
Inhibition of Fungal Strains Isolated from Cereal Grains via Vapor Phase of Essential Oils

 and
and 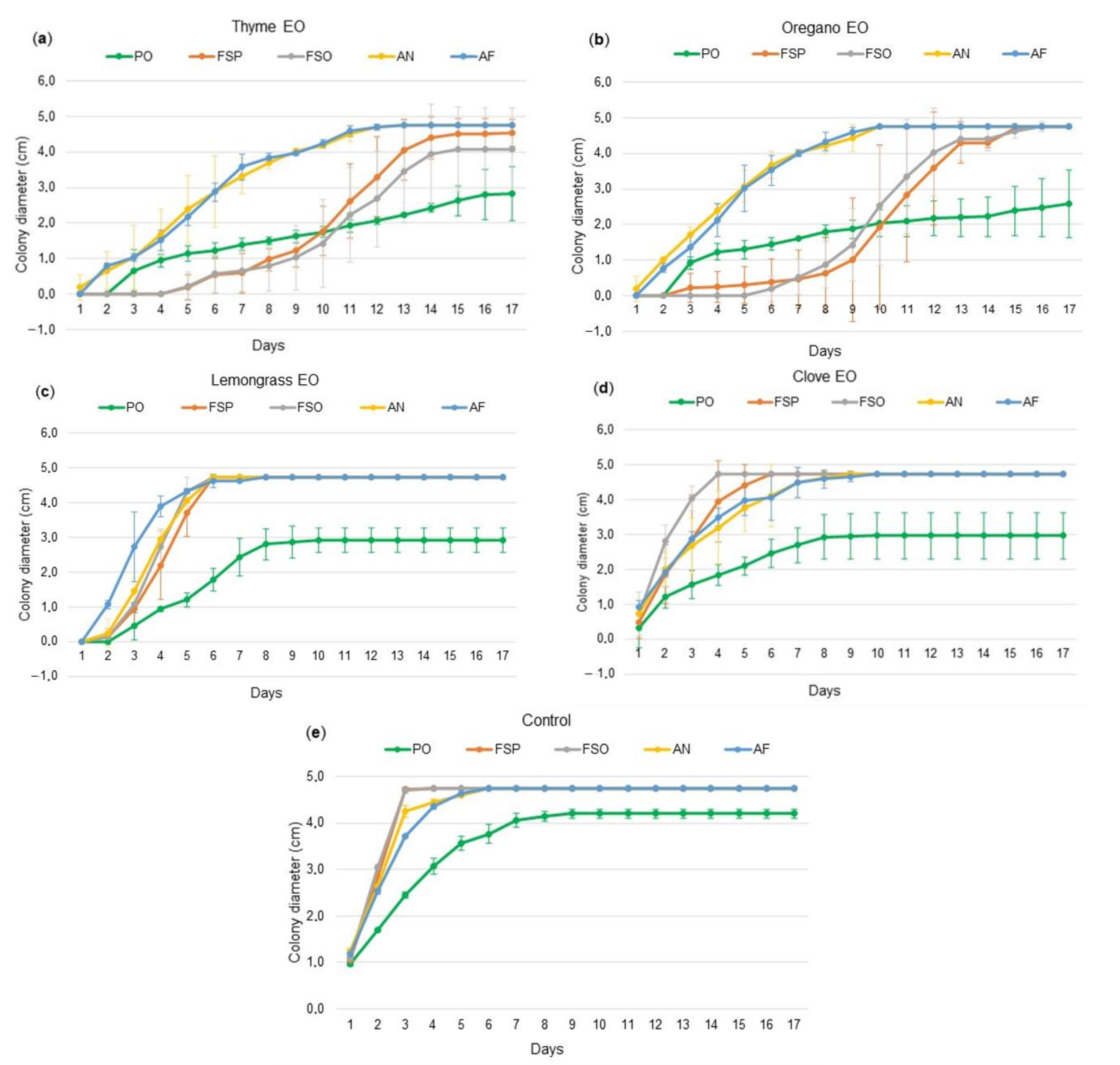{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Compositions of Essential Oils
2.2. Antifungal Effect of the Essential Oils
3. Discussion
4. Materials and Methods
4.1. Essential Oils
4.2. Essential Oil Analysis
4.3. Microorganisms
4.4. Determination of Microorganisms
4.5. Antimicrobial Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Mohapatra, D.; Kumar, S.; Kotwaliwale, N.; Singh, K.K. Critical factors responsible for fungi growth in stored food grains and non-chemical approaches for their control. Ind. Crops Prod. 2017, 108, 162–182. [Google Scholar] [CrossRef]
- Roselló, J.; Sempere, F.; Sanz-Berzosa, I.; Chiralt, A.; Santamarina, M.P. Antifungal activity and potential use of essential oils against Fusarium culmorum and Fusarium verticillioides. J. Essent. Oil Bearing Plants 2015, 18, 359–367. [Google Scholar] [CrossRef]
- Raila, A.; Lugauskas, A.; Steponavičius, D.; Railienė, M.; Steponavičienė, A.; Zvicevicius, E. E. Application of Ozone for Reduction of Mycological Infection. Ann. Agric. Environ. Med. 2006, 13, 287–294. [Google Scholar]
- Ishaq, M.; Tahira, R.; Javed, A.; Jamal, A.; Raja, M.U.; Munir, A.; Ur-Rehman, A. Lemongrass essential oil as an alternate approach to manage seed associated fungi of wheat and rice. Int. J. Agric. Biol. 2017, 19, 1301–1306. [Google Scholar] [CrossRef]
- Kedia, A.; Prakash, B.; Mishra, P.K.; Dubey, N.K. Antifungal and antiaflatoxigenic properties of Cuminum cyminum (L.) seed essential oil and its efficacy as a preservative in stored commodities. Int. J. Food Microbiol. 2014, 168–169, 1–7. [Google Scholar] [CrossRef]
- Kumar, S.A.; Chandrabhan, S.; Shriram, P. Isolation of Aspergillus flavus from stored food commodities and Thymus vulgaris (L.) essential oil used as a safe plant based preservative. Pharmacogn. Mag. 2009, 5, 343–349. [Google Scholar] [CrossRef]
- Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.C.; Nebbia, C.S.; Nielsen, E.; et al. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18. [Google Scholar] [CrossRef]
- Sreenivasa, M.Y.; Dass, R.S.; Charith Raj, A.P.; Nagendra Prasad, M.N.; Achar, P.N.; Janardhana, G.R. Assessment of the growth inhibiting effect of some plant essential oils on different Fusarium species isolated from sorghum and maize grains. J. Plant Dis. Prot. 2011, 118, 208–213. [Google Scholar] [CrossRef]
- Anžlovar, S.; Likar, M.; Koce, J.D. Antifungal potential of thyme essential oil as a preservative for storage of wheat seeds. Acta Bot. Croat. 2017, 76, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Sumalan, R.M.; Alexa, E.; Poiana, M.A. Assessment of inhibitory potential of essential oils on natural mycoflora and Fusarium mycotoxins production in wheat. Chem. Cent. J. 2013, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Matusinsky, P.; Zouhar, M.; Pavela, R.; Novy, P. Antifungal effect of five essential oils against important pathogenic fungi of cereals. Ind. Crops Prod. 2015, 67, 208–215. [Google Scholar] [CrossRef]
