
Effect of Plasma Activated Water on Selected Chemical Compounds of Rocket-Salad (Eruca sativa Mill.) Leaves

 ,
,  ,
,  ,
,  , ,
, ,  ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Results and Discussion
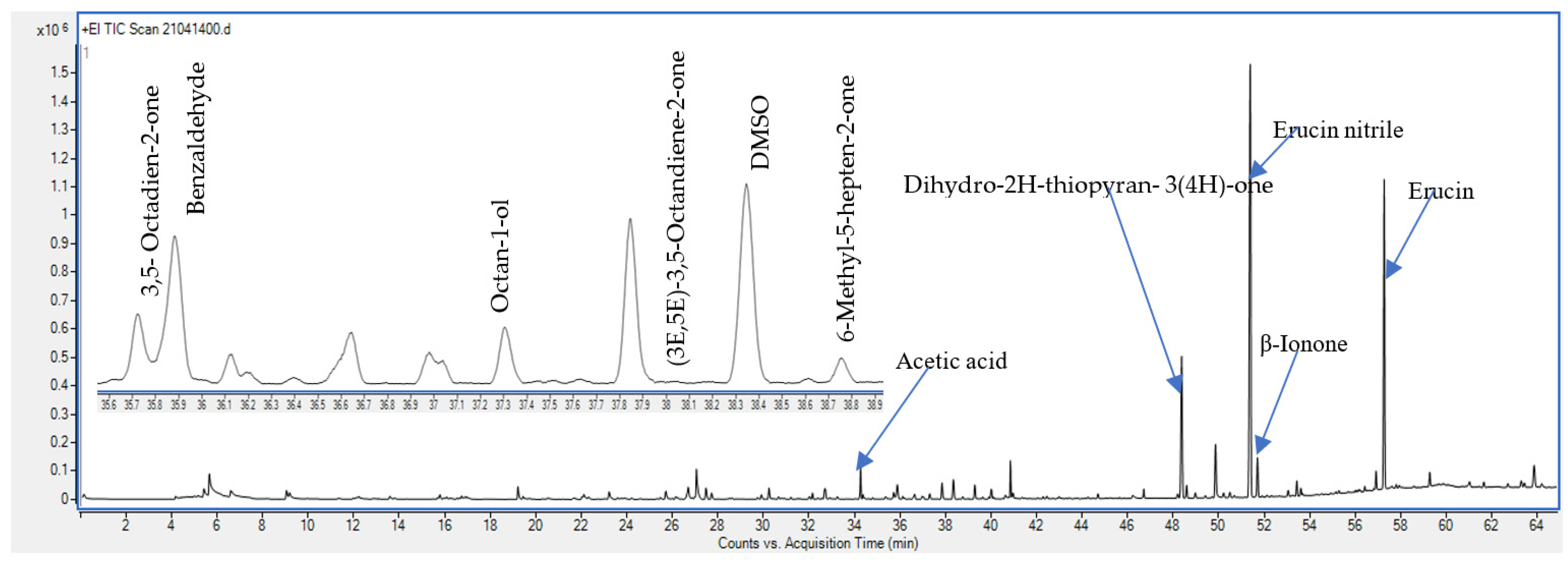
2.1. Effect of PAW Treatment on the Main VOCs in Rocket Leaves
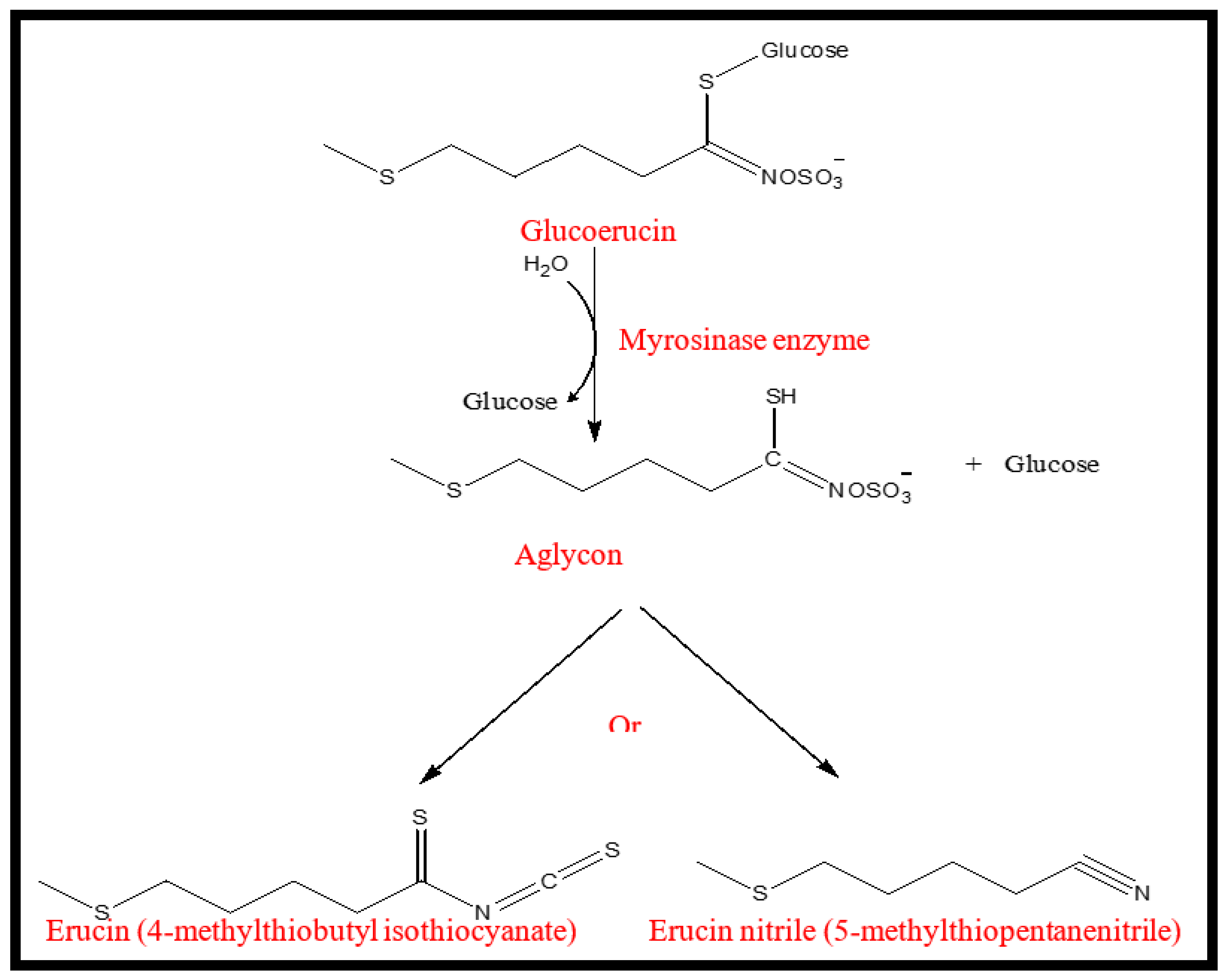
2.1.1. Glucosinolate Hydrolysis Products (GHPs)
2.1.2. Sulphur Compounds
2.1.3. Ketones
2.1.4. Aldehydes
2.1.5. Alcohols
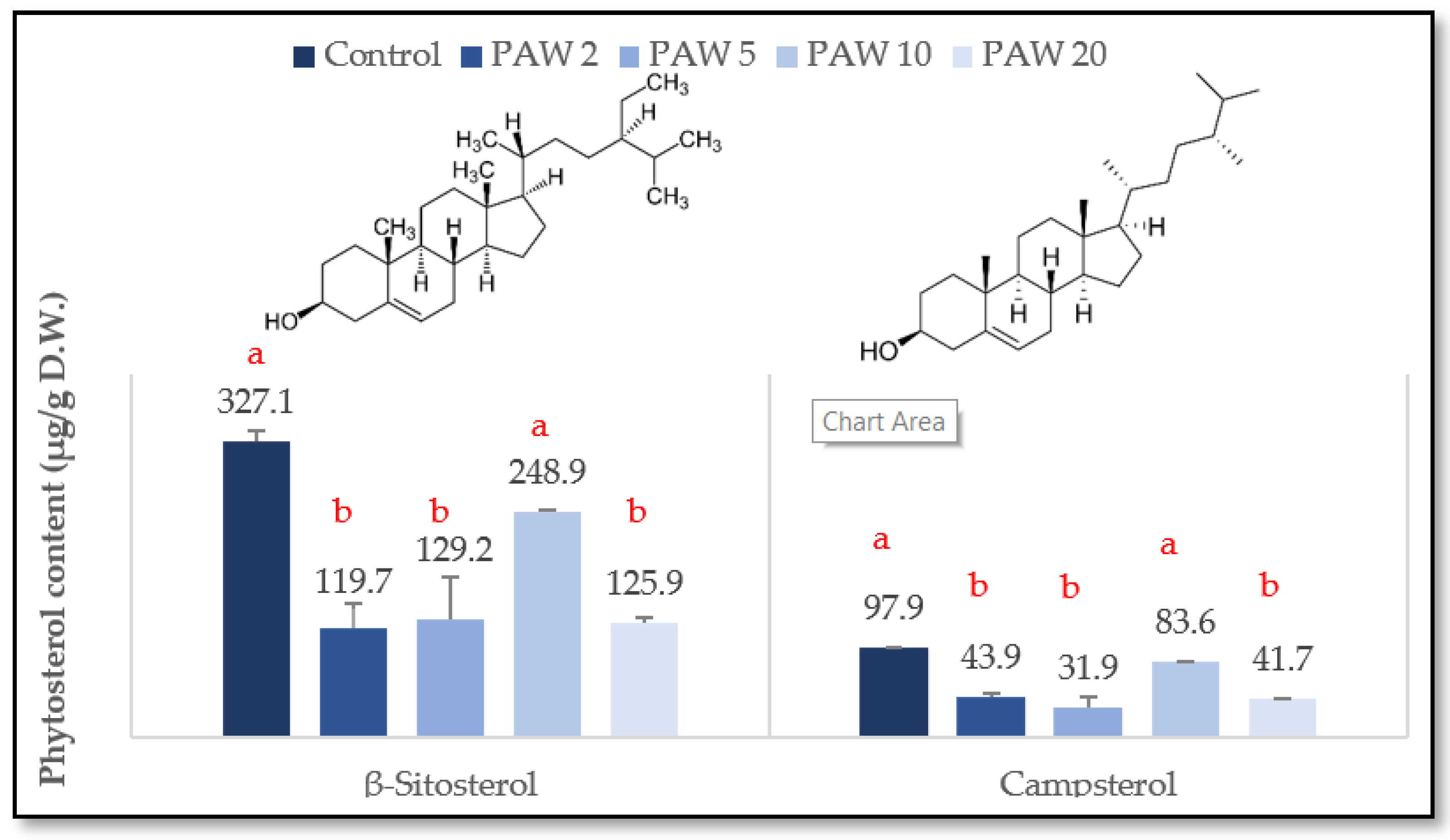
2.2. Effect of PAW Treatment on the Main Phytosterols in Rocket Leaves
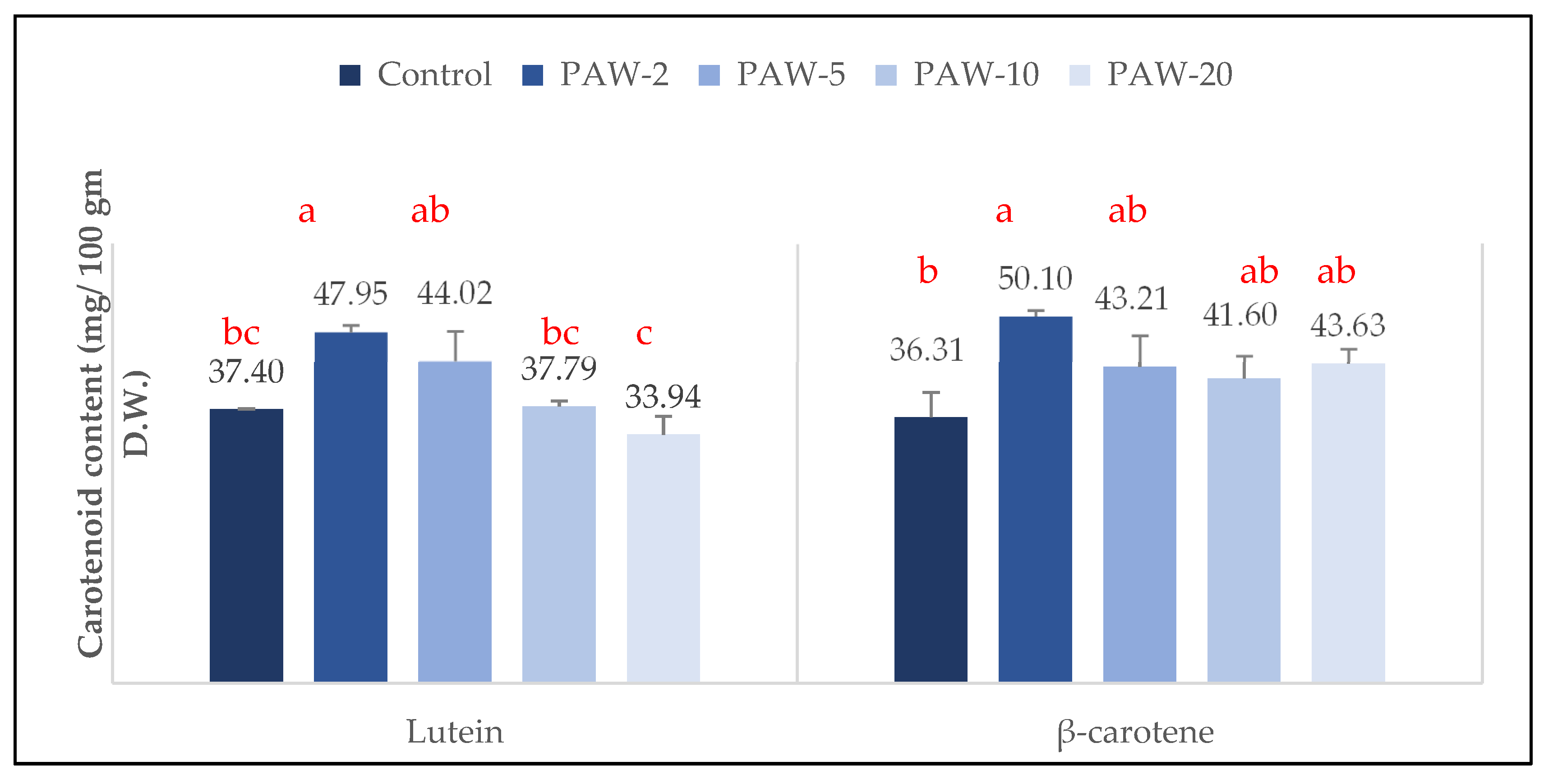
2.3. β-Carotene and Lutein Contents
2.4. Chlorophylls (Chl) Content
3. Materials and Methods
3.1. Raw Materials
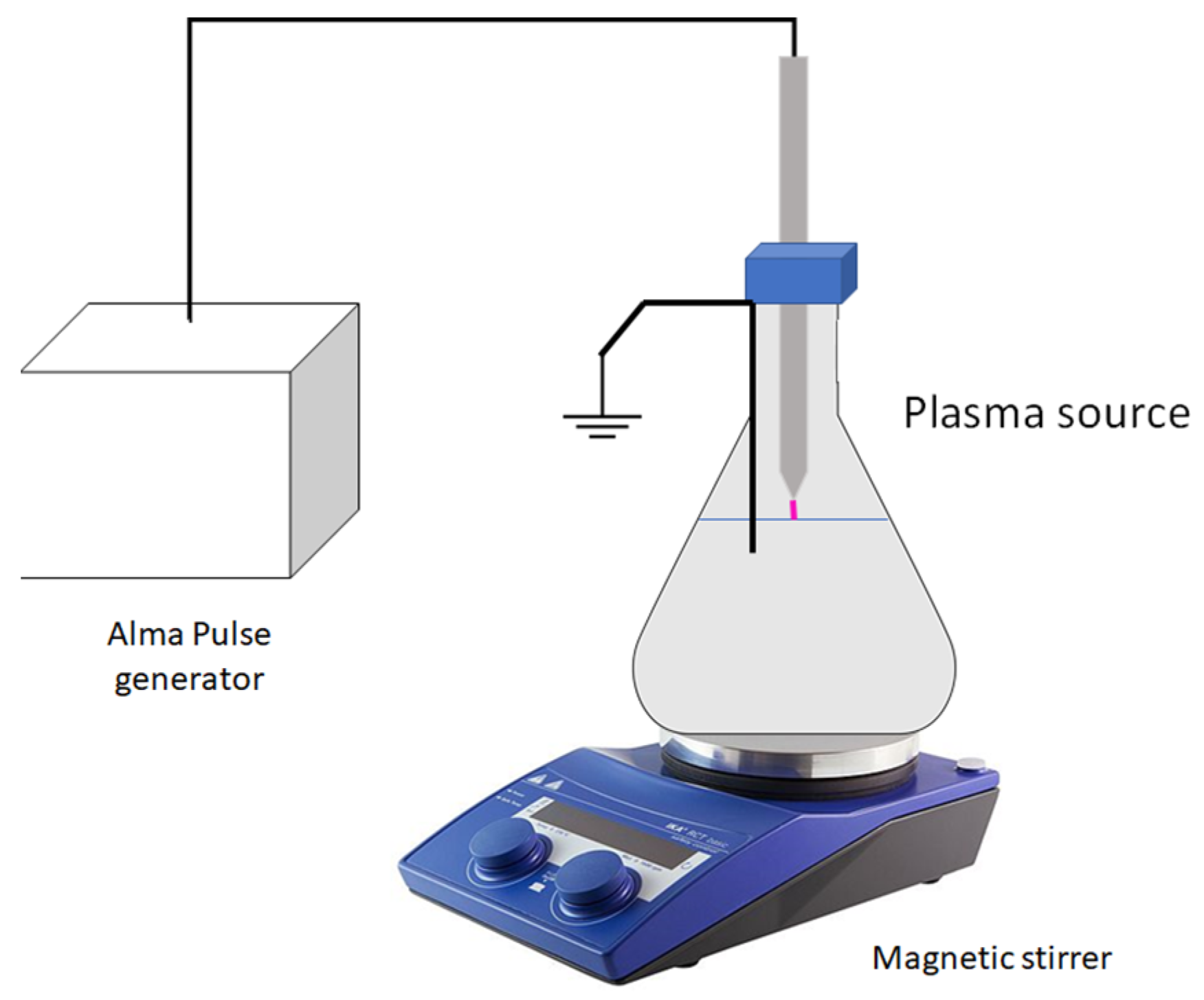
3.2. PAW Generation
3.3. PAW Treatment
3.4. Volatile Organic Compounds (VOCs) Analysis
3.4.1. Headspace Solid-Phase Microextraction (Hs-Spme)
3.4.2. Gc–Ms Analysis
3.5. Phytosterol Analysis by HPLC–DAD
3.5.1. Extraction of Phytosterols from Rocket Samples
3.5.2. HPLC–DAD Analyses
3.6. Determination of β-Carotene and Lutein Contents by HPLC–DAD
3.7. Chlorophyll Content
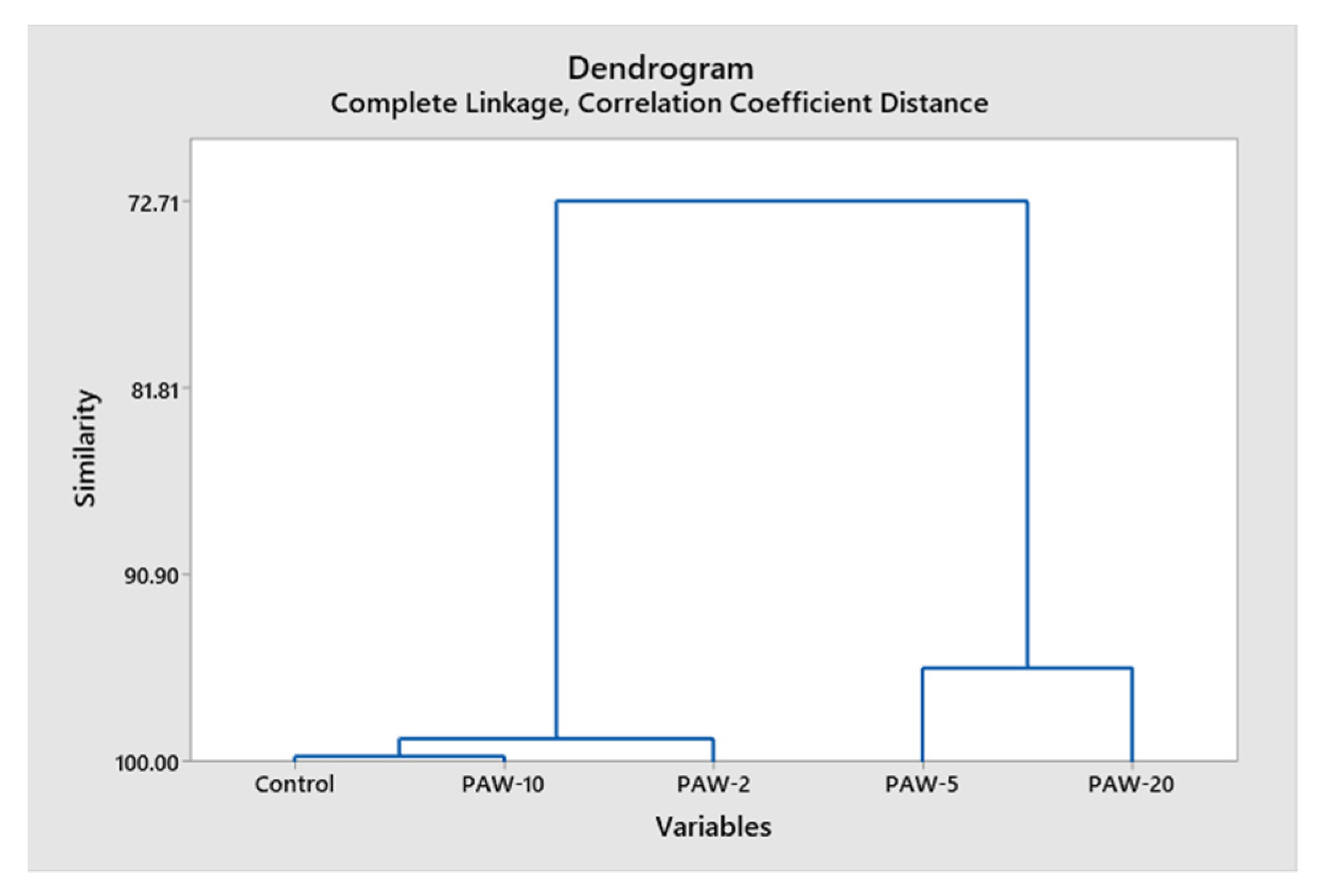
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bell, L.; Spadafora, N.D.; Müller, C.T.; Wagstaff, C.; Rogers, H.J. Use of TD-GC–TOF-MS to assess volatile composition during post-harvest storage in seven accessions of rocket salad (Eruca sativa). Food Chem. 2016, 194, 626–636. [Google Scholar] [CrossRef] [Green Version]
- Bell, L.; Wagstaff, C. Rocket science: A review of phytochemical & health-related research in Eruca & Diplotaxis species. Food Chem. X 2019, 1, 100002. [Google Scholar]
- Azarenko, O.; Jordan, M.A.; Wilson, L. Erucin, the major isothiocyanate in arugula (Eruca sativa), inhibits proliferation of MCF7 tumor cells by suppressing microtubule dynamics. PLoS ONE 2014, 9, e100599. [Google Scholar] [CrossRef] [Green Version]
- Jonvik, K.L.; Nyakayiru, J.; Pinckaers, P.J.; Senden, J.M.; Van Loon, L.J.; Verdijk, L.B. Nitrate-rich vegetables increase plasma nitrate and nitrite concentrations and lower blood pressure in healthy adults. J. Nutr. 2016, 146, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Blažević, I.; Mastelić, J. Free and bound volatiles of rocket (Eruca sativa Mill.). Flavour Fragr. J. 2008, 23, 278–285. [Google Scholar] [CrossRef]
- Khoobchandani, M.; Ganesh, N.; Gabbanini, S.; Valgimigli, L.; Srivastava, M. Phytochemical potential of Eruca sativa for inhibition of melanoma tumor growth. Fitoterapia 2011, 82, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Wagstaff, C. Glucosinolates, myrosinase hydrolysis products, and flavonols found in rocket (Eruca sativa and Diplotaxis tenuifolia). J. Agric. Food Chem. 2014, 62, 4481–4492. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Sánchez, A.; Allende, A.; Bennett, R.N.; Ferreres, F.; Gil, M.I. Microbial, nutritional and sensory quality of rocket leaves as affected by different sanitizers. Postharvest Biol. Technol. 2006, 42, 86–97. [Google Scholar] [CrossRef]
