
ACTH(6–9)PGP Peptide Protects SH-SY5Y Cells from H2O2, tert-Butyl Hydroperoxide, and Cyanide Cytotoxicity via Stimulation of Proliferation and Induction of Prosurvival-Related Genes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ACTH(6–9)PGP Stimulated Cell Proliferation and Increased Cell Survival after H2O2, tert-Butyl Hydroperoxide, MPP+, and KCN Treatment
2.2. ACTH(6–9)PGP Decreased Apoptosis and Increased Cell Viability but Did Not Affect Acute ROS Level
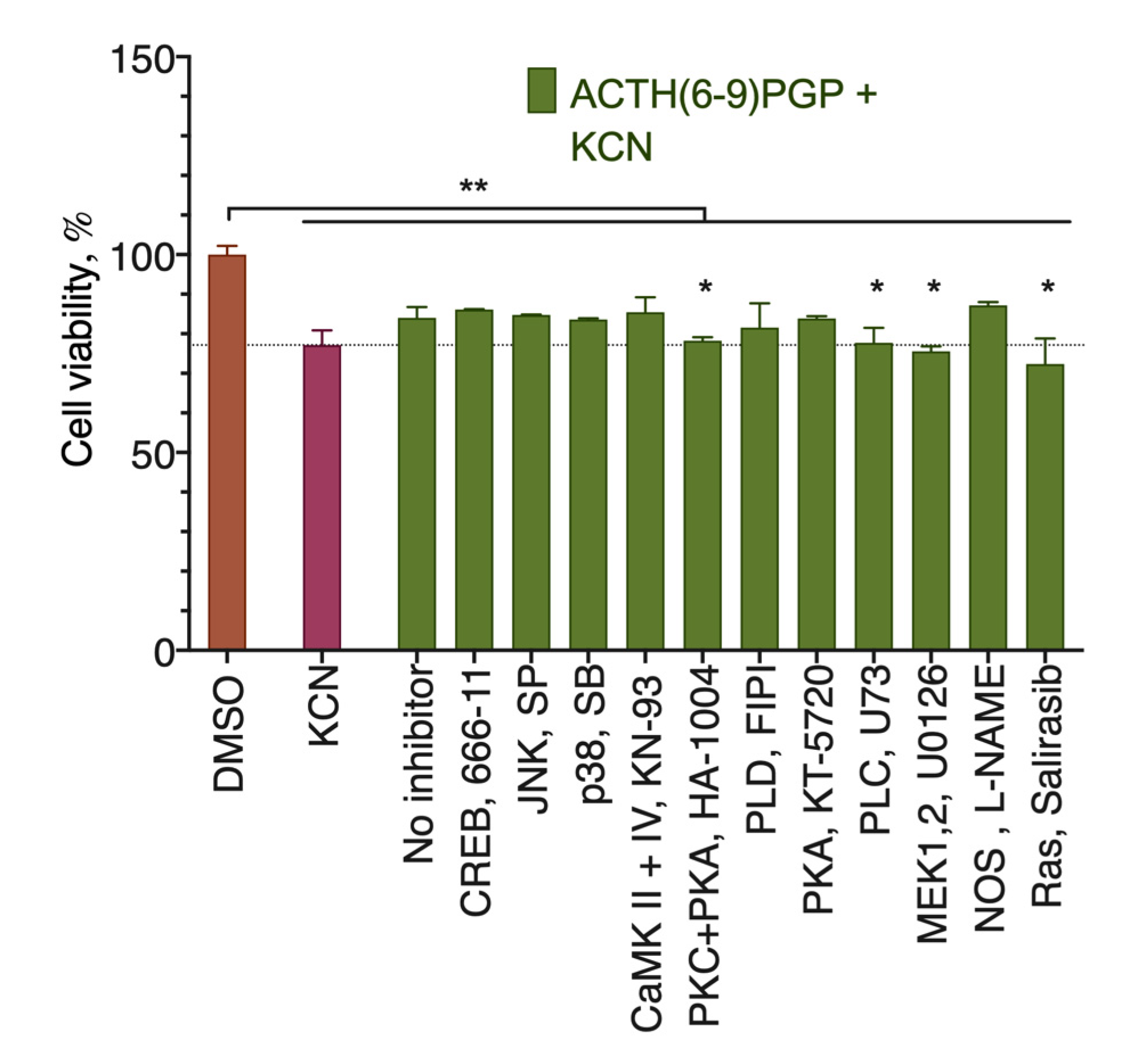
