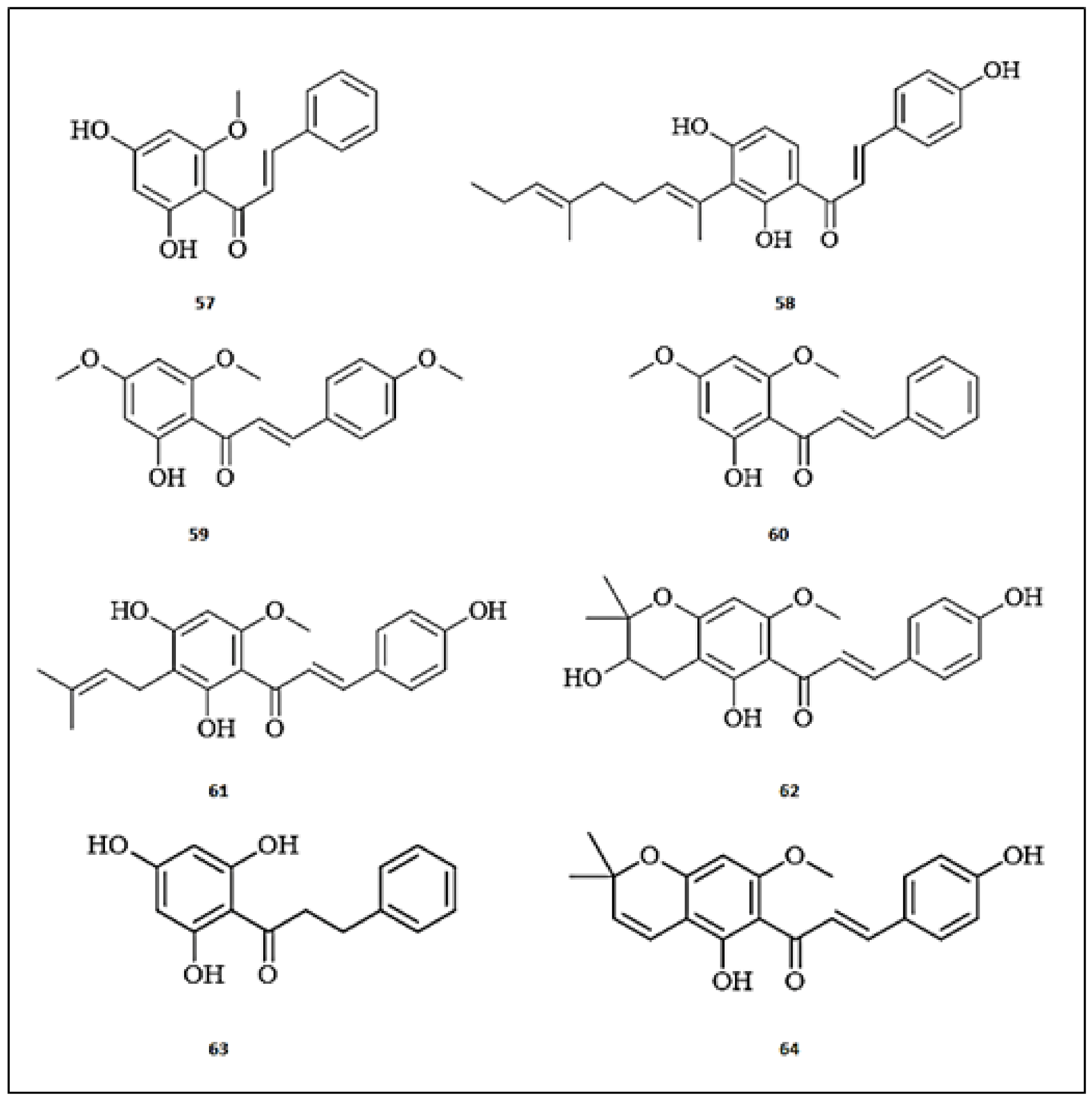
3.6. Chalcones
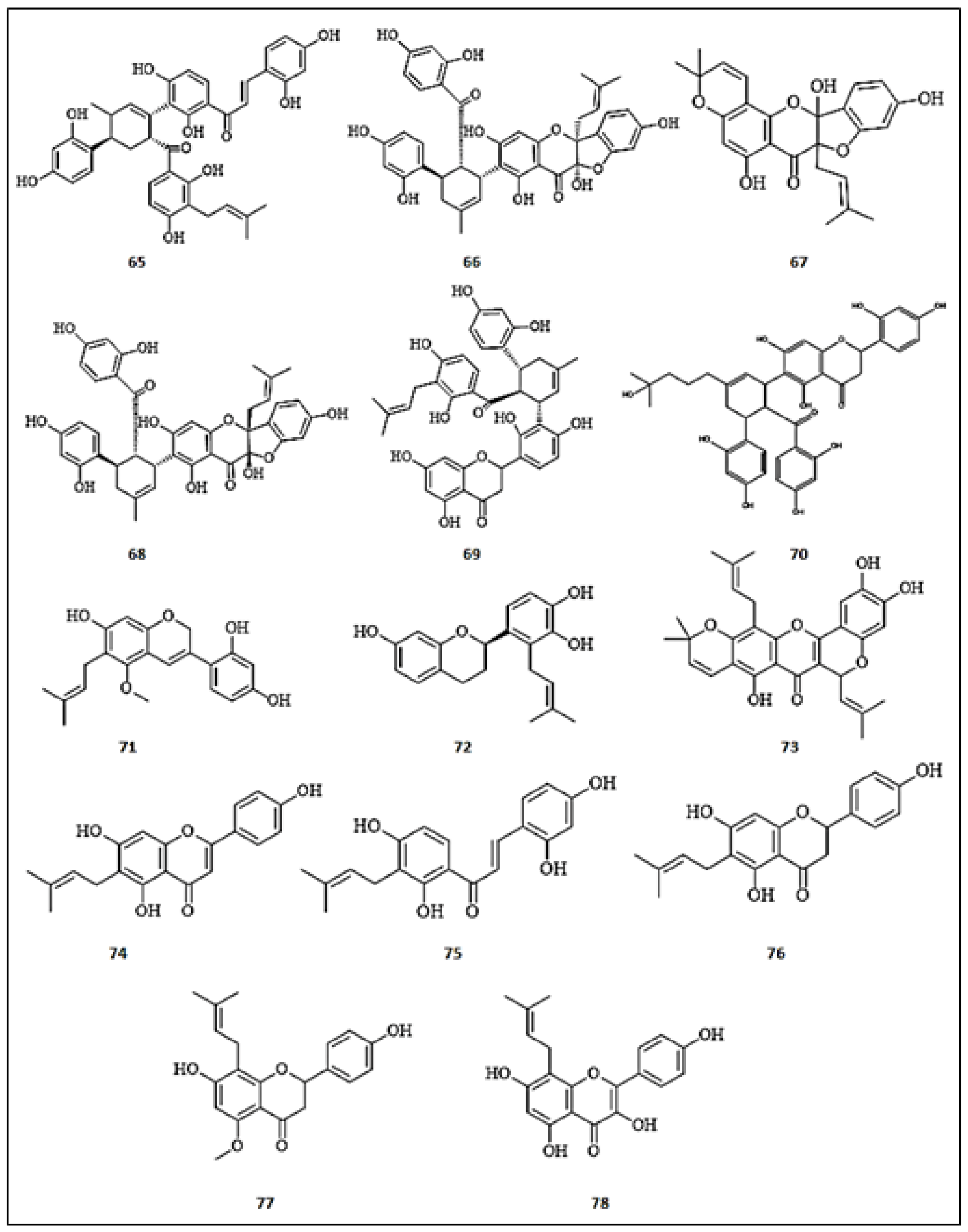
3.7. Prenylated Flavonoids
3.8. Biflavonoids
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gou, L.; Lee, J.; Hao, H.; Park, Y.D.; Zhan, Y.; Lü, Z.R. The effect of oxaloacetic acid on tyrosinase activity and structure: Integration of inhibition kinetics with docking simulation. Int. J. Biol. Macromol. 2017, 101, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, M.V. Signaling pathways in melanosome biogenesis and pathology. Int. J. Biochem. Cell Biol. 2010, 42, 1094–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.W.; Kim, J.H.; Song, H.; Seok, J.K.; Hong, S.S.; Boo, Y.C. Luteolin 7-Sulfate Attenuates Melanin Synthesis through Inhibition of CREB- and MITF-Mediated Tyrosinase Expression. Antioxidants 2019, 8, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hałdys, K.; Goldeman, W.; Jewgiński, M.; Wolińska, E.; Anger, N.; Rossowska, J.; Latajka, R. Inhibitory properties of aromatic thiosemicarbazones on mushroom tyrosinase: Synthesis, kinetic studies, molecular docking and effectiveness in melanogenesis inhibition. Bioorg. Chem. 2018, 81, 577–586. [Google Scholar] [CrossRef]
- Asanuma, M.; Miyazaki, I.; Ogawa, N. Dopamine- or L-DOPA-induced neurotoxicity: The role of dopamine quinone formation and tyrosinase in a model of Parkinson’s disease. Neurotox. Res. 2003, 5, 165–176. [Google Scholar] [CrossRef]
- El-Din, M.I.G.; Youssef, F.S.; Ashour, M.L.; Eldahshan, O.A.; Singab, A.N.B. New γ-pyrone glycoside from Pachira glabra and assessment of its gastroprotective activity using an alcohol-induced gastric ulcer model in rats. Food Funct. 2020, 11, 1958–1965. [Google Scholar] [CrossRef]
- Arung, E.T.; Shimizu, K.; Tanaka, H.; Kondo, R. 3-Prenyl luteolin, a new prenylated flavone with melanin biosynthesis inhibitory activity from wood of Artocarpus heterophyllus. Fitoterapia 2010, 81, 640–643. [Google Scholar] [CrossRef]
- Arroo, R.R.J.; Sari, S.; Barut, B.; Özel, A.; Ruparelia, K.C.; Şöhretoğlu, D. Flavones as tyrosinase inhibitors: Kinetic studies in vitro and in silico. Phytochem. Anal. PCA 2020, 31, 314–321. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [Green Version]
- Gholamhoseinian, A.; Razmi, Z. Screening the methanolic extracts of some plants for tyrosinase inhibitory activity. Toxicol. Environ. Chem. 2012, 94, 310–318. [Google Scholar] [CrossRef]
- El-Din, M.I.G.; Youssef, F.S.; Said, R.S.; Ashour, M.L.; Eldahshan, O.A.; Singab, A.N.B. Chemical constituents and gastro-protective potential of Pachira glabra leaves against ethanol-induced gastric ulcer in experimental rat model. Inflammopharmacology 2021, 29, 317–332. [Google Scholar] [CrossRef]
- Rupasinghe, H. Special Issue “flavonoids and their disease prevention and treatment potential”: Recent advances and future perspectives. Molecules 2020, 25, 4746. [Google Scholar] [CrossRef]
- Brunetti, C.; Di Ferdinando, M.; Fini, A.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants and developmental regulators: Relative significance in plants and humans. Int. J. Mol. Sci. 2013, 14, 3540–3555. [Google Scholar] [CrossRef] [Green Version]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Jacob, V.; Hagai, T.; Soliman, K. Structure-activity relationships of flavonoids. Curr. Org. Chem. 2011, 15, 2641–2657. [Google Scholar] [CrossRef]
- Lin, J.-W.; Chiang, H.-M.; Lin, Y.-C.; Wen, K.-C. Natural products with skin-whitening effects. J. Food Drug Anal. 2008, 16, 8. [Google Scholar] [CrossRef]