- Spadafora, N.D.; Amaro, A.L.; Pereira, M.J.; Müller, C.T.; Pintado, M.; Rogers, H.J. Multi-trait analysis of post-harvest storage in rocket salad (Diplotaxis tenuifolia) links sensorial, volatile and nutritional data. Food Chem. 2016, 211, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Nunes, T.P.; Martins, C.G.; Faria, A.F.; Bíscola, V.; de Oliveira Souza, K.L.; Mercadante, A.Z.; Cordenunsi, B.R.; Landgraf, M. Changes in total ascorbic acid and carotenoids in minimally processed irradiated Arugula (Eruca sativa Mill) stored under refrigeration. Radiat. Phys. Chem. 2013, 90, 125–130. [Google Scholar] [CrossRef]
- Gutiérrez, D.R.; Lemos, L.; Rodríguez, S.d.C. Effect of UV-C and ozone on the bioactive compounds and antioxidant capacity of minimally processed rocket (Eruca Sativa Mill.). Int. J. New Technol. Res 2018, 4, 23–29. [Google Scholar] [CrossRef]
- de Castro, D.R.G.; Mar, J.M.; da Silva, L.S.; da Silva, K.A.; Sanches, E.A.; de Araújo Bezerra, J.; Rodrigues, S.; Fernandes, F.A.; Campelo, P.H. Dielectric barrier atmospheric cold plasma applied on camu-camu juice processing: Effect of the excitation frequency. Food Res. Int. 2020, 131, 109044. [Google Scholar] [CrossRef]
- Thirumdas, R.; Kothakota, A.; Annapure, U.; Siliveru, K.; Blundell, R.; Gatt, R.; Valdramidis, V.P. Plasma activated water (PAW): Chemistry, physico-chemical properties, applications in food and agriculture. Trends Food Sci. Technol. 2018, 77, 21–31. [Google Scholar] [CrossRef]
- Laurita, R.; Gozzi, G.; Tappi, S.; Capelli, F.; Bisag, A.; Laghi, G.; Gherardi, M.; Cellini, B.; Abouelenein, D.; Vittori, S. Effect of plasma activated water (PAW) on rocket leaves decontamination and nutritional value. Innov. Food Sci. Emerg. Technol. 2021, 102805. [Google Scholar] [CrossRef]
- Raffo, A.; Masci, M.; Moneta, E.; Nicoli, S.; Del Pulgar, J.S.; Paoletti, F. Characterization of volatiles and identification of odor-active compounds of rocket leaves. Food Chem. 2018, 240, 1161–1170. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.; Methven, L.; Signore, A.; Oruna-Concha, M.J.; Wagstaff, C. Analysis of seven salad rocket (Eruca sativa) accessions: The relationships between sensory attributes and volatile and non-volatile compounds. Food Chem. 2017, 218, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Bell, L.; Kitsopanou, E.; Oloyede, O.O.; Lignou, S. Important Odorants of Four Brassicaceae Species, and Discrepancies between Glucosinolate Profiles and Observed Hydrolysis Products. Foods 2021, 10, 1055. [Google Scholar] [CrossRef]
- Mullaney, J.A.; Kelly, W.J.; McGhie, T.K.; Ansell, J.; Heyes, J.A. Lactic acid bacteria convert glucosinolates to nitriles efficiently yet differently from Enterobacteriaceae. J. Agric. Food Chem. 2013, 61, 3039–3046. [Google Scholar] [CrossRef]
- Jirovetz, L.; Smith, D.; Buchbauer, G. Aroma compound analysis of Eruca sativa (Brassicaceae) SPME headspace leaf samples using GC, GC−MS, and olfactometry. J. Agric. Food Chem. 2002, 50, 4643–4646. [Google Scholar] [CrossRef] [PubMed]