2.3. ACTH(6–9)PGP Protection against KCN Cytotoxicity Is Inhibited by the MEK, PKC, PLC, and Ras Inhibitors
2.4. ACTH(6–9)PGP Did Not Alter Cellular cAMP Content
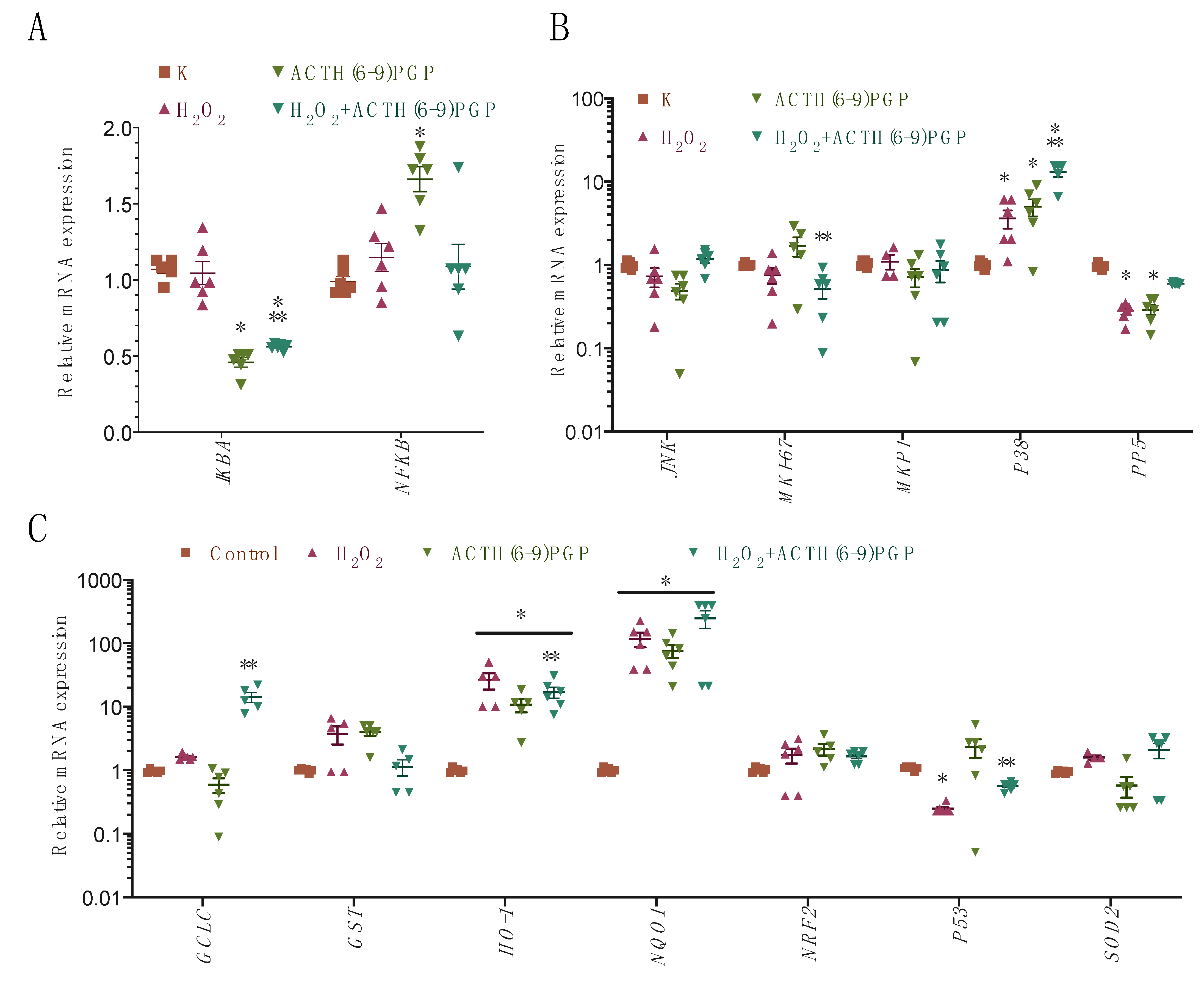
2.5. ACTH(6–9)PGP Decreased the Expression of the NF-κB- and Nrf-2-Related Genes but Not of the Antioxidant Enzymes
- NF-κB pathway: AKT3, IκB, and NF-κB;
- Nrf-2 pathway: Nrf2, HO-1, GST, NQO1, GCLC, SOD1, SOD2, and CAT (catalase);
- MAPK pathway: JNK, P38, MKP1, PP2A, PP5, and Ki-67;
- DNA-damage-related pathway: P53.
