
Combined Effects of Carotenoids and Polyphenols in Balancing the Response of Skin Cells to UV Irradiation


 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Carotenoids and Polyphenols Inhibit UVB-Induced Activation of the NFκB Transcription System
2.2. Cooperative Inhibition of UVB-Induced IL-6 Secretion by Combinations of Carotenoids and Polyphenols
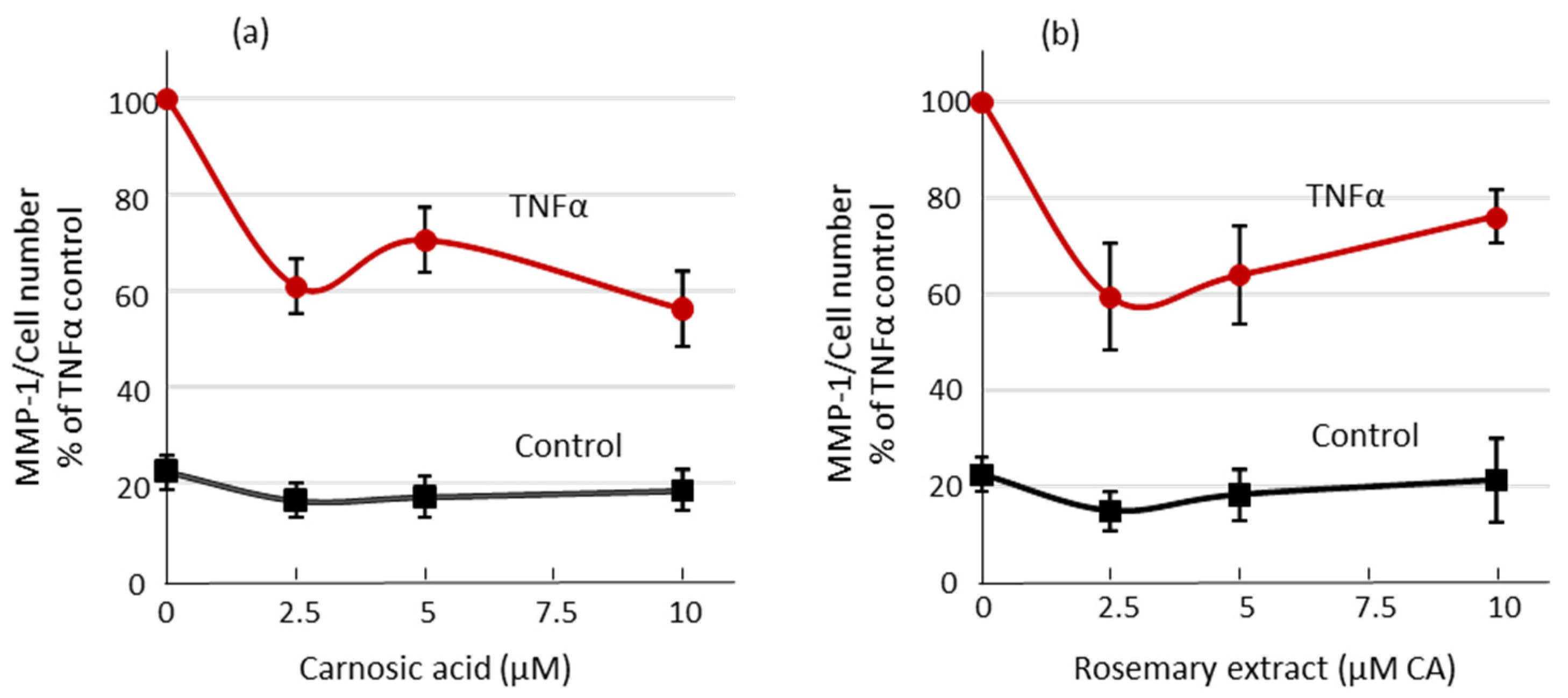
2.3. Matrix Metalloproteinase-1 (MMP-1) Secretion from Human Dermal Fibroblasts Is Reduced by Carnosic Acid Treatment
2.4. Synergistic Activation of the ARE/Nrf2 Transcription System in Skin Cells by Phytonutrient Combinations
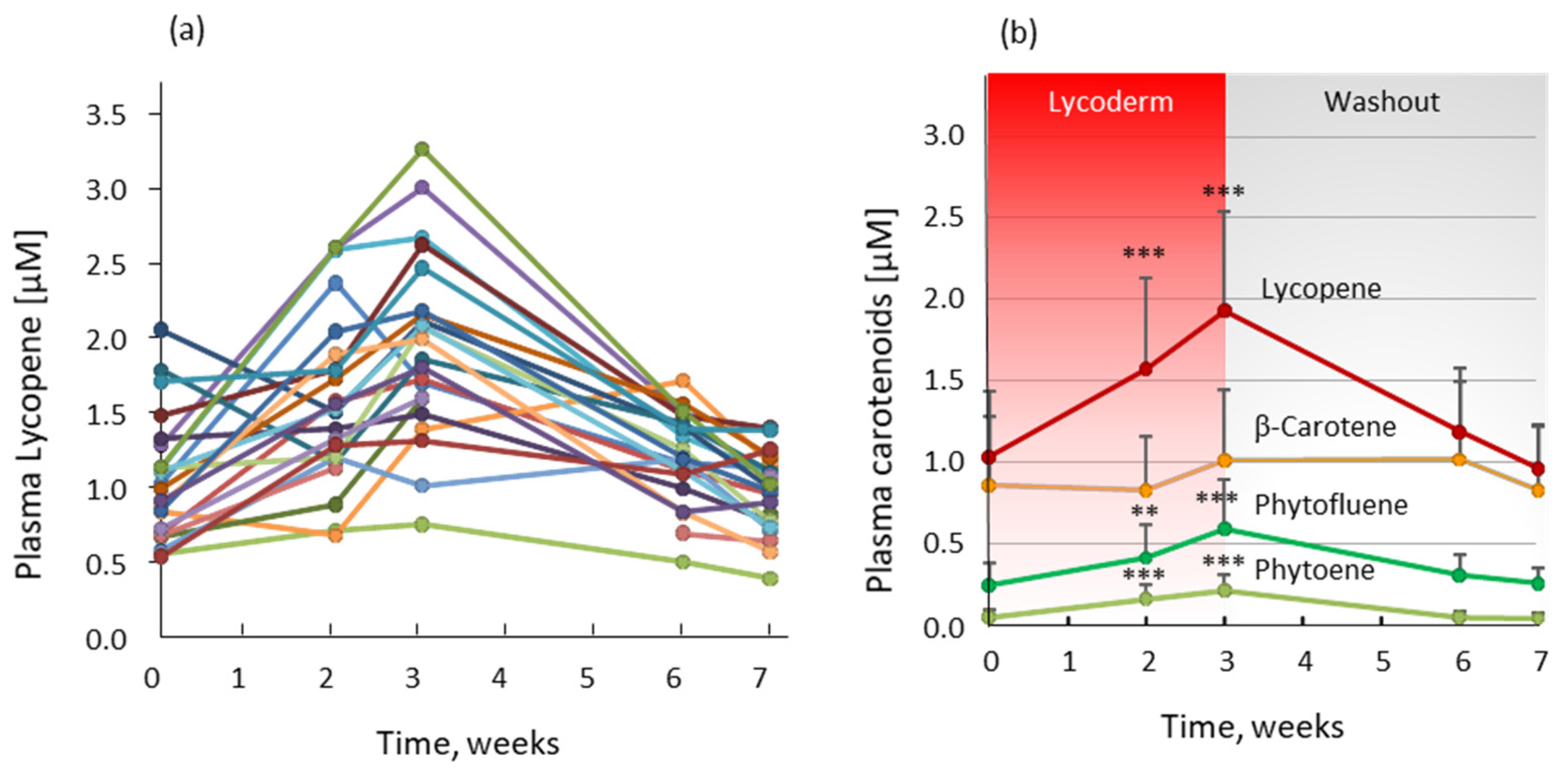
2.5. Bioavailability of Carotenoids from Lycored Tomato Nutrient Complex for Skin (LycodermTM)
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Treatment
4.3. Reporter Constructs
4.4. Transient Transfection and Reporter Gene Assay
4.5. Measurement of IL-6 Levels
4.6. Measurement of Matrix Metalloproteinase-1 (MMP-1)
4.7. Bioavailability of Carotenoid Supplements
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Aust, O.; Stahl, W.; Sies, H.; Tronnier, H.; Heinrich, U. Supplementation with tomato-based products increases lycopene, phytofluene, and phytoene levels in human serum and protects against UV-light-induced erythema. Int. J. Vitam. Nutr. Res. 2005, 75, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Greul, A.K.; Grundmann, J.U.; Heinrich, F.; Pfitzner, I.; Bernhardt, J.; Ambach, A.; Biesalski, H.K.; Gollnick, H. Photoprotection of UV-irradiated human skin: An antioxidative combination of vitamins E and C, carotenoids, selenium and proanthocyanidins. Skin Pharmacol. Appl. Skin Physiol. 2002, 15, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, U.; Gartner, C.; Wiebusch, M.; Eichler, O.; Sies, H.; Tronnier, H.; Stahl, W. Supplementation with beta-carotene or a similar amount of mixed carotenoids protects humans from UV-induced erythema. J. Nutr. 2003, 133, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Eicker, J.; Kurten, V.; Wild, S.; Riss, G.; Goralczyk, R.; Krutmann, J.; Berneburg, M. Betacarotene supplementation protects from photoaging-associated mitochondrial DNA mutation. Photochem. Photobiol. Sci. 2003, 2, 655–659. [Google Scholar] [CrossRef]
- Grether-Beck, S.; Marini, A.; Jaenicke, T.; Stahl, W.; Krutmann, J. Molecular evidence that oral supplementation with lycopene or lutein protects human skin against ultraviolet radiation: Results from a double-blinded, placebo-controlled, crossover study. Br. J Dermatol. 2017, 176, 1231–1240. [Google Scholar] [CrossRef]