- Leng, C.; Zhang, Y.; Wang, M.; Wang, P.; Gu, Z.; Yang, R. Dynamic variation of glucosinolates and isothiocyanates in broccoli sprouts during hydrolysis. Sci. Hortic. 2019, 255, 128–133. [Google Scholar] [CrossRef]
- Melchini, A.; Traka, M.H. Biological profile of erucin: A new promising anticancer agent from cruciferous vegetables. Toxins 2010, 2, 593–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughn, S.F.; Berhow, M.A. Glucosinolate hydrolysis products from various plant sources: pH effects, isolation, and purification. Ind. Crop. Prod. 2005, 21, 193–202. [Google Scholar] [CrossRef]
- Bell, L.; Yahya, H.N.; Oloyede, O.O.; Methven, L.; Wagstaff, C. Changes in rocket salad phytochemicals within the commercial supply chain: Glucosinolates, isothiocyanates, amino acids and bacterial load increase significantly after processing. Food Chem. 2017, 221, 521–534. [Google Scholar] [CrossRef]
- Saengha, W.; Karirat, T.; Buranrat, B.; Matra, K.; Deeseenthum, S.; Katisart, T.; Luang-In, V. Cold Plasma Treatment on Mustard Green Seeds and its Effect on Growth, Isothiocyanates, Antioxidant Activity and Anticancer Activity of Microgreens. Int. J. Agric. Biol. 2021, 25, 667–676. [Google Scholar] [CrossRef]
- Bußler, S. Cold Atmospheric Pressure Plasma Treatment of Food Matrices. Ph.D. Thesis, Technische Universität Berlin, Berlin, Germany, 2017. [Google Scholar]
- Taviano, M.F.; Cavò, E.; Spadaro, V.; Raimondo, F.M.; Musolino, V.; Cacciola, F.; El Majdoub, Y.O.; Mondello, L.; Condurso, C.; Cincotta, F. Phytochemical Constituents, Antioxidant Activity, and Toxicity Assessment of the Aerial Part Extracts from the Infraspecific Taxa of Matthiola fruticulosa (Brassicaceae) Endemic to Sicily. Molecules 2021, 26, 4114. [Google Scholar] [CrossRef]
- Oulad El Majdoub, Y.; Alibrando, F.; Cacciola, F.; Arena, K.; Pagnotta, E.; Matteo, R.; Micalizzi, G.; Dugo, L.; Dugo, P.; Mondello, L. Chemical Characterization of Three Accessions of Brassica juncea L. Extracts from Different Plant Tissues. Molecules 2020, 25, 5421. [Google Scholar] [CrossRef]
- Korachi, M.; Ozen, F.; Aslan, N.; Vannini, L.; Guerzoni, M.E.; Gottardi, D.; Ekinci, F.Y. Biochemical changes to milk following treatment by a novel, cold atmospheric plasma system. Int. Dairy J. 2015, 42, 64–69. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, H.; Luo, J.; Liu, J.; Zhao, S.; Hu, Q.; Ding, C. Effect of dielectric barrier discharge cold plasma treatments on flavor fingerprints of brown rice. Food Chem. 2021, 352, 129402. [Google Scholar] [CrossRef]
- Pérez-Andrés, J.M.; Cropotova, J.; Harrison, S.M.; Brunton, N.P.; Cullen, P.J.; Rustad, T.; Tiwari, B.K. Effect of Cold Plasma on Meat Cholesterol and Lipid Oxidation. Foods 2020, 9, 1786. [Google Scholar] [CrossRef]
- Silveira, M.R.; Coutinho, N.M.; Esmerino, E.A.; Moraes, J.; Fernandes, L.M.; Pimentel, T.C.; Freitas, M.Q.; Silva, M.C.; Raices, R.S.; Ranadheera, C.S. Guava-flavored whey beverage processed by cold plasma technology: Bioactive compounds, fatty acid profile and volatile compounds. Food Chem. 2019, 279, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, A.; Comporti, M.; Fulceri, R.; Esterbauer, H. Cytotoxic aldehydes originating from the peroxidation of liver microsomal lipids: Identification of 4, 5-dihydroxydecenal. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1984, 792, 172–181. [Google Scholar] [CrossRef]