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Oxidative Stress Induction
4.4. Cell Viability Assay
4.5. cAMP Assay
4.6. ROS Assay
4.7. Apoptosis Assay
4.8. mRNA Assay
4.9. Proliferation Assay via the BrdU Incorporation
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Starowicz, K.; Przewłocka, B. The role of melanocortins and their receptors in inflammatory processes, nerve regeneration and nociception. Life Sci. 2003, 73, 823–847. [Google Scholar] [CrossRef]
- De Wied, D. Neuropeptides in learning and memory processes. Behav. Brain Res. 1997, 83, 83–90. [Google Scholar] [CrossRef]
- De Wied, D.; De Kloet, E.R. Pro-opiomelanocortin (POMC) as Homeostatic Control System. Ann. N. Y. Acad. Sci. 1987, 512, 328–337. [Google Scholar] [CrossRef]
- Ashmarin, I.P.; Nezavibat’ko, V.N.; Myasoedov, N.F.; Kamenskii, A.A.; Grivennikov, I.A.; Ponomareva-Stepnaya, M.A.; Andreeva, L.A.; Kaplan, A.Y.; Koshelev, V.B. A Nootropic Analog of Adrenocorticotrophic Hormone 4-10--Semax (Experience of 15 Years of Development and Investigation). Zh. Vyssh. Nerv. Deyat. 1997, 47, 420–430. [Google Scholar]
- Walter, R.; Simmons, W.; Yoshimoto, T. Proline specific endo- and exopeptidases. Mol. Cell. Biochem. 1980, 30, 111–127. [Google Scholar] [CrossRef]
- Walker, J.R.; Altman, R.K.; Warren, J.W.; Altman, E. Using protein-based motifs to stabilize peptides. J. Pept. Res. 2003, 62, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.F.; Snelgrove, R.J. The multifaceted roles of the matrikine Pro-Gly-Pro in pulmonary health and disease. Eur. Respir. Rev. 2018, 27, 180017. [Google Scholar] [CrossRef]
- Ashmarin, I.P. Glyprolines in regulatory tripeptides. Neurochem. J. 2007, 1, 173–175. [Google Scholar] [CrossRef]
- Ponomareva-Stepnaya, M.A.; Nezavibatko, V.N.; Antonova, L.V.; Andreeva, L.A.; Alfeeva, L.Y.; Potaman, V.N.; Kamensky, A.A.; Ashmarin, I.P. Analogue AKTG4-10—A Stimulant for Learning with Prolonged Action. Khim. Pharm. Zurnal 1984, 7, 790–795. [Google Scholar]
- Shevchenko, K.V.; Nagaev, I.Y.; Andreeva, L.A.; Shevchenko, V.P.; Myasoedov, N.F. Stability of Proline-Containing Peptides in Biological Media. Biochem. (Moscow) Suppl. Ser. B Biomed. Chem. 2019, 13, 179–201. [Google Scholar] [CrossRef]
- Cain, J.P.; Mayorov, A.V.; Cai, M.; Wang, H.; Tan, B.; Chandler, K.; Lee, Y.; Petrov, R.R.; Trivedi, D.; Hruby, V.J. Design, synthesis, and biological evaluation of a new class of small molecule peptide mimetics targeting the melanocortin receptors. Bioorg. Med. Chem. Lett. 2006, 16, 5462–5467. [Google Scholar] [CrossRef] [Green Version]
- Yasenyavskaya, A.; Samotrueva, M.; Tsibizova, A.; Bashkina, O.; Andreeva, L.; Myasoedov, N. Effects of melanocortinson the behavior of ratsin thetest ofelevated cruciform maze and experimentally induced ofsocials. Arch. Euromedica 2020, 10, 35–38. [Google Scholar] [CrossRef]
- Yasenyavskaya, A.; Samotrueva, M.; Tsibizova, A.; Bashkina, O.; Myasoedov, N.; Andreeva, L. Effects of neuropeptides on behavior of rats in open field test and experimentally induced social stress. Arch. Euromedica 2020, 10, 25–28. [Google Scholar] [CrossRef]
- Bakaeva, Z.V.; Surin, A.M.; Lizunova, N.V.; Zgodova, A.E.; Krasilnikova, I.A.; Fisenko, A.P.; Frolov, D.A.; Andreeva, L.A.; Myasoedov, N.F.; Pinelis, V.G. Neuroprotective Potential of Peptides HFRWPGP (ACTH6–9PGP), KKRRPGP, and PyrRP in Cultured Cortical Neurons at Glutamate Excitotoxicity. Dokl. Biochem. Biophys. 2020, 491, 62–66. [Google Scholar] [CrossRef]