- Quan, N.V.; Thien, D.D.; Khanh, T.D.; Tran, H.D.; Xuan, T.D. Momilactones A, B, and Tricin in Rice Grain and By-Products are Potential Skin Aging Inhibitors. Foods 2019, 8, 602. [Google Scholar] [CrossRef] [Green Version]
- El-Nashar, H.A.; Eldehna, W.M.; Al-Rashood, S.T.; Alharbi, A.; Eskandrani, R.O.; Aly, S.H. GC/MS Analysis of Essential Oil and Enzyme Inhibitory Activities of Syzygium cumini (Pamposia) Grown in Egypt: Chemical Characterization and Molecular Docking Studies. Molecules 2021, 26, 6984. [Google Scholar] [CrossRef]
- Abdelghffar, E.A.; El-Nashar, H.A.; Al-Mohammadi, A.G.; Eldahshan, O.A. Orange fruit (Citrus sinensis) peel extract attenuates chemotherapy-induced toxicity in male rats. Food Funct. 2021, 12, 9443–9455. [Google Scholar] [CrossRef]
- Wang, N.; Hebert, D.N. Tyrosinase maturation through the mammalian secretory pathway: Bringing color to life. Pigment Cell Res. 2006, 19, 3–18. [Google Scholar] [CrossRef]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Seo, S.Y.; Sharma, V.K.; Sharma, N. Mushroom tyrosinase: Recent prospects. J. Agric. Food Chem. 2003, 51, 2837–2853. [Google Scholar] [CrossRef]
- Halaouli, S.; Record, E.; Casalot, L.; Hamdi, M.; Sigoillot, J.C.; Asther, M.; Lomascolo, A. Cloning and characterization of a tyrosinase gene from the white-rot fungus Pycnoporus sanguineus, and overproduction of the recombinant protein in Aspergillus niger. Appl. Microbiol. Biotechnol. 2006, 70, 580–589. [Google Scholar] [CrossRef]
- Aroca, P.; Garcia-Borron, J.C.; Solano, F.; Lozano, J.A. Regulation of mammalian melanogenesis. I: Partial purification and characterization of a dopachrome converting factor: Dopachrome tautomerase. Biochim. Et Biophys. Acta 1990, 1035, 266–275. [Google Scholar] [CrossRef]
- Elkhawas, Y.A.; Elissawy, A.M.; Elnaggar, M.S.; Mostafa, N.M.; Al-Sayed, E.; Bishr, M.M.; Singab, A.N.B.; Salama, O.M. Chemical diversity in species belonging to soft coral genus Sacrophyton and its impact on biological activity: A review. Mar. Drugs 2020, 18, 41. [Google Scholar] [CrossRef] [Green Version]
- Hirobe, T. Role of keratinocyte-derived factors involved in regulating the proliferation and differentiation of mammalian epidermal melanocytes. Pigment Cell Res. 2005, 18, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enerstvedt, K.H.; Jordheim, M.; Andersen, Ø.M. Isolation and identification of flavonoids found in Zostera marina collected in Norwegian coastal waters. Anal. Polyphen. Content Mar. Aquat. Angiosperms Nor. Coast. Waters 2016, 7, 1163–1172. [Google Scholar] [CrossRef] [Green Version]
- Santi, M.D.; Bouzidi, C.; Gorod, N.S.; Puiatti, M.; Michel, S.; Grougnet, R.; Ortega, M.G. In vitro biological evaluation and molecular docking studies of natural and semisynthetic flavones from Gardenia oudiepe (Rubiaceae) as tyrosinase inhibitors. Bioorg. Chem. 2019, 82, 241–245. [Google Scholar] [CrossRef]
- Apaza Ticona, L.; Thiebaut Estrada, C.; Rumbero Sánchez, Á. Inhibition of melanin production and tyrosinase activity by flavonoids isolated from Loranthus acutifolius. Nat. Prod. Res. 2020, 35, 4690–4693. [Google Scholar] [CrossRef]
- Yin, X.S.; Zhang, X.Q.; Yin, J.T.; Kong, D.Z.; Li, D.Q. Screening and identification of potential tyrosinase inhibitors from Semen Oroxyli extract by ultrafiltration LC-MS and in silico molecular docking. J. Chromatogr. Sci. 2019, 57, 838–846. [Google Scholar] [CrossRef]