- Mead, J.F. Free radical mechanisms of lipid damage and consequences for cellular membranes. Eur. J. Immunol. 1976, 9, 426–433. [Google Scholar]
- Liu, J.; Liu, M.; He, C.; Song, H.; Guo, J.; Wang, Y.; Yang, H.; Su, X. A comparative study of aroma–active compounds between dark and milk chocolate: Relationship to sensory perception. J. Sci. Food Agric. 2015, 95, 1362–1372. [Google Scholar] [CrossRef] [PubMed]
- Ruther, J.; Kleier, S. Plant–plant signaling: Ethylene synergizes volatile emission in Zea mays induced by exposure to (Z)-3-hexen-1-ol. J. Chem. Ecol. 2005, 31, 2217–2222. [Google Scholar] [CrossRef]
- Hu, C.-J.; Li, D.; Ma, Y.-X.; Zhang, W.; Lin, C.; Zheng, X.-Q.; Liang, Y.-R.; Lu, J.-L. Formation mechanism of the oolong tea characteristic aroma during bruising and withering treatment. Food Chem. 2018, 269, 202–211. [Google Scholar] [CrossRef]
- Tawfik, M.M.; Yamato, K.T.; Kohchi, T.; Koeduka, T.; Matsui, K. n-Hexanal and (Z)-3-hexenal are generated from arachidonic acid and linolenic acid by a lipoxygenase in Marchantia polymorpha L. Biosci. Biotechnol. Biochem. 2017, 81, 1148–1155. [Google Scholar] [CrossRef] [Green Version]
- Campelo, P.H.; Alves Filho, E.G.; Silva, L.M.; de Brito, E.S.; Rodrigues, S.; Fernandes, F.A. Modulation of aroma and flavor using glow discharge plasma technology. Innov. Food Sci. Emerg. Technol. 2020, 62, 102363. [Google Scholar] [CrossRef]
- da Silva Marineli, R.; Furlan, C.P.B.; Marques, A.Y.C.; Bicas, J.; Pastore, G.M.; Maróstica, M.R., Jr. Phytosterols: Biological effects and mechanisms of hypocholesterolemic action. Biotechnol. Bioact. Compd. Sources Appl. 2015, 565–581. [Google Scholar]
- Lee, K.H.; Woo, K.S.; Yong, H.I.; Jo, C.; Lee, S.K.; Lee, B.W.; Oh, S.-K.; Lee, Y.-Y.; Lee, B.; Kim, H.-J. Assessment of microbial safety and quality changes of brown and white cooked rice treated with atmospheric pressure plasma. Food Sci. Biotechnol. 2018, 27, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, N.; Bayliss, D.; Chope, G.; Penson, S.; Perehinec, T.; Fisk, I.D. Cold plasma: A new technology to modify wheat flour functionality. Food Chem. 2016, 202, 247–253. [Google Scholar] [CrossRef]
- Van Durme, J.; Vandamme, J. Non-thermal plasma as preparative technique to evaluate olive oil adulteration. Food Chem. 2016, 208, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Wąsowicz, E.; Gramza, A.; Hęś, M.; Jeleñ, H.H.; Korczak, J.; Małecka, M.; Mildner-Szkudlarz, S.; Rudziñska, M.; Samotyja, U.; Zawirska-Wojtasiak, R. Oxidation of lipids in food. Pol. J. Food Nutr. Sci. 2004, 13, 87–100. [Google Scholar]
- Yodpitak, S.; Mahatheeranont, S.; Boonyawan, D.; Sookwong, P.; Roytrakul, S.; Norkaew, O. Cold plasma treatment to improve germination and enhance the bioactive phytochemical content of germinated brown rice. Food Chem. 2019, 289, 328–339. [Google Scholar] [CrossRef] [PubMed]