- Akimov, M.G.; Ashba, A.M.; Fomina-Ageeva, E.V.; Gretskaya, N.M.; Myasoedov, N.F.; Bezuglov, V.V. Neuroprotective Action of Amidic Neurolipins in Models of Neurotoxicity on the Culture of Human Neural-Like Cells SH-SY5Y. Dokl. Biochem. Biophys. 2019, 485, 141–144. [Google Scholar] [CrossRef]
- Armogida, M.; Nisticò, R.; Mercuri, N.B. Therapeutic potential of targeting hydrogen peroxide metabolism in the treatment of brain ischaemia. Br. J. Pharmacol. 2012, 166, 1211–1224. [Google Scholar] [CrossRef] [Green Version]
- Shirley, R.; Ord, E.N.J.; Work, L.M. Oxidative Stress and the Use of Antioxidants in Stroke. Antioxidants 2014, 3, 472–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-H.; Kim, C.-S.; Lee, Y.J. Astaxanthin protects against MPTP/MPP+-induced mitochondrial dysfunction and ROS production in vivo and in vitro. Food Chem. Toxicol. 2011, 49, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.N.; Kim, S.-G.; Lim, J.-Y.; Dutta, R.K.; Kim, S.-J.; Choe, S.-K.; So, H.-S.; Park, R. 3-Aminotriazole protects from CoCl2-induced ototoxicity by inhibiting the generation of reactive oxygen species and proinflammatory cytokines in mice. Arch. Toxicol. 2015, 90, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Biggs, T.; Cooke, S.; Barton, C.; Harris, M.; Saksela, K.; Mann, D. Induction of activator protein 1 (AP-1) in macrophages by human immunodeficiency virus type-1 NEF is a cell-type-specific response that requires both Hck and MAPK signaling events 1 1Edited by J. Karn. J. Mol. Biol. 1999, 290, 21–35. [Google Scholar] [CrossRef]
- Gentili, C.; Boland, R.; De Boland, A.R. PTH stimulates PLCβ and PLCγ isoenzymes in rat enterocytes: Influence of ageing. Cell. Signal. 2001, 13, 131–138. [Google Scholar] [CrossRef]
- Zhang, H.; Chi, Y.; Gao, K.; Zhang, X.; Yao, J. p53 Protein-mediated Up-regulation of MAP Kinase Phosphatase 3 (MKP-3) Contributes to the Establishment of the Cellular Senescent Phenotype through Dephosphorylation of Extracellular Signal-regulated Kinase 1/2 (ERK1/2). J. Biol. Chem. 2015, 290, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Jaafaru, M.S.; Nordin, N.; Rosli, R.; Shaari, K.; Bako, H.Y.; Saad, N.; Noor, N.M.; Razis, A.F.A. Neuroprotective effects of glucomoringin-isothiocyanate against H2O2-Induced cytotoxicity in neuroblastoma (SH-SY5Y) cells. NeuroToxicology 2019, 75, 89–104. [Google Scholar] [CrossRef]
- Guo, H.; Guo, S.; Liu, H. Antioxidant activity and inhibition of ultraviolet radiation-induced skin damage of Selenium-rich peptide fraction from selenium-rich yeast protein hydrolysate. Bioorg. Chem. 2020, 105, 104431. [Google Scholar] [CrossRef]
- Ma, W.; Zhu, X.; Ding, X.; Li, T.; Hu, Y.; Hu, X.; Yuan, L.; Lei, L.; Hu, A.; Luo, Y.; et al. Protective effects of SS31 on t‑BHP induced oxidative damage in 661W cells. Mol. Med. Rep. 2015, 12, 5026–5034. [Google Scholar] [CrossRef] [Green Version]
- Sobotta, M.C.; Barata, A.G.; Schmidt, U.; Mueller, S.; Millonig, G.; Dick, T.P. Exposing cells to H2O2: A quantitative comparison between continuous low-dose and one-time high-dose treatments. Free Radic. Biol. Med. 2013, 60, 325–335. [Google Scholar] [CrossRef]
- Bhattacharya, R.; Flora, S. Cyanide Toxicity and its Treatment. In Handbook of Toxicology of Chemical Warfare Agents; Elsevier BV: Amsterdam, The Netherlands, 2009; pp. 255–270. [Google Scholar]