- Kopcke, W.; Krutmann, J. Protection from sunburn with beta-Carotene--a meta-analysis. Photochem. Photobiol. 2008, 84, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Stahl, W. Nutritional protection against skin damage from sunlight. Annu. Rev. Nutr. 2004, 24, 173–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, U.; Moore, C.E.; De Spirt, S.; Tronnier, H.; Stahl, W. Green tea polyphenols provide photoprotection, increase microcirculation, and modulate skin properties of women. J. Nutr. 2011, 141, 1202–1208. [Google Scholar] [CrossRef] [Green Version]
- Eichler, O.; Sies, H.; Stahl, W. Divergent optimum levels of lycopene, beta-carotene and lutein protecting against UVB irradiation in human fibroblastst. Photochem. Photobiol. 2002, 75, 503–506. [Google Scholar] [CrossRef]
- Park, M.; Han, J.; Lee, C.S.; Soo, B.H.; Lim, K.M.; Ha, H. Carnosic acid, a phenolic diterpene from rosemary, prevents UV-induced expression of matrix metalloproteinases in human skin fibroblasts and keratinocytes. Exp. Dermatol. 2013, 22, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Sharoni, Y.; Linnewiel-Hermoni, K.; Zango, G.; Khanin, M.; Salman, H.; Veprik, A.; Danilenko, M.; Levy, J. The role of lycopene and its derivatives in the regulation of transcription systems: Implications for cancer prevention. Am. J. Clin. Nutr. 2012, 96, 1173S–1178S. [Google Scholar] [CrossRef] [PubMed]
- Abeyama, K.; Eng, W.; Jester, J.V.; Vink, A.A.; Edelbaum, D.; Cockerell, C.J.; Bergstresser, P.R.; Takashima, A. A role for NF-kappaB-dependent gene transactivation in sunburn. J. Clin. Investig. 2000, 105, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Wen, X.; Hao, D.; Zhang, N.; He, G.; Jiang, X. NF-κB signaling in skin aging. Mech. Ageing Dev. 2019, 184, 111160. [Google Scholar] [CrossRef]
- Cho, J.W.; Lee, K.S.; Kim, C.W. Curcumin attenuates the expression of IL-1beta, IL-6, and TNF-alpha as well as cyclin E in TNF-alpha-treated HaCaT cells; NF-kappaB and MAPKs as potential upstream targets. Int. J. Mol. Med. 2007, 19, 469–474. [Google Scholar] [PubMed]
- Yoshizumi, M.; Nakamura, T.; Kato, M.; Ishioka, T.; Kozawa, K.; Wakamatsu, K.; Kimura, H. Release of cytokines/chemokines and cell death in UVB-irradiated human keratinocytes, HaCaT. Cell Biol. Int. 2008, 32, 1405–1411. [Google Scholar] [CrossRef]
- Kim, J.K.; Mun, S.; Kim, M.S.; Kim, M.B.; Sa, B.K.; Hwang, J.K. 5,7-Dimethoxyflavone, an activator of PPARalpha/gamma, inhibits UVB-induced MMP expression in human skin fibroblast cells. Exp. Dermatol. 2012, 21, 211–216. [Google Scholar] [CrossRef]