- Gutiérrez, L.; Sánchez, C.; Batlle, R.; Nerín, C. New antimicrobial active package for bakery products. Trends Food Sci. Technol. 2009, 20, 92–99. [Google Scholar] [CrossRef]
- Božik, M.; Císarová, M.; Tančinová, D.; Kouřimská, L.; Hleba, L.; Klouček, P. Selected essential oil vapours inhibit growth of Aspergillus spp. in oats with improved consumer acceptability. Ind. Crops Prod. 2017, 98, 146–152. [Google Scholar] [CrossRef]
- Císarová, M.; Hleba, L.; Medo, J.; Tančinová, D.; Mašková, Z.; Čuboň, J.; Kováčik, A.; Foltinová, D.; Božik, M.; Klouček, P. The in vitro and in situ effect of selected essential oils in vapour phase against bread spoilage toxicogenic aspergilli. Food Control 2020, 110. [Google Scholar] [CrossRef]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Equisetum arvense hydro-alcoholic extract: Phenolic composition and antifungal and antimycotoxigenic effect against Aspergillus flavus and Fusarium verticillioides in stored maize. J. Sci. Food Agric. 2013, 93, 2248–2253. [Google Scholar] [CrossRef] [PubMed]
- Tzortzakis, N.G.; Economakis, C.D. Antifungal activity of lemongrass (Cympopogon citratus L.) essential oil against key postharvest pathogens. Innov. Food Sci. Emerg. Technol. 2007, 8, 253–258. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, S.; Kumar, A.; Sharma, A.K. Antifungal efficacy of plant essential oils against stored grain fungi of Fusarium spp. J. Food Sci. Technol. 2016, 53, 3725–3734. [Google Scholar] [CrossRef] [Green Version]
- Maxia, A.; Falconieri, D.; Piras, A.; Porcedda, S.; Marongiu, B.; Frau, M.A.; Gonçalves, M.J.; Cabral, C.; Cavaleiro, C.; Salgueiro, L. Chemical Composition and Antifungal Activity of Essential Oils and Supercritical CO2Extracts of Apium nodiflorum (L.) Lag. Mycopathologia 2012, 174, 61–67. [Google Scholar] [CrossRef]
- Alexa, E.; Sumalan, R.M.; Danciu, C.; Obistioiu, D.; Negrea, M.; Poiana, M.A.; Rus, C.; Radulov, I.; Pop, G.; Dehelean, C. Synergistic antifungal, allelopatic and anti-proliferative potential of salvia officinalis L., and thymus vulgaris L. Essential oils. Molecules 2018, 23, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Shao, Y.L.; Tang, Y.J.; Zhou, W.W. Antifungal activity of essential oil compounds (geraniol and citral) and inhibitory mechanisms on grain pathogens (aspergillus flavus and aspergillus ochraceus). Molecules 2018, 23, 2108. [Google Scholar] [CrossRef] [Green Version]
- Kloucek, P.; Smid, J.; Frankova, A.; Kokoska, L.; Valterova, I.; Pavela, R. Fast screening method for assessment of antimicrobial activity of essential oils in vapor phase. Food Res. Int. 2012, 47, 161–165. [Google Scholar] [CrossRef]