- Anantharam, V.; Kaul, S.; Song, C.; Kanthasamy, A.; Kanthasamy, A.G. Pharmacological inhibition of neuronal NADPH oxidase protects against 1-methyl-4-phenylpyridinium (MPP+)-induced oxidative stress and apoptosis in mesencephalic dopaminergic neuronal cells. NeuroToxicology 2007, 28, 988–997. [Google Scholar] [CrossRef] [Green Version]
- Li, D.-W.; Li, G.-R.; Lu, Y.; Liu, Z.-Q.; Chang, M.; Yao, M.; Cheng, W.; Hu, L.-S. α-lipoic acid protects dopaminergic neurons against MPP+-induced apoptosis by attenuating reactive oxygen species formation. Int. J. Mol. Med. 2013, 32, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Alsharif, I.; Boukhzar, L.; Lefranc, B.; Godefroy, D.; Aury-Landas, J.; Rego, J.-L.D.; Naudet, F.; Arabo, A.; Chagraoui, A.; Maltête, D.; et al. Cell-penetrating, antioxidant SELENOT mimetic protects dopaminergic neurons and ameliorates motor dysfunction in Parkinson’s disease animal models. Redox Biol. 2021, 40, 101839. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Zhang, Z.; Li, H.; Ma, J.; Sun, L.; Shao, S.; Zhang, Z.; Hölscher, C. A GLP-2 Analogue Protects SH-SY5Y and Neuro-2a Cells Against Mitochondrial Damage, Autophagy Impairments and Apoptosis in a Parkinson Model. Drug Res. 2021, 71, 43–50. [Google Scholar] [CrossRef]
- Dong, D.; Xie, J.; Wang, J. Neuroprotective Effects of Brain-Gut Peptides: A Potential Therapy for Parkinson’s Disease. Neurosci. Bull. 2019, 35, 1085–1096. [Google Scholar] [CrossRef]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Gergs, U.; Jahn, T.; Werner, F.; Köhler, C.; Köpp, F.; Großmann, C.; Neumann, J. Overexpression of protein phosphatase 5 in the mouse heart: Reduced contractility but increased stress tolerance—Two sides of the same coin? PLoS ONE 2019, 14, e0221289. [Google Scholar] [CrossRef]
- Joyce, D.; Albanese, C.; Steer, J.; Fu, M.; Bouzahzah, B.; Pestell, R.G. NF-κB and cell-cycle regulation: The cyclin connection. Cytokine Growth Factor Rev. 2001, 12, 73–90. [Google Scholar] [CrossRef]
- Milder, J.; Patel, M. Modulation of oxidative stress and mitochondrial function by the ketogenic diet. Epilepsy Res. 2012, 100, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.C. Regulation of glutathione synthesis. Mol. Asp. Med. 2009, 30, 42–59. [Google Scholar] [CrossRef] [Green Version]
- Hanukoglu, I.; Feuchtwanger, R. Mechanism of corticotropin and cAMP induction of mitochondrial cytochrome P450 system enzymes in adrenal cortex cells. J. Biol. Chem. 1990, 265, 20602–20608. [Google Scholar] [CrossRef]
- Murataeva, N.; Daily, L.; Taylor, X.; Dhopeshwarkar, A.; Hu, S.S.-J.; Miller, S.; McHugh, D.; Oehler, O.; Li, S.; Bonanno, J.A.; et al. Evidence for a GPR18 Role in Chemotaxis, Proliferation, and the Course of Wound Closure in the Cornea. Cornea 2019, 38, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Ferro, R.; Adamska, A.; Lattanzio, R.; Mavrommati, I.; Edling, C.E.; Arifin, S.A.; Fyffe, C.A.; Sala, G.; Sacchetto, L.; Chiorino, G.; et al. GPR55 signalling promotes proliferation of pancreatic cancer cells and tumour growth in mice, and its inhibition increases effects of gemcitabine. Oncogene 2018, 37, 6368–6382. [Google Scholar] [CrossRef]