- Lee, C.S.; Bae, I.H.; Han, J.; Choi, G.Y.; Hwang, K.H.; Kim, D.H.; Yeom, M.H.; Park, Y.H.; Park, M. Compound K inhibits MMP-1 expression through suppression of c-Src-dependent ERK activation in TNF-alpha-stimulated dermal fibroblast. Exp. Dermatol. 2014, 23, 819–824. [Google Scholar] [CrossRef]
- Oh, J.; Kim, J.H.; Park, J.G.; Yi, Y.S.; Park, K.W.; Rho, H.S.; Lee, M.S.; Yoo, J.W.; Kang, S.H.; Hong, Y.D.; et al. Radical scavenging activity-based and AP-1-targeted anti-inflammatory effects of lutein in macrophage-like and skin keratinocytic cells. Mediat. Inflamm. 2013, 2013, 787042. [Google Scholar] [CrossRef]
- Tanaka, K.; Asamitsu, K.; Uranishi, H.; Iddamalgoda, A.; Ito, K.; Kojima, H.; Okamoto, T. Protecting skin photoaging by NF-kappaB inhibitor. Curr. Drug. Metab. 2010, 11, 431–435. [Google Scholar] [CrossRef]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Talalay, P.; Dinkova-Kostova, A.T.; Holtzclaw, W.D. Importance of phase 2 gene regulation in protection against electrophile and reactive oxygen toxicity and carcinogenesis. Adv. Enzym. Regul. 2003, 43, 121–134. [Google Scholar] [CrossRef]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids activate the antioxidant response element transcription system. Mol. Cancer Ther. 2005, 4, 177–186. [Google Scholar]
- Adhami, V.M.; Afaq, F.; Ahmad, N. Suppression of ultraviolet B exposure-mediated activation of NF-kappaB in normal human keratinocytes by resveratrol. Neoplasia 2003, 5, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Saliou, C.; Kitazawa, M.; McLaughlin, L.; Yang, J.P.; Lodge, J.K.; Tetsuka, T.; Iwasaki, K.; Cillard, J.; Okamoto, T.; Packer, L. Antioxidants modulate acute solar ultraviolet radiation-induced NF-kappa-B activation in a human keratinocyte cell line. Free Radic. Biol. Med. 1999, 26, 174–183. [Google Scholar] [CrossRef]
- Ryu, J.; Kwon, M.J.; Nam, T.J. Nrf2 and NF-kappaB Signaling Pathways Contribute to Porphyra-334-Mediated Inhibition of UVA-Induced Inflammation in Skin Fibroblasts. Mar. Drugs 2015, 13, 4721–4732. [Google Scholar] [CrossRef] [Green Version]
- Linnewiel-Hermoni, K.; Motro, Y.; Miller, Y.; Levy, J.; Sharoni, Y. Carotenoid derivatives inhibit nuclear factor kappa B activity in bone and cancer cells by targeting key thiol groups. Free Radic. Biol. Med. 2014, 75, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Linnewiel, K.; Ernst, H.; Caris-Veyrat, C.; Ben-Dor, A.; Kampf, A.; Salman, H.; Danilenko, M.; Levy, J.; Sharoni, Y. Structure activity relationship of carotenoid derivatives in activation of the electrophile/antioxidant response element transcription system. Free Radic. Biol. Med. 2009, 47, 659–667. [Google Scholar] [CrossRef]