- Morcia, C.; Malnati, M.; Terzi, V. In vitro antifungal activity of terpinen-4-ol, eugenol, carvone, 1,8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2012, 29, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential oils: Sources of antimicrobials and food preservatives. Front. Microbiol. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, R.C.; Carvajal-Moreno, M.; Correa, B.; Rojo-Callejas, F. Cellular, physiological and molecular approaches to investigate the antifungal and anti-aflatoxigenic effects of thyme essential oil on Aspergillus flavus. Food Chem. 2020, 315. [Google Scholar] [CrossRef]
- Mateo, E.M.; Gómez, J.V.; Domínguez, I.; Gimeno-Adelantado, J.V.; Mateo-Castro, R.; Gavara, R.; Jiménez, M. Impact of bioactive packaging systems based on EVOH films and essential oils in the control of aflatoxigenic fungi and aflatoxin production in maize. Int. J. Food Microbiol. 2017, 254, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, G.; Maurya, S.; de Lampasona, M.P.; Catalan, C. Chemical constituents, antifungal and antioxidative potential of Foeniculum vulgare volatile oil and its acetone extract. Food Control 2006, 17, 745–752. [Google Scholar] [CrossRef]
- Tullio, V.; Nostro, A.; Mandras, N.; Dugo, P.; Banche, G.; Cannatelli, M.A.; Cuffini, A.M.; Alonzo, V.; Carlone, N.A. Antifungal activity of essential oils against filamentous fungi determined by broth microdilution and vapour contact methods. J. Appl. Microbiol. 2007, 102, 1544–1550. [Google Scholar] [CrossRef]
- Vilela, G.R.; de Almeida, G.S.; D’Arce, M.A.B.R.; Moraes, M.H.D.; Brito, J.O.; da Silva, M.F.d.G.F.; Silva, S.C.; de Stefano Piedade, S.M.; Calori-Domingues, M.A.; da Gloria, E.M. Activity of essential oil and its major compound, 1,8-cineole, from Eucalyptus globulus Labill., against the storage fungi Aspergillus flavus Link and Aspergillus parasiticus Speare. J. Stored Prod. Res. 2009, 45, 108–111. [Google Scholar] [CrossRef]
- Gómez, J.V.; Tarazona, A.; Mateo-Castro, R.; Gimeno-Adelantado, J.V.; Jiménez, M.; Mateo, E.M. Selected plant essential oils and their main active components, a promising approach to inhibit aflatoxigenic fungi and aflatoxin production in food. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2018, 35, 1581–1595. [Google Scholar] [CrossRef]
- Tyagi, A.K.; Malik, A. Antimicrobial potential and chemical composition of Eucalyptus globulus oil in liquid and vapour phase against food spoilage microorganisms. Food Chem. 2011, 126, 228–235. [Google Scholar] [CrossRef]
- Tyagi, A.K.; Malik, A. Antimicrobial potential and chemical composition of Mentha piperita oil in liquid and vapour phase against food spoiling microorganisms. Food Control 2011, 22, 1707–1714. [Google Scholar] [CrossRef]
- Yahyazadeh, M.; Omidbaigi, R.; Zare, R.; Taheri, H. Effect of some essential oils on mycelial growth of Penicillium digitatum Sacc. World J. Microbiol. Biotechnol. 2008, 24, 1445–1450. [Google Scholar] [CrossRef]