- Lee, H.J.; Sim, M.O.; Woo, K.W.; Jeong, D.E.; Jung, H.K.; An, B.; Cho, H.W. Antioxidant and Antimelanogenic Activities of Compounds Isolated from the Aerial Parts of Achillea alpina L. Chem. Biodivers. 2019, 16, e1900033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Hu, X.; Zhang, B.; Wang, Z.; Hao, C.; Xin, J.; Guo, Q. Whitening Activity of Constituents Isolated from the Trichosanthes Pulp. Evid. Based Complement. Alternat. Med. 2020, 2020, 2582579. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Hou, J.; Xiong, J.; Li, M. Characterization of sulfuretin as a depigmenting agent. Fundam. Clin. Pharmacol. 2019, 33, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.X.; Nguyen, N.T.; Nguyen, M.H.K.; Le, T.H.; Van Do, T.N.; Hung, T.M.; Nguyen, M.T.T. Tyrosinase inhibitory activity of flavonoids from Artocarpus heterophyllous. Chem. Cent. J. 2016, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Bouzaiene, N.N.; Chaabane, F.; Sassi, A.; Chekir-Ghedira, L.; Ghedira, K. Effect of apigenin-7-glucoside, genkwanin and naringenin on tyrosinase activity and melanin synthesis in B16F10 melanoma cells. Life Sci. 2016, 144, 80–85. [Google Scholar] [CrossRef]
- Lee, B.; Moon, K.M.; Lee, B.-S.; Yang, J.-H.; Park, K.I.; Cho, W.-K.; Ma, J.Y. Swertiajaponin inhibits skin pigmentation by dual mechanisms to suppress tyrosinase. Oncotarget 2017, 8, 95530. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhao, X.; Tao, G.-J.; Chen, J.; Zheng, Z.-P. Investigating the inhibitory activity and mechanism differences between norartocarpetin and luteolin for tyrosinase: A combinatory kinetic study and computational simulation analysis. Food Chem. 2017, 223, 40–48. [Google Scholar] [CrossRef]
- Park, S.-A.; Jegal, J.; Chung, K.W.; Jung, H.J.; Noh, S.G.; Chung, H.Y.; Ahn, J.; Kim, J.; Yang, M.H. Isolation of tyrosinase and melanogenesis inhibitory flavonoids from Juniperus chinensis fruits. Biosci. Biotechnol. Biochem. 2018, 82, 2041–2048. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Shan, Y.; Yang, Z.; Zhang, L.; Ling, W.; Liang, Y.; Ouyang, Z.; Zhong, B.; Zhang, J. Chemical composition, antioxidant, antibacterial, and tyrosinase inhibition activity of extracts from Newhall navel orange (Citrus sinensis Osbeck cv. Newhall). Peel. J. Sci. Food Agric. 2020, 100, 2664–2674. [Google Scholar] [CrossRef]
- Ren, G.; Xue, P.; Sun, X.; Zhao, G. Determination of the volatile and polyphenol constituents and the antimicrobial, antioxidant, and tyrosinase inhibitory activities of the bioactive compounds from the by-product of Rosa rugosa Thunb. var. plena Regal Tea. BMC Complement. Altern. Med. 2018, 18, 307. [Google Scholar] [CrossRef] [Green Version]
- Kishore, N.; Twilley, D.; Blom van Staden, A.; Verma, P.; Singh, B.; Cardinali, G.; Kovacs, D.; Picardo, M.; Kumar, V.; Lall, N. Isolation of Flavonoids and Flavonoid Glycosides from Myrsine africana and Their Inhibitory Activities against Mushroom Tyrosinase. J. Nat. Prod. 2018, 81, 49–56. [Google Scholar] [CrossRef]
- Lee, S.-G.; Karadeniz, F.; Seo, Y.; Kong, C.-S. Anti-melanogenic effects of flavonoid glycosides from Limonium tetragonum (Thunb.) Bullock via inhibition of tyrosinase and tyrosinase-related proteins. Molecules 2017, 22, 1480. [Google Scholar] [CrossRef] [Green Version]
- Şöhretoğlu, D.; Sari, S.; Barut, B.; Özel, A. Tyrosinase inhibition by some flavonoids: Inhibitory activity, mechanism by in vitro and in silico studies. Bioorg. Chem. 2018, 81, 168–174. [Google Scholar] [CrossRef]