- Ohta, K.; Naruse, T.; Ishida, Y.; Shigeishi, H.; Nakagawa, T.; Fukui, A.; Nishi, H.; Sasaki, K.; Ogawa, I.; Takechi, M. TNF-alpha-induced IL-6 and MMP-9 expression in immortalized ameloblastoma cell line established by hTERT. Oral Dis. 2017, 23, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Kahari, V.M.; Saarialho-Kere, U. Matrix metalloproteinases in skin. Exp. Dermatol. 1997, 6, 199–213. [Google Scholar] [CrossRef]
- Odes-Barth, S.; Khanin, M.; Linnewiel-Hermoni, K.; Miller, Y.; Abramov, K.; Levy, J.; Sharoni, Y. Inhibition of Osteoclast Differentiation by Carotenoid Derivatives through Inhibition of the NF-ƙB Pathway. Antioxidants (Basel) 2020, 9, 1167. [Google Scholar] [CrossRef] [PubMed]
- Linnewiel-Hermoni, K.; Khanin, M.; Danilenko, M.; Zango, G.; Amosi, Y.; Levy, J.; Sharoni, Y. The anti-cancer effects of carotenoids and other phytonutrients resides in their combined activity. Arch. Biochem. Biophys. 2015, 572, 28–35. [Google Scholar] [CrossRef]
- De Jager, T.L.; Cockrell, A.E.; Du Plessis, S.S. Ultraviolet Light Induced Generation of Reactive Oxygen Species. In Ultraviolet Light in Human Health, Diseases and Environment; Ahmad, S.I., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 15–23. [Google Scholar] [CrossRef]
- Xian, D.; Xiong, X.; Xu, J.; Xian, L.; Lei, Q.; Song, J.; Zhong, J. Nrf2 Overexpression for the Protective Effect of Skin-Derived Precursors against UV-Induced Damage: Evidence from a Three-Dimensional Skin Model. Oxidative Med. Cell. Longev. 2019, 2019, 7021428. [Google Scholar] [CrossRef] [Green Version]
- Boo, Y.C. Natural Nrf2 Modulators for Skin Protection. Antioxidants (Basel Switzerland) 2020, 9, 812. [Google Scholar] [CrossRef] [PubMed]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroon, P.A.; Clifford, M.N.; Crozier, A.; Day, A.J.; Donovan, J.L.; Manach, C.; Williamson, G. How should we assess the effects of exposure to dietary polyphenols in vitro? Am. J. Clin. Nutr. 2004, 80, 15–21. [Google Scholar] [CrossRef]
- Brand, R.M.; Wipf, P.; Durham, A.; Epperly, M.W.; Greenberger, J.S.; Falo, L.D. Targeting Mitochondrial Oxidative Stress to Mitigate UV-Induced Skin Damage. Front. Pharmacol. 2018, 9, 920. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Yerra, V.G.; Negi, G.; Sharma, S.; Kumar, A. Potential therapeutic effects of the simultaneous targeting of the Nrf2 and NF-κB pathways in diabetic neuropathy. Redox Biol. 2013, 1, 394–397. [Google Scholar] [CrossRef] [Green Version]