- Mosca, M.G.; Mangini, M.; Barba, P.; Mariggiò, S. Peptide Targeting of the Lysolipid-Sensing GPR55 for Osteoclastogenesis Tuning; In Review. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Li, Y.; Tweedie, D.; Mattson, M.P.; Holloway, H.W.; Greig, N.H. Enhancing the GLP-1 receptor signaling pathway leads to proliferation and neuroprotection in human neuroblastoma cells. J. Neurochem. 2010, 113, 1621–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hager, M.V.; Clydesdale, L.; Gellman, S.H.; Sexton, P.M.; Wootten, D. Characterization of signal bias at the GLP-1 receptor induced by backbone modification of GLP-1. Biochem. Pharmacol. 2017, 136, 99–108. [Google Scholar] [CrossRef]
- Xiong, S.; Xu, Y.; Ma, M.; Wang, H.; Wei, F.; Gu, Q.; Xu, X. Neuroprotective effects of a novel peptide, FK18, under oxygen-glucose deprivation in SH-SY5Y cells and retinal ischemia in rats via the Akt pathway. Neurochem. Int. 2017, 108, 78–90. [Google Scholar] [CrossRef]
- Brewer, J.R.; Mazot, P.; Soriano, P. Genetic insights into the mechanisms of Fgf signaling. Genes Dev. 2016, 30, 751–771. [Google Scholar] [CrossRef] [Green Version]
- Akhter, S.; Chakraborty, S.; Moutinho, D.; Álvarez-Coiradas, E.; Rosa, I.; Viñuela, J.; Domínguez, E.; García, A.; Requena, J.R. The human VGF-derived bioactive peptide TLQP-21 binds heat shock 71 kDa protein 8 (HSPA8)on the surface of SH-SY5Y cells. PLoS ONE 2017, 12, e0185176. [Google Scholar] [CrossRef] [PubMed]
- Pechan, P.M. Heat shock proteins and cell proliferation. FEBS Lett. 1991, 280, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Shevchenko, K.V.; Dulov, S.A.; Andreeva, L.A.; Nagaev, I.Y.; Shevchenko, V.P.; Radilov, A.S.; Myasoedov, N.F. Stability of His-Phe-Arg-Trp-Pro-Gly-Pro to Leucine Aminopeptidase, Carboxypeptidase Y, and Rat Nasal Mucus, Blood, and Plasma. Russ. J. Bioorg. Chem. 2016, 42, 153–161. [Google Scholar] [CrossRef]
- van de Loosdrecht, A.; Beelen, R.; Ossenkoppele, G.; Broekhoven, M.; Langenhuijsen, M. A tetrazolium-based colorimetric MTT assay to quantitate human monocyte mediated cytotoxicity against leukemic cells from cell lines and patients with acute myeloid leukemia. J. Immunol. Methods 1994, 174, 311–320. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akimov, M.G.; Fomina-Ageeva, E.V.; Dudina, P.V.; Andreeva, L.A.; Myasoyedov, N.F.; Bezuglov, V.V. ACTH(6–9)PGP Peptide Protects SH-SY5Y Cells from H2O2, tert-Butyl Hydroperoxide, and Cyanide Cytotoxicity via Stimulation of Proliferation and Induction of Prosurvival-Related Genes. Molecules 2021, 26, 1878. https://doi.org/10.3390/molecules26071878
Akimov MG, Fomina-Ageeva EV, Dudina PV, Andreeva LA, Myasoyedov NF, Bezuglov VV. ACTH(6–9)PGP Peptide Protects SH-SY5Y Cells from H2O2, tert-Butyl Hydroperoxide, and Cyanide Cytotoxicity via Stimulation of Proliferation and Induction of Prosurvival-Related Genes. Molecules. 2021; 26(7):1878. https://doi.org/10.3390/molecules26071878
Chicago/Turabian StyleAkimov, Mikhail G., Elena V. Fomina-Ageeva, Polina V. Dudina, Ludmila A. Andreeva, Nikolay F. Myasoyedov, and Vladimir V. Bezuglov. 2021. "ACTH(6–9)PGP Peptide Protects SH-SY5Y Cells from H2O2, tert-Butyl Hydroperoxide, and Cyanide Cytotoxicity via Stimulation of Proliferation and Induction of Prosurvival-Related Genes" Molecules 26, no. 7: 1878. https://doi.org/10.3390/molecules26071878
APA StyleAkimov, M. G., Fomina-Ageeva, E. V., Dudina, P. V., Andreeva, L. A., Myasoyedov, N. F., & Bezuglov, V. V. (2021). ACTH(6–9)PGP Peptide Protects SH-SY5Y Cells from H2O2, tert-Butyl Hydroperoxide, and Cyanide Cytotoxicity via Stimulation of Proliferation and Induction of Prosurvival-Related Genes. Molecules, 26(7), 1878. https://doi.org/10.3390/molecules26071878