- López-Gámez, G.; Elez-Martínez, P.; Martín-Belloso, O.; Soliva-Fortuny, R. Recent advances toward the application of non-thermal technologies in food processing: An insight on the bioaccessibility of health-related constituents in plant-based products. Foods 2021, 10, 1538. [Google Scholar] [CrossRef]
- Žnidarčič, D.; Ban, D.; Šircelj, H. Carotenoid and chlorophyll composition of commonly consumed leafy vegetables in Mediterranean countries. Food Chem. 2011, 129, 1164–1168. [Google Scholar] [CrossRef]
- Villatoro-Pulido, M.; Priego-Capote, F.; Álvarez-Sánchez, B.; Saha, S.; Philo, M.; Obregón-Cano, S.; De Haro-Bailón, A.; Font, R.; Del Río-Celestino, M. An approach to the phytochemical profiling of rocket [Eruca sativa (Mill.) Thell]. J. Sci. Food Agric. 2013, 93, 3809–3819. [Google Scholar] [CrossRef]
- Dvorakova, M.; Pumprova, K.; Antonínová, Ž.; Rezek, J.; Haisel, D.; Ekrt, L.; Vanek, T.; Langhansova, L. Nutritional and antioxidant potential of fiddleheads from European ferns. Foods 2021, 10, 460. [Google Scholar] [CrossRef] [PubMed]
- Keyata, E.O.; Tola, Y.B.; Bultosa, G.; Forsido, S.F. Phytochemical contents, antioxidant activity and functional properties of Raphanus sativus L, Eruca sativa L. and Hibiscus sabdariffa L. growing in Ethiopia. Heliyon 2021, 7, e05939. [Google Scholar] [CrossRef]
- Fernandes, F.A.; Santos, V.O.; Rodrigues, S. Effects of glow plasma technology on some bioactive compounds of acerola juice. Food Res. Int. 2019, 115, 16–22. [Google Scholar] [CrossRef]
- Martínez, J.M.; Delso, C.; Angulo, J.; Álvarez, I.; Raso, J. Pulsed electric field-assisted extraction of carotenoids from fresh biomass of Rhodotorula glutinis. Innov. Food Sci. Emerg. Technol. 2018, 47, 421–427. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Ramazzina, I.; Berardinelli, A.; Rizzi, F.; Tappi, S.; Ragni, L.; Sacchetti, G.; Rocculi, P. Effect of cold plasma treatment on physico-chemical parameters and antioxidant activity of minimally processed kiwifruit. Postharvest Biol. Technol. 2015, 107, 55–65. [Google Scholar] [CrossRef]
- Santos Jr, L.; Cubas, A.; Moecke, E.; Ribeiro, D.; Amante, E. Use of cold plasma to inactivate Escherichia coli and physicochemical evaluation in pumpkin puree. J. Food Prot. 2018, 81, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.; Adhikari, M.; Ghimire, B.; Park, G.; Choi, E.H. Cold atmospheric plasma-activated water irrigation induces defense hormone and gene expression in tomato seedlings. Sci. Rep. 2019, 9, 16080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nzekoue, F.K.; Caprioli, G.; Ricciutelli, M.; Cortese, M.; Alesi, A.; Vittori, S.; Sagratini, G. Development of an innovative phytosterol derivatization method to improve the HPLC-DAD analysis and the ESI-MS detection of plant sterols/stanols. Food Res. Int. 2020, 131, 108998. [Google Scholar] [CrossRef]