- Hennig, P.; Fenini, G.; Di Filippo, M.; Beer, H.-D. Electrophiles against (Skin) Diseases: More Than Nrf2. Biomolecules 2020, 10, 271. [Google Scholar] [CrossRef] [Green Version]
- Brück, J.; Dringen, R.; Amasuno, A.; Pau-Charles, I.; Ghoreschi, K. A review of the mechanisms of action of dimethylfumarate in the treatment of psoriasis. Exp. Dermatol. 2018, 27, 611–624. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-E.; You, D.-J.; Lee, C.; Ahn, C.; Seong, J.Y.; Hwang, J.-I. Suppression of NF-κB signaling by KEAP1 regulation of IKKβ activity through autophagic degradation and inhibition of phosphorylation. Cell. Signal. 2010, 22, 1645–1654. [Google Scholar] [CrossRef]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [Green Version]
- Carroll, Y.L.; Corridan, B.M.; Morrissey, P.A. Lipoprotein carotenoid profiles and the susceptibility of low density lipoprotein to oxidative modification in healthy elderly volunteers. Eur. J. Clin. Nutr. 2000, 54, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Cooney, R.V.; Joseph Kappock, T.; Pung, A.; Bertram, J.S. Solubilization, cellular uptake, and activity of β-carotene and other carotenoids as inhibitors of neoplastic transformation in cultured cells. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1993; Volume 214, pp. 55–68. [Google Scholar]
- Walfisch, Y.; Walfisch, S.; Agbaria, R.; Levy, J.; Sharoni, Y. Lycopene in serum, skin and adipose tissues after tomato-oleoresin supplementation in patients undergoing haemorrhoidectomy or peri-anal fistulotomy. Br. J. Nutr. 2003, 90, 759–766. [Google Scholar] [CrossRef] [Green Version]
- Tarshish, E.; Hermoni, K.; Schwartz, S.R. Effect of Oral Supplement “Lycopene” On Reducing the Signs of Skin Ageing. Clin. Pharmacol. Biopharm. 2020, 9, 195. [Google Scholar] [CrossRef]
- Groten, K.; Marini, A.; Grether-Beck, S.; Jaenicke, T.; Ibbotson, S.H.; Moseley, H.; Ferguson, J.; Krutmann, J. Tomato Phytonutrients Balance UV Response: Results from a Double-Blind, Randomized, Placebo-Controlled Study. Skin Pharmacol. Physiol. 2019, 32, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.; Bosin, E.; Feldman, B.; Giat, Y.; Miinster, A.; Danilenko, M.; Sharoni, Y. Lycopene is a more potent inhibitor of human cancer cell proliferation than either alpha-carotene or beta-carotene. Nutr. Cancer 1995, 24, 257–266. [Google Scholar] [CrossRef]
- Stahl, W.; Sundquist, A.R.; Hanusch, M.; Schwarz, W.; Sies, H. Separation of beta-carotene and lycopene geometrical isomers in biological samples. Clin. Chem. 1993, 39, 810–814. [Google Scholar] [CrossRef] [PubMed]