- Tadrent, W.; Alabdul Magid, A.; Kabouche, A.; Harakat, D.; Voutquenne-Nazabadioko, L.; Kabouche, Z. A new sulfonylated flavonoid and other bioactive compounds isolated from the aerial parts of Cotula anthemoides L. Nat. Prod. Res. 2017, 31, 1437–1445. [Google Scholar] [CrossRef]
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef]
- Choi, Y.-H.; Lee, J.Y.; Lee, J.E.; Jung, Y.W.; Jeong, W.; Hong, S.S.; Cho, Y.-R.; Choi, C.W. Skin-Related Properties and Constituents from the Aerial Parts Extract of Persicaria senticosa. Oxid Med Cell Longev 2020, 2020, 6627752. [Google Scholar] [CrossRef]
- Zengin, G.; Stefanucci, A.; Rodrigues, M.J.; Mollica, A.; Custodio, L.; Aumeeruddy, M.Z.; Mahomoodally, M.F. Scrophularia lucida L. as a valuable source of bioactive compounds for pharmaceutical applications: In vitro antioxidant, anti-inflammatory, enzyme inhibitory properties, in silico studies, and HPLC profiles. J. Pharm. Biomed. Anal. 2019, 162, 225–233. [Google Scholar] [CrossRef]
- Odonbayar, B.; Murata, T.; Batkhuu, J.; Yasunaga, K.; Goto, R.; Sasaki, K. Antioxidant flavonols and phenolic compounds from Atraphaxis frutescens and their inhibitory activities against insect phenoloxidase and mushroom tyrosinase. J. Nat. Prod. 2016, 79, 3065–3071. [Google Scholar] [CrossRef]
- Chen, J.; Yu, X.; Huang, Y. Inhibitory mechanisms of glabridin on tyrosinase. Spectrochimica acta. Part A Mol. Biomol. Spectrosc. 2016, 168, 111–117. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, M.M.; Sun, Y.; Wang, L.F.; Wang, H.; Zhang, Y.J.; Li, H.Y.; Zhuang, P.W.; Yang, Z. Synergistic Promotion on Tyrosinase Inhibition by Antioxidants. Molecules 2018, 23, 106. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.E.; Chang, B.Y.; Ham, S.O.; Kim, Y.C.; Kim, S.Y. Neobavaisoflavone Inhibits Melanogenesis through the Regulation of Akt/GSK-3β and MEK/ERK Pathways in B16F10 Cells and a Reconstructed Human 3D Skin Model. Molecules 2020, 25, 2683. [Google Scholar] [CrossRef]
- Promden, W.; Viriyabancha, W.; Monthakantirat, O.; Umehara, K.; Noguchi, H.; De-Eknamkul, W. Correlation between the Potency of Flavonoids on Mushroom Tyrosinase Inhibitory Activity and Melanin Synthesis in Melanocytes. Molecules 2018, 23, 1403. [Google Scholar] [CrossRef] [Green Version]
- Qu, L.; Song, K.; Zhang, Q.; Guo, J.; Huang, J. Simultaneous Determination of Six Isoflavones from Puerariae Lobatae Radix by CPE-HPLC and Effect of Puerarin on Tyrosinase Activity. Molecules 2020, 25, 344. [Google Scholar] [CrossRef] [Green Version]
- Wagle, A.; Seong, S.H.; Jung, H.A.; Choi, J.S. Identifying an isoflavone from the root of Pueraria lobata as a potent tyrosinase inhibitor. Food Chem. 2019, 276, 383–389. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, H.Y.; Kang, S.Y.; Kim, J.-B.; Kim, Y.H.; Jin, C.H. Chemical Constituents from Apios americana and Their Inhibitory Activity on Tyrosinase. Molecules 2018, 23, 232. [Google Scholar] [CrossRef] [Green Version]
- Abenavoli, L.; Capasso, R.; Milic, N.; Capasso, F. Milk thistle in liver diseases: Past, present, future. Phytother. Res. PTR 2010, 24, 1423–1432. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, J.Y.; Jenis, J.; Li, Z.P.; Ban, Y.J.; Baiseitova, A.; Park, K.H. Tyrosinase inhibitory study of flavonolignans from the seeds of Silybum marianum (Milk thistle). Bioorg. Med. Chem. 2019, 27, 2499–2507. [Google Scholar] [CrossRef]