- Caprioli, G.; Kamgang Nzekoue, F.; Fiorini, D.; Scocco, P.; Trabalza-Marinucci, M.; Acuti, G.; Tardella, F.M.; Sagratini, G.; Catorci, A. The effects of feeding supplementation on the nutritional quality of milk and cheese from sheep grazing on dry pasture. Int. J. Food Sci. Nutr. 2020, 71, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Lamichhane, P.; Veerana, M.; Lim, J.S.; Mumtaz, S.; Shrestha, B.; Kaushik, N.K.; Park, G.; Choi, E.H. Low-Temperature Plasma-Assisted Nitrogen Fixation for Corn Plant Growth and Development. Int. J. Mol. Sci. 2021, 22, 5360. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Control | PAW-2 | PAW-5 | P-10 | P-20 | RI | |
|---|---|---|---|---|---|---|---|
| Glucosinolate hydrolysis products (GHPs) | 54.58 ± 1.48 a | 57.02 ± 0.08 a | 49.61 ± 3.85 a | 53.87 ± 4.44 a | 60.44 ± 3.15 a | ||
| 1 | Methyl thiocyanate | 0.04 ± 0.01 c | 0.09 ± 0.01 bc | 0.17 ± 0.04 a | 0.07 ± 0.01 bc | 0.12 ± 0.01 ab | 1282 |
| 2 | 5-Methyl Hexanenitrile | 0.09 ± 0.01 a | 0.10 ± 0.01 a | Nd ± 0.00 c | 0.11 ± 0.01 a | 0.03 ± 0.01 b | 1362 |
| 3 | Heptanonitrile | 0.18 ± 0.04 a | 0.18 ± 0.01 a | Nd ± 0.00 c | 0.15 ± 0.01 a | 0.07 ± 0.01 b | 1403 |
| 4 | 1-Butene 4-isothiocyanate | 0.07 ± 0.01 b | 0.14 ± 0.02 b | 0.56 ± 0.10 a | 0.13 ± 0.04 b | 0.42 ± 0.04 a | 1452 |
| 5 | 4-Methylthio butanenitrile | 0.35 ± 0.11 ab | 0.38 ± 0.08 ab | Nd ± 0.00 c | 0.45 ± 0.08 a | 0.11 ± 0.01 bc | 1784 |
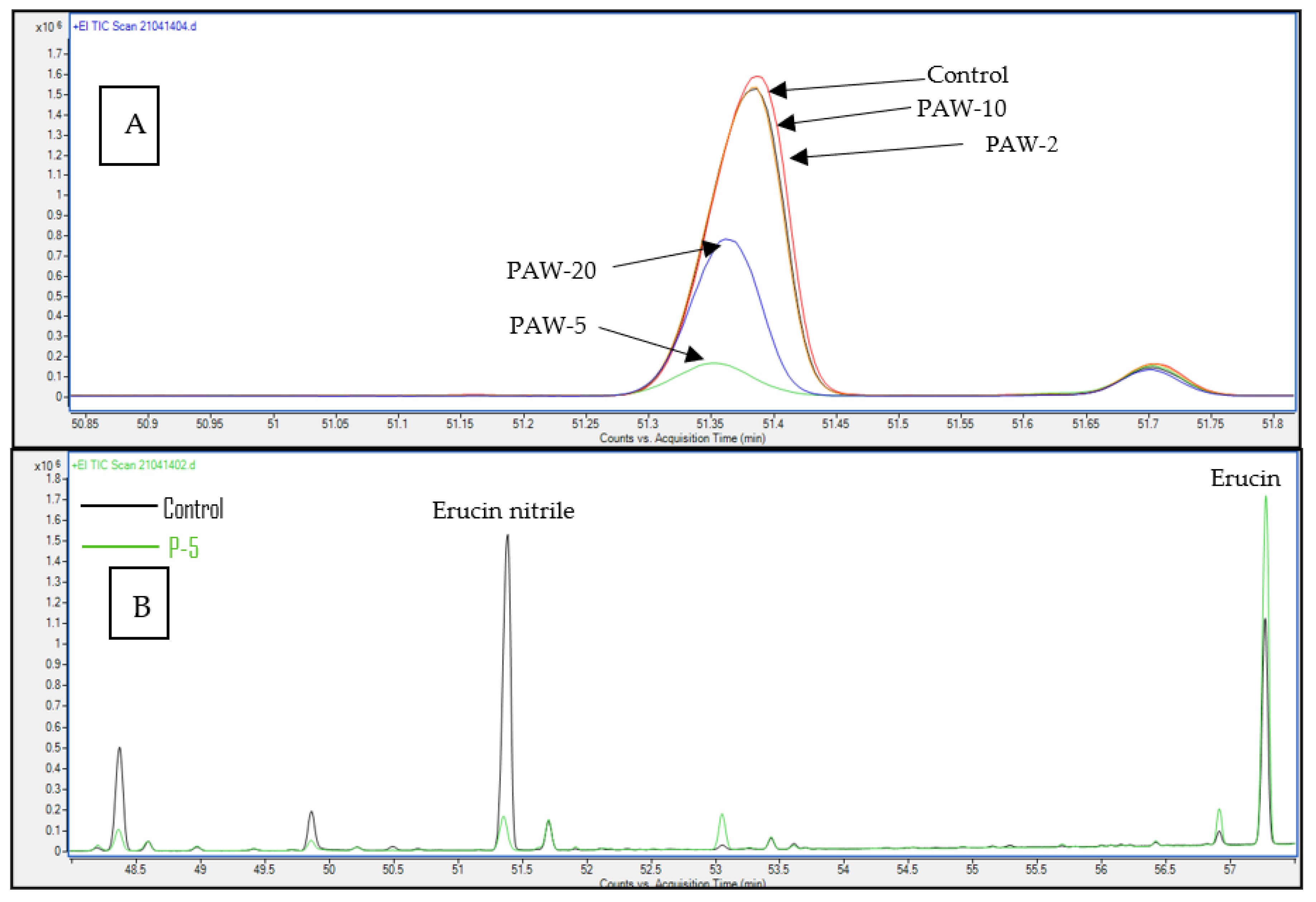
| 6 | Erucin nitrile | 37.52 ± 2.54 a | 34.52 ± 1.60 a | 4.71 ± 0.27 c | 36.91 ± 5.41 a | 21.74 ± 0.06 b | 1935 |
| 7 | Erucin | 16.35 ± 4.16 b | 21.63 ± 0.74 b | 44.17 ± 4.19 a | 16.06 ± 0.82 b | 37.96 ± 3.27 a | 2143 |
| Sulphur compounds | 19.15 ± 0.81 a | 13.23 ± 0.05 bc | 10.08 ± 1.51 c | 15.02 ± 1.03 b | 10.57 ± 1.11 c | ||
| 8 | Methyl disulphide | 0.92 ± 0.11 b | 0.62 ± 0.05 c | 1.47 ± 0.01 a | 0.54 ± 0.01 c | 1.14 ± 0.05 b | 746 |
| 9 | Dimethyl sulphide | 3.30 ± 1.56 a | 3.32 ± 0.35 a | 3.22 ± 1.20 a | 3.57 ± 0.01 a | 3.82 ± 0.57 a | 760 |
| 10 | Dimethyl trisulphide | 0.18 ± 0.04 a | 0.08 ± 0.01 a | 0.33 ± 0.05 a | 0.14 ± 0.04 a | 0.07 ± 0.01 a | 1385 |
| 11 | Dimethyl Sulfoxide | 1.37 ± 0.19 a | 1.86 ± 0.18 a | 1.91 ± 0.21 a | 1.77 ± 0.13 a | 1.66 ± 0.11 a | 1577 |
| 12 | Dihydro-2H-thiopyran-3(4H)-one | 13.09 ± 2.64 a | 6.98 ± 0.56 bc | 2.79 ± 0.12 c | 8.76 ± 0.82 ab | 3.49 ± 0.37 c | 1845 |
| 13 | Dimethyl sulfone | 0.31 ± 0.01 a | 0.39 ± 0.04 a | 0.38 ± 0.04 a | 0.25 ± 0.05 a | 0.40 ± 0.03 a | 1899 |
| Ketones | 10.20 ± 0.81 b | 13.85 ± 0.73 ab | 15.48 ± 1.66 a | 11.64 ± 0.72 ab | 11.36 ± 0.58 b | ||
| 14 | 2,5-Dimethyl-3-hexanone | 0.04 ± 0.01 bc | Nd ± 0.00 d | 0.20 ± 0.01 a | 0.12 ± 0.02 b | 0.10 ± 0.04 b | 1188 |
| 15 | 3-Hydroxybutan-2-one | 0.06 ± 0.01 a | 0.05 ± 0.00 a | 0.07 ± 0.01 a | 0.05 ± 0.01 a | 0.05 ± 0.01 a | 1297 |
| 16 | 1-Hydroxypropan-2-one | 0.08 ± 0.02 ab | 0.05 ± 0.01 ab | 0.13 ± 0.04 a | 0.03 ± 0.00 b | 0.06 ± 0.02 ab | 1309 |
| 17 | 6-Methyl-5-hepten-2-one | 0.41 ± 0.01 b | 0.95 ± 0.10 a | 0.46 ± 0.05 b | 0.52 ± 0.09 b | 0.39 ± 0.07 b | 1344 |
| 18 | 3-Octen-2-one | 0.05 ± 0.01 b | 0.12 ± 0.01 b | 0.28 ± 0.04 a | 0.10 ± 0.02 b | 0.09 ± 0.00 b | 1409 |