- Yigit, F.; Özcan, M.; Akgül, A. Inhibitory effect of some spice essential oils on Penicillium digitatum causing postharvest rot in citrus. Grasas Aceites 2000, 51, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Bluma, R.; Landa, M.F.; Etcheverry, M. Impact of volatile compounds generated by essential oils on Aspergillus section Flavi growth parameters and aflatoxin accumulation. J. Sci. Food Agric. 2009, 89, 1473–1480. [Google Scholar] [CrossRef]
- Linde, G.A.; Gazim, Z.C.; Cardoso, B.K.; Jorge, L.F.; Tešević, V.; Glamoćlija, J.; Soković, M.; Colauto, N.B. Antifungal and antibacterial activities of Petroselinum crispum essential oil. Genet. Mol. Res. 2016, 15, 1–11. [Google Scholar] [CrossRef]
- Jahani, M.; Pira, M.; Aminifard, M.H. Antifungal effects of essential oils against Aspergillus niger in vitro and in vivo on pomegranate (Punica granatum) fruits. Sci. Hortic. 2020, 264, 109188. [Google Scholar] [CrossRef]
- Kohiyama, C.Y.; Yamamoto Ribeiro, M.M.; Mossini, S.A.G.; Bando, E.; Bomfim, N.D.S.; Nerilo, S.B.; Rocha, G.H.O.; Grespan, R.; Mikcha, J.M.G.; Machinski, M. Antifungal properties and inhibitory effects upon aflatoxin production of Thymus vulgaris L. by Aspergillus flavus Link. Food Chem. 2015, 173, 1006–1010. [Google Scholar] [CrossRef] [Green Version]
- Krzyśko-Łupicka, T.; Sokół, S.; Piekarska-Stachowiak, A. Evaluation of fungistatic activity of eight selected essential oils on four heterogeneous Fusarium isolates obtained from cereal grains in southern Poland. Molecules 2020, 25, 292. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Kistlerr, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Hubka, V.; Kolarik, M. β-tubulin paralogue tubC is frequently misidentified as the benA gene in Aspergillus section Nigri taxonomy: Primer specificity testing and taxonomic consequences. Persoonia Mol. Phylogeny Evol. Fungi 2012, 29, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, T. NCBI_blast Information. In The NCBI Handbook; National Center for Biotechnology Information: Bethesda, MD, USA, 2013; pp. 1–15. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Střelková, T.; Nemes, B.; Kovács, A.; Novotný, D.; Božik, M.; Klouček, P. Inhibition of Fungal Strains Isolated from Cereal Grains via Vapor Phase of Essential Oils. Molecules 2021, 26, 1313. https://doi.org/10.3390/molecules26051313
Střelková T, Nemes B, Kovács A, Novotný D, Božik M, Klouček P. Inhibition of Fungal Strains Isolated from Cereal Grains via Vapor Phase of Essential Oils. Molecules. 2021; 26(5):1313. https://doi.org/10.3390/molecules26051313
Chicago/Turabian StyleStřelková, Tereza, Bence Nemes, Anett Kovács, David Novotný, Matěj Božik, and Pavel Klouček. 2021. "Inhibition of Fungal Strains Isolated from Cereal Grains via Vapor Phase of Essential Oils" Molecules 26, no. 5: 1313. https://doi.org/10.3390/molecules26051313
APA StyleStřelková, T., Nemes, B., Kovács, A., Novotný, D., Božik, M., & Klouček, P. (2021). Inhibition of Fungal Strains Isolated from Cereal Grains via Vapor Phase of Essential Oils. Molecules, 26(5), 1313. https://doi.org/10.3390/molecules26051313