- Chunhakant, S.; Chaicharoenpong, C. Antityrosinase, Antioxidant, and Cytotoxic Activities of Phytochemical Constituents from Manilkara zapota L. Bark. Molecules 2019, 24, 2798. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Zhang, G.; Pan, J.; Gong, D. An inhibition mechanism of dihydromyricetin on tyrosinase and the joint effects of vitamins B(6), D(3) or E. Food Funct. 2017, 8, 2601–2610. [Google Scholar] [CrossRef]
- Sari, S.; Barut, B.; Özel, A.; Kuruüzüm-Uz, A.; Şöhretoğlu, D. Tyrosinase and α-glucosidase inhibitory potential of compounds isolated from Quercus coccifera bark: In vitro and in silico perspectives. Bioorg. Chem. 2019, 86, 296–304. [Google Scholar] [CrossRef]
- Li, H.X.; Park, J.U.; Su, X.D.; Kim, K.T.; Kang, J.S.; Kim, Y.R.; Kim, Y.H.; Yang, S.Y. Identification of Anti-Melanogenesis Constituents from Morus alba L. Leaves. Molecules 2018, 23, 2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imen, M.B.; Chaabane, F.; Nadia, M.; Soumaya, K.J.; Kamel, G.; Leila, C.G. Anti-melanogenesis and antigenotoxic activities of eriodictyol in murine melanoma (B16-F10) and primary human keratinocyte cells. Life Sci. 2016, 135, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Salah, N.M.; Souleman, A.M.; Shaker, K.H.; Hawary, S.; El-Hady, F. Acetylcholinesterase, alpha-glucosidase and tyrosinase inhibitors from Egyptian propolis. Int. J. Pharm. Phytochem. Res. 2017, 9, 528–536. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, I.S.; So, Y.K.; Kim, H.-H.; Kim, Y.H. Kushenol A and 8-prenylkaempferol, tyrosinase inhibitors, derived from Sophora flavescens. J. Enzym. Inhib. Med. Chem. 2018, 33, 1048–1054. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Mei, H.-C.; Kuo, I.C.; Lee, T.-H.; Chen, Y.-H.; Lee, C.-K. Characterizing Tyrosinase Modulators from the Roots of Angelica keiskei Using Tyrosinase Inhibition Assay and UPLC-MS/MS as the Combinatorial Novel Approach. Molecules 2019, 24, 3297. [Google Scholar] [CrossRef] [Green Version]
- Mohd Sakeh, N.; Md Razip, N.N.; Mohd Ma’in, F.I.; Abdul Bahari, M.N.; Latif, N.; Akhtar, M.N.; Balia Yusof, Z.N.; Ahmad, S. Melanogenic Inhibition and Toxicity Assessment of Flavokawain A and B on B16/F10 Melanoma Cells and Zebrafish (Danio rerio). Molecules 2020, 25, 3403. [Google Scholar] [CrossRef]
- Kim, D.W.; Woo, H.S.; Kim, J.Y.; Ryuk, J.A.; Park, K.H.; Ko, B.S. Phenols displaying tyrosinase inhibition from Humulus lupulus. J. Enzym. Inhib. Med. Chem. 2016, 31, 742–747. [Google Scholar] [CrossRef] [Green Version]
- Lall, N.; Mogapi, E.; De Canha, M.N.; Crampton, B.; Nqephe, M.; Hussein, A.A.; Kumar, V. Insights into tyrosinase inhibition by compounds isolated from Greyia radlkoferi Szyszyl using biological activity, molecular docking and gene expression analysis. Bioorg. Med. Chem. 2016, 24, 5953–5959. [Google Scholar] [CrossRef]
- Hu, X.; Yu, M.H.; Yan, G.R.; Wang, H.Y.; Hou, A.J.; Lei, C. Isoprenylated phenolic compounds with tyrosinase inhibition from Morus nigra. J. Asian Nat. Prod. Res. 2018, 20, 488–493. [Google Scholar] [CrossRef]
- Hu, S.; Zheng, Z.; Chen, F.; Wang, M. The depigmenting effect of natural resorcinol type polyphenols Kuwanon O and Sanggenon T from the roots of Morus australis. J. Ethnopharmacol. 2017, 195, 196–203. [Google Scholar] [CrossRef]
- Lim, J.W.; Ha, J.H.; Jeong, Y.J.; Park, S.N. Anti-melanogenesis effect of dehydroglyasperin C through the downregulation of MITF via the reduction of intracellular cAMP and acceleration of ERK activation in B16F1 melanoma cells. Pharmacol. Rep. PR 2018, 70, 930–935. [Google Scholar] [CrossRef]