| 19 | 3,5-Octadien-2-one | 0.20 ± 0.07 b | 0.74 ± 0.01 ab | 0.82 ± 0.21 a | 0.64 ± 0.09 ab | 0.85 ± 0.23 a | 1514 |
| 20 | (3E,5E)-3,5-Octandiene-2-one | 0.79 ± 0.41 a | 1.27 ± 0.08 a | 1.30 ± 0.15 a | 1.00 ± 0.07 a | 1.28 ± 0.11 a | 1565 |
| 21 | 6-Methyl-3,5-heptadien-2-one | 0.18 ± 0.04 a | 0.24 ± 0.00 a | 0.34 ± 0.08 a | 0.24 ± 0.08 a | 0.15 ± 0.01 a | 1587 |
| 22 | (E)-β-Ionone | 2.73 ± 0.30 b | 2.96 ± 0.21 b | 3.70 ± 0.02 a | 2.75 ± 0.07 b | 3.11 ± 0.04 ab | 1945 |
| 23 | β-Ionone-5,6-epoxide Norisoprenoid | 0.97 ± 0.04 c | 1.21 ± 0.06 ab | 1.27 ± 0.07 ab | 1.40 ± 0.04 a | 1.12 ± 0.04 bc | 1999 |
| 24 | 6,10,14-Trimethylpentadecan-2-one | 1.61 ± 0.54 b | 1.75 ± 0.04 ab | 3.12 ± 0.59 a | 1.27 ± 0.07 b | 1.39 ± 0.01 b | 2129 |
| 25 | (E)-geranylacetone | 0.99 ± 0.25 ab | 1.64 ± 0.24 a | 1.09 ± 0.08 ab | 1.00 ± 0.09 ab | 0.85 ± 0.01 b | 1852 |
| 26 | Dihydroactinidiolide Norisoprenoid | 2.12 ± 0.13 ab | 2.87 ± 0.24 a | 2.73 ± 0.32 ab | 2.56 ± 0.06 ab | 1.95 ± 0.10 b | 2371 |
| Aldehydes | 6.48 ± 0.13 b | 6.84 ± 0.30 b | 12.71 ± 0.07 a | 7.75 ± 0.96 b | 8.00 ± 0.88 b | ||
| 27 | 2-Methyl propanal | 0.87 ± 0.10 b | 1.14 ± 0.08 b | 2.26 ± 0.14 a | 1.20 ± 0.17 b | 1.22 ± 0.22 b | 810 |
| 28 | 2-Methyl butanal | 0.77 ± 0.01 b | 0.56 ± 0.02 c | 1.53 ± 0.03 a | 0.64 ± 0.05 bc | 0.69 ± 0.06 bc | 911 |
| 29 | 3-Methyl butanal | 0.77 ± 0.15 ab | 0.34 ± 0.04 c | 0.79 ± 0.02 a | 0.45 ± 0.10bc | 0.47 ± 0.03 bc | 915 |
| 30 | Pentanal | 0.11 ± 0.01 b | 0.12 ± 0.01 b | 0.19 ± 0.02 a | 0.16 ± 0.01 ab | 0.11 ± 0.02 b | 976 |
| 31 | Hexanal | 0.15 ± 0.04 b | 0.19 ± 0.01 b | 0.33 ± 0.05 a | 0.18 ± 0.04 b | 0.10 ± 0.00 b | 1088 |
| 32 | 2-hexenal (E) | 0.41 ± 0.02 a | 0.44 ± 0.06 a | 0.78 ± 0.32 a | 0.65 ± 0.21 a | 0.32 ± 0.13 a | 1236 |
| 33 | Octanal | 0.11 ± 0.01 bc | 0.15 ± 0.01 bc | 0.29 ± 0.02 a | 0.23 ± 0.05 ab | 0.23 ± 0.01 ab | 1299 |
| 34 | Nonanal | 0.83 ± 0.10 bc | 0.62 ± 0.03 c | 0.92 ± 0.02 ab | 0.61 ± 0.02 c | 1.12 ± 0.10 a | 1395 |
| 35 | 3-Furfural | 0.17 ± 0.04 c | 0.18 ± 0.03 c | 0.85 ± 0.05 a | 0.16 ± 0.10 c | 0.55 ± 0.08 b | 1459 |
| 36 | Benzaldehyde | 1.15 ± 0.05 c | 1.62 ± 0.01 bc | 2.33 ± 0.01 a | 1.79 ± 0.05 ab | 1.28 ± 0.33 bc | 1518 |
| 37 | β-cyclocitral | 0.59 ± 0.01 b | 0.81 ± 0.05 ab | 1.00 ± 0.07 a | 0.74 ± 0.17 ab | 0.73 ± 0.01 ab | 1619 |
| 38 | Benzene acetaldehyde | 0.39 ± 0.02 a | 0.49 ± 0.19 a | 0.66 ± 2.16 a | 0.42 ± 0.11 a | 0.58 ± 0.06 a | 1635 |
| 39 | 2-Methyl benzaldehyde | 0.20 ± 0.04 a | 0.22 ± 0.09 a | 0.83 ± 0.32 a | 0.55 ± 0.13 a | 0.63 ± 0.06 a | 1643 |
| Fatty acids and esters | 2.16 ± 0.07 b | 1.66 ± 0.13 b | 3.99 ± 0.34 a | 1.96 ± 0.30 b | 2.52 ± 0.11 b | ||
| 40 | Acetic acid | 1.23 ± 0.31 b | 1.09 ± 0.04 b | 2.37 ± 0.25 a | 1.23 ± 0.23 b | 1.71 ± 0.07 ab | 1447 |
| 41 | Propanoic acid | 0.05 ± 0.01 b | 0.06 ± 0.00 ab | 0.11 ± 0.02 a | 0.08 ± 0.02 ab | 0.08 ± 0.01 ab | 1530 |
| 42 | Hexanoic acid | 0.24 ± 0.04 b | 0.33 ± 0.04 ab | 0.54 ± 0.11 a | 0.23 ± 0.04 b | 0.44 ± 0.05 ab | 1840 |