- Lim, J.; Nam, S.; Jeong, J.H.; Kim, M.J.; Yang, Y.; Lee, M.-S.; Lee, H.G.; Ryu, J.-H.; Lim, J.-S. Kazinol U inhibits melanogenesis through the inhibition of tyrosinase-related proteins via AMP kinase activation. Br. J. Pharm. 2019, 176, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, S.A.; Jamil, S.; Basar, N.; Abdul Lathiff, S.M.; Mohd Arriffin, N. Flavonoids from the leaves and heartwoods of Artocarpus lowii King and their bioactivities. Nat. Prod. Res. 2017, 31, 1113–1120. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.Y.; Liu, X.; He, X.R.; Ren, G.; Feng, L.H.; Zhou, Z.W. Artopithecins A-D, Prenylated 2-Arylbenzofurans from the Twigs of Artocarpus pithecogallus and Their Tyrosinase Inhibitory Activities. Chem. Pharm. Bull. 2018, 66, 1199–1202. [Google Scholar] [CrossRef]
- Zar Wynn Myint, K.; Kido, T.; Kusakari, K.; Prasad Devkota, H.; Kawahara, T.; Watanabe, T. Rhusflavanone and mesuaferrone B: Tyrosinase and elastase inhibitory biflavonoids extracted from the stamens of Mesua ferrea L. Nat. Prod. Res. 2021, 35, 1024–1028. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Flavonoid | Source | Assay Type | IC50 or % Inhibition | Ref. | |
|---|---|---|---|---|---|
| Flavones | |||||
| 1. | Tricin (3′,5′-dimethoxy-4′,5,7-trihydroxyflavone) | Oryza sativa | In vitro | 15.69% | [17] |
| 2. | Luteolin-7-sulfate (3′,4′,5-trihydroxy-flavone-7-sulphate) | Phyllospadix iwatensis Zostera marina | In vitro | Dose-dependent | [18] |
| 3. | 2′,4′,6-trimethoxyflavone | Loranthus acutifolius | In vitro | 4.00 μM | [20] |
| 4. | 3′,4′,5-trihydroxy-6,7,8-trimethoxyflavone | 11.30 μM | |||
| 5. | 4′,5-dihydroxy-6,7,8-trimethoxyflavone | 8.60 μM | |||
| 6. | Baicalein (5,6,7-trihydroxyflavone) | Oroxylum indicum | In silico | 0.29 mM | [21] |
| 7. | Oroxin A (baicalin-7-glucoside) | Oroxylum indicum | In silico | 0.50 mM | [22] |
| 8. | Isovitexin (apigenin-6-C-glucoside) | Achillea alpina | In vitro | Dose-dependent | |
| 9. | Diosmetin (3′,5,7-trihydroxy-4′-methoxyflavone) | Trichosanthes kirilowii | In vitro | Dose-dependent | [23] |
| 10. | Chrysoeriol (4′,5,7-trihydroxy-3′-methoxyflavone) | ||||
| 11. | Scutellarin (4′,5,6,7-tetrahydroxy-flavone-7-β-d-glucopyranuronoside) | ||||
| 12. | 3′,5-dihydroxy-7-(β-d-glucopyranosyloxy)−4′-methoxyflavone | ||||
| 13. | Sulfuretin (6,3′,4′-trihydroxyaurone) | Rhus verniciflua | In vivo | 20 μM | [24] |
| 14. | artocaepin E | Artocarpus heterophyllous | In vitro | 6.7 μM | [25] |
| 15. | Genkwanin (4′,5-dihydroxy-7-methoxyflavone) | Daphne gnidium Alnus glutinosa Asplenium normale | In vitro | Dose-dependent | [26] |
| 16. | Apigenin-7-O-β-d-glucopyranoside | Thymus species | In vitro | Dose-dependent | [26] |
| 17. | Swertiajaponin ((6-C-β-d-glucopyranosyl-7-O-methylluteolin) | Swertia japonica, Cymbopogon citratus | Human skin model | 43.47 μM | [27] |
| 18. | Norartocarpetin (5,7,2′,4′-tetrahydroxyflavone) | Artocarpus dadah | In silico | 0.12 μM | [28] |
| 19. | Luteolin (3′,4′,5,7-tetrahydroxy-flavone) | Reseda luteola Elsholtzia rugulosa | 266.67 μM | ||
| 20. | Hypolaetin-7-O-β-d-glucopyranoside (8-Hydroxyluteolin-7-O-β-d-glucopyranoside) | Juniperus chinensis | In vitro In silico | 73.30% | [29] |
| Flavonols | |||||
| 21. | Swertiajaponin ((6-C-β-d-glucopyranosyl-7-O-methylluteolin) | Swertia japonica Cymbopogon citratus | Human skin model | Dose-dependent | [27] |
| 22. | 4′,5,6,7-tetramethoxyflavone | ||||
| 23. | Nobiletin | ||||
| 24. | 3,3′,4′,5,6,7-hexamethoxyflavone | ||||
| 25. | Hyperoside | Rosa rugosa | In vitro | IC50 ˂ 1 μg/mL | [32] |
| 26. | Kaempferol-3-O-rutinoside | ||||
| 27. | Rutin (quercetin 3-rutinoside) | Rosa rugose Myrsine africana | In vitro | 2.30 μM | [32,33] |
| 28. | Myricetin-3-O-α-l-rhamnopyranoside | Myrsine africana | In vitro In silico | 2.00 μM | [31,41] |
| 29. | Myricetin-3-O-β-galactopyranoside | Limonium tetragonum | In vitro | 65% | [34] |
| 30. | Quercetin-3-O-β-galactopyranoside | 63% | |||
| 31. | 5,7,4′,5′-tetrahydoxyflavonol 2′-[propanoic acid-(2″’-acetoxy-1″’-sulfonyl)]−5′-O-β-d-glucopyranoside | Cotula anthemoides | In vitro | 100 ± 0.5 μM | [36] |
| 32. | Quercetin | Persicaria senticosa | In silico In vitro | 3.08 ± 0.74 mol L−1 14.31 ± 3.93 μM | [37,38] |
| 33. | Quercetin-3-O-α-arabinopyranosyl-(1→6)-β-glucopyranoside | Scrophularia lucida | In silico | 46.94 ± 3.09 | [39] |
| 34. | Hesperidin (hesperetin 7-rutinoside) | Undetermined | |||
| 35. | 8-β-d-glucopyranosyloxy-3′,4′,5,5′-tetrahydroxy-7-methoxy-3-α-l-rhamnopyranosyloxyflavone | Atraphaxis frutescens | In vitro | 0.90 ± 0.02 mM | [40] |
| 36. | 3′,4′,5,5′,8-pentahydroxy-7-methoxy-3-α-l-rhamnopyranosyloxyflavone | 1.20 ± 0.03 mM | |||
| 37. | Europetin 3-O-α-l-rhamnopyranoside | 1.10 ± 0.07 mM | |||
| 38. | Myricetin-3-O-α-l-rhamnopyranoside | Myrsine africana | In vitro In silico | 0.12 ± 0.002 mM | [41] |
| Isoflavones | |||||
| 39. | Glabridin | Glycyrrhiza glabra | In silico | 0.43 μmol/L | [41] |
| 40. | Neobavaisoflavone | Pueraria 1lobate | In vitro Human skin model | 10–45% | [42,43] |
| 41. | (6aR,11aR)-3,8-dihydroxy-9-methoxy pterocarpan | Dalbergia parviflora | In vitro | 84.60% | [45] |
| 42. | Puerarin (daidzein-8-C-glucoside) | Pueraria lobata | In vitro | 0.537 mg/mL | [46] |
| 43. | Calycosin (3′,7-dihydroxy-4′-methoxyisoflavone) | In vitro In silico | 85.60% | [47] | |
| 44. | Lupinalbin A | Apios americana | In vitro In silico | 39.70 ± 1.5 µg/mL | [48] |
| 45. | 2′-hydroxygenistein-7-O-gentibioside | 50.00 ± 3.70 µg/mL | |||
| Flavan-3-ols | |||||
| 46. | Silybin | Silybum marianum | In vitro | 1.70 ± 0.07 µM | [50] |
| 47. | (+)-dihydrokaempferol | Manilkara zapota | In vitro | 45.35 ± 0.60 µM | [51] |
| 48. | Dihydromyricetin | In silico | 36.6 ± 0.14 µM | [52] | |
| 49. | (−)-8-chlorocatechin | Quercus coccifera | In silico | 4.05 ± 0.30 µg/mL | [53] |
| Flavanones | |||||
| 50. | Steppogenin (5,7,2′,4′-tetrahydroxyflavanone) | Artocarpus heterophyllous Morus alba | In vitro In silico | 7.50 ± 0.50 μM | [25,54] |
| 51. | Eriodictyol (3′,4′,5,7-tetrahydroxyflavanone) | Eriodictyon californicum | In vitro | Dose-dependent | [55] |
| 52. | Artocarpanone (2′,4′,5-trihydroxy-7-methoxy-Isoflavone) | Artocarpus heterophyllous | In vitro | 2.0 ± 0.1 μM | [25] |
| 53. | Liquiritigenin (4′,7-dihydroxyflavanone) | Artocarpus heterophyllous | In vitro | 22.00 ± 2.50 | [25] |
| 54. | Pinostrobin ((2s)-5-hydroxy-7-methoxyflavanone) | Egyptian Propolis | 36.30% | ||
| 55. | kushenol A | Sophora flavescens | In silico | 1.10 ± 0.70 μM | [57] |
| Chalcones | |||||
| 56. | 2′4′-dihydroxy-6′-methoxy-chalcone | Loranthus acutifolius | IC50 = 5.70 ± 0.02 μM | [56] | |
| 57. | Xanthoangelol (2′,4,4′-trihydroxy-3′-geranylchalcone) | Angelica keiskei | In vitro | 15.87 ± 1.21 μM | [58] |
| 58. | Flavokawain A (2′-hydroxy-4,4′,6′-trimethoxychalcone) | Piper methysticum | In vitro In vivo zebrafish | Dose-dependent | [59] |
| 59. | Flavokawain B (4′,6′-dimethoxy-2′-hydroxychalcone) | ||||