| 43 | Methyl palmitate | 0.65 ± 0.21 ab | 0.19 ± 0.05 c | 0.99 ± 0.04 a | 0.43 ± 0.01 bc | 0.30 ± 0.01 bc | 2221 |
| Alcohols | 1.14 ± 0.10 b | 1.61 ± 0.08 a | 1.23 ± 0.08 ab | 1.57 ± 0.13 a | 1.39 ± 0.11 ab | ||
| 44 | Pent-1-en-3-ol | 0.12 ± 0.01 b | 0.31 ± 0.02 a | 0.14 ± 0.01 b | 0.25 ± 0.07 ab | 0.17 ± 0.01 b | 1180 |
| 45 | pentan-1-ol | 0.06 ± 0.01 b | 0.10 ± 0.01 a | 0.08 ± 0.01 ab | 0.06 ± 0.01 b | 0.07 ± 0.01 ab | 1267 |
| 46 | (Z)-2-penten-1-ol | 0.04 ± 0.01 c | 0.10 ± 0.01 b | 0.07 ± 0.01 bc | 0.16 ± 0.03 a | 0.06 ± 0.01 bc | 1329 |
| 47 | Hexan-1-ol | 0.05 ± 0.01 b | 0.09 ± 0.00 ab | 0.11 ± 0.01 a | 0.08 ± 0.01 ab | 0.10 ± 0.01 a | 1360 |
| 48 | Hex-3-ene -1-ol | 0.27 ± 0.10 b | 0.38 ± 0.01 ab | 0.26 ± 0.02 b | 0.51 ± 0.01 a | 0.20 ± 0.04 b | 1388 |
| 49 | Octan-1-ol | 0.40 ± 0.04 b | 0.34 ± 0.02 b | 0.50 ± 0.03 ab | 0.36 ± 0.04 b | 0.65 ± 0.07 a | 1552 |
| 50 | Nonan-1-ol | 0.08 ± 0.01 ab | 0.11 ± 0.01 a | 0.09 ± 0.01 a | 0.07 ± 0.01 ab | 0.04 ± 0.01 b | 1654 |
| 51 | Phenylethyl alcohol | 0.14 ± 0.02 b | 0.20 ± 0.01 a | Nd ± 0.00 c | 0.09 ± 0.02 b | 0.10 ± 0.01 b | 1913 |
| Alkanes | 0.16 ± 0.01 c | 0.12 ± 0.06 c | 0.95 ± 0.00 a | 0.40 ± 0.03 b | 0.47 ± 0.05 b | ||
| 52 | Undecane | 0.16 ± 0.01 c | 0.12 ± 0.06 c | 0.95 ± 0.01 a | 0.40 ± 0.03 a | 0.47 ± 0.05 b | 1094 |
| Control | PAW-2 | PAW-5 | PAW-10 | PAW-20 | |
|---|---|---|---|---|---|
| Chla | 145.0 ± 4.1 a | 114.4 ± 15.2 ab | 111.4 ± 21.6 ab | 131.6 ± 9.3 ab | 102.4 ± 8.4 b |
| Chlb | 98.5 ± 13.4 b | 121.9 ± 17.3 a | 95.9 ± 11.8 b | 111.6 ± 3.3 ab | 100.4 ± 14.4 ab |
| Total | 243.4 ± 17.1 a | 236.3 ± 32.3 a | 207.3 ± 32.1 b | 243.2 ± 12.6 a | 202.7 ± 22.8 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abouelenein, D.; Angeloni, S.; Caprioli, G.; Genovese, J.; Mustafa, A.M.; Nzekoue, F.K.; Petrelli, R.; Rocculi, P.; Sagratini, G.; Tappi, S.; et al. Effect of Plasma Activated Water on Selected Chemical Compounds of Rocket-Salad (Eruca sativa Mill.) Leaves. Molecules 2021, 26, 7691. https://doi.org/10.3390/molecules26247691
Abouelenein D, Angeloni S, Caprioli G, Genovese J, Mustafa AM, Nzekoue FK, Petrelli R, Rocculi P, Sagratini G, Tappi S, et al. Effect of Plasma Activated Water on Selected Chemical Compounds of Rocket-Salad (Eruca sativa Mill.) Leaves. Molecules. 2021; 26(24):7691. https://doi.org/10.3390/molecules26247691
Chicago/Turabian StyleAbouelenein, Doaa, Simone Angeloni, Giovanni Caprioli, Jessica Genovese, Ahmed M. Mustafa, Franks Kamgang Nzekoue, Riccardo Petrelli, Pietro Rocculi, Gianni Sagratini, Silvia Tappi, and et al. 2021. "Effect of Plasma Activated Water on Selected Chemical Compounds of Rocket-Salad (Eruca sativa Mill.) Leaves" Molecules 26, no. 24: 7691. https://doi.org/10.3390/molecules26247691
APA StyleAbouelenein, D., Angeloni, S., Caprioli, G., Genovese, J., Mustafa, A. M., Nzekoue, F. K., Petrelli, R., Rocculi, P., Sagratini, G., Tappi, S., Torregiani, E., & Vittori, S. (2021). Effect of Plasma Activated Water on Selected Chemical Compounds of Rocket-Salad (Eruca sativa Mill.) Leaves. Molecules, 26(24), 7691. https://doi.org/10.3390/molecules26247691