| 60. | Xanthohumol (2′,4,4′-trihydroxy-6′-methoxy-3′-prenylchalcone) | Humulus lupulus | In vitro | 15.40–22.1 μM | [60] |
| 61. | Xanthoumol B (dehydrocycloxanthohumol hydrate) | 41.10–46.70 μM | |||
| 62. | Xanthoumol C (dehydrocycloxanthohumol) | ||||
| 63. | 2′,4′,6′-trihydroxydihydrochalcone | Greyia radlkoferi | In vitro In silico | 17.70 μg/mL | [61] |
| Prenylated flavonoids | |||||
| 64. | Kuwanon J | Morus nigra | In vitro | 0.17 ± 0.01 µM | [62] |
| 65. | Sanggenon C | 1.17 ± 0.03 µM | |||
| 66. | Sanggenon M | 13.06 ± 0.58 µM | |||
| 67. | Sanggenon O | 1.15 ± 0.03 µM | |||
| 68. | Kuwanon O | Morus australis | In vitro Artificial skin models | 15-20% 8-12% | [63] |
| 69. | Sanggenon T | ||||
| 70. | Dehydroglyasperin C | Glycyrrhiza uralensis | In vitro | Dose-dependent | [64] |
| 71. | Kazinol U | Broussonetia kazinoki Sieb | In vitro In vivo zebrafish | Dose-dependent | [65] |
| 72. | Cycloheterophyllin | Artocarpus lowii | In vitro | 104.6 μM | [66] |
| 73. | Morachalcone A | Artocarpus pithecogallus | In vitro | 0.77 ± 0.01 μM | [67] |
| 74. | 6-prenylapigenin | 24.29 ± 0.12 μM | |||
| 75. | 6-prenylnaringenin | Humulus lupulus | In vitro | 38.1 µM | [60] |
| 76. | Isoxanthohumol | 77.4 µM | |||
| 77. | 8-prenylkaempferol | Sophora flavescents | In silico | 2.40 ± 1.10 μM | [57] |
| Biflavonoids | |||||
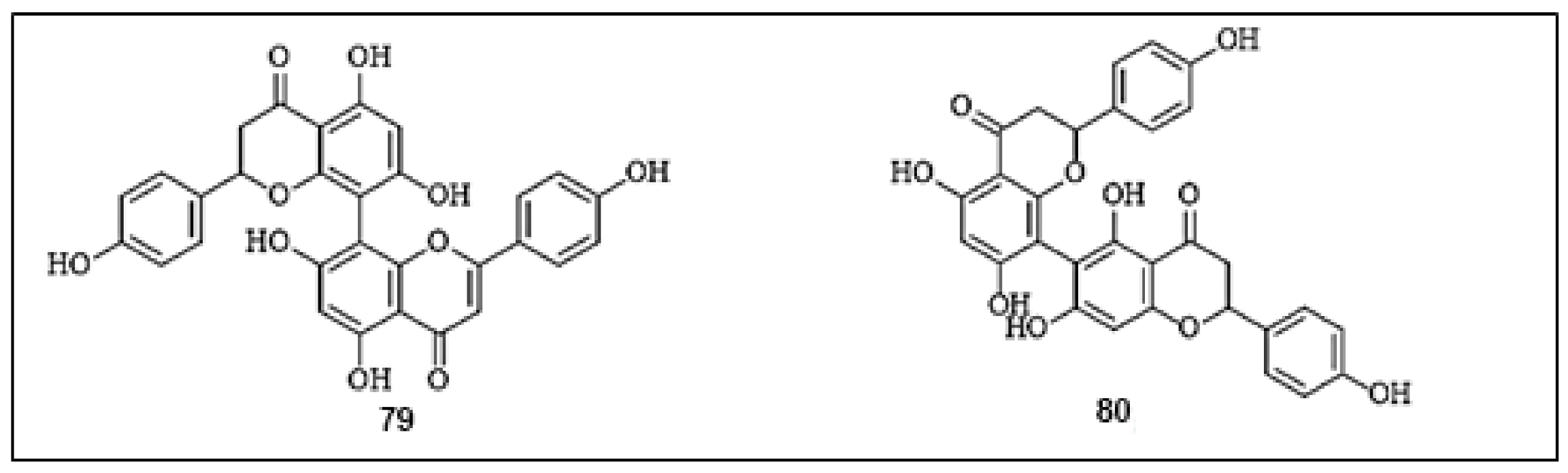
| 78. | Rhusflavanone | Mesua ferrea | In vitro | 10.60 µg/mL | [68] |
| 79. | mesuaferrone B | 10.30 µg/mL | |||
| Mode of Action | Compounds |
|---|---|
| Competitive inhibition of tyrosinase | 2′,4′,6-trimethoxyflavone (3) 3′,4′,5-trihydroxy-6,7,8-trimethoxyflavone (4) 4′,5-dihydroxy-6,7,8-trimethoxyflavone (5) Artocaepin E (14) Hypolaetin-7-O-β-d-glucopyranoside (20) Quercetin-7-O-α-l-rhamnopyranoside (21) Lupinalbin A (45) 2′-hydroxygenistein-7-O-gentibioside (46) (+)-dihydrokaempferol (48) (−)-8-chlorocatechin (50) Xanthohumol (61) Xanthoumol B (62) Xanthoumol C (63) 8-prenylkaempferol (78) |
| Non-competitive inhibition of tyrosinase | Tricin (1) Luteolin (11) Glabridin (40) |
| Mixedinhibition of tyrosinase | Puerarin (43) Silybin (47) Dihydromyricetin (49) 6-prenylnaringenin (76) |
| Downregulation of MITF expression | Luteolin-7-sulfate (2) Sulfuretin (13) Swertiajaponin (17) Neobavaisoflavone (41) Flavokawain A (59) Dehydroglyasperin C (71) Kazinol U (72) |
| Suppression of cAMP-CREB signaling pathway | Luteolin-7-sulfate (2) Steppogenin (51) Dehydroglyasperin C (71) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Nashar, H.A.S.; El-Din, M.I.G.; Hritcu, L.; Eldahshan, O.A. Insights on the Inhibitory Power of Flavonoids on Tyrosinase Activity: A Survey from 2016 to 2021. Molecules 2021, 26, 7546. https://doi.org/10.3390/molecules26247546
El-Nashar HAS, El-Din MIG, Hritcu L, Eldahshan OA. Insights on the Inhibitory Power of Flavonoids on Tyrosinase Activity: A Survey from 2016 to 2021. Molecules. 2021; 26(24):7546. https://doi.org/10.3390/molecules26247546
Chicago/Turabian StyleEl-Nashar, Heba A. S., Mariam I. Gamal El-Din, Lucian Hritcu, and Omayma A. Eldahshan. 2021. "Insights on the Inhibitory Power of Flavonoids on Tyrosinase Activity: A Survey from 2016 to 2021" Molecules 26, no. 24: 7546. https://doi.org/10.3390/molecules26247546
APA StyleEl-Nashar, H. A. S., El-Din, M. I. G., Hritcu, L., & Eldahshan, O. A. (2021). Insights on the Inhibitory Power of Flavonoids on Tyrosinase Activity: A Survey from 2016 to 2021. Molecules, 26(24), 7546. https://doi.org/10.3390/molecules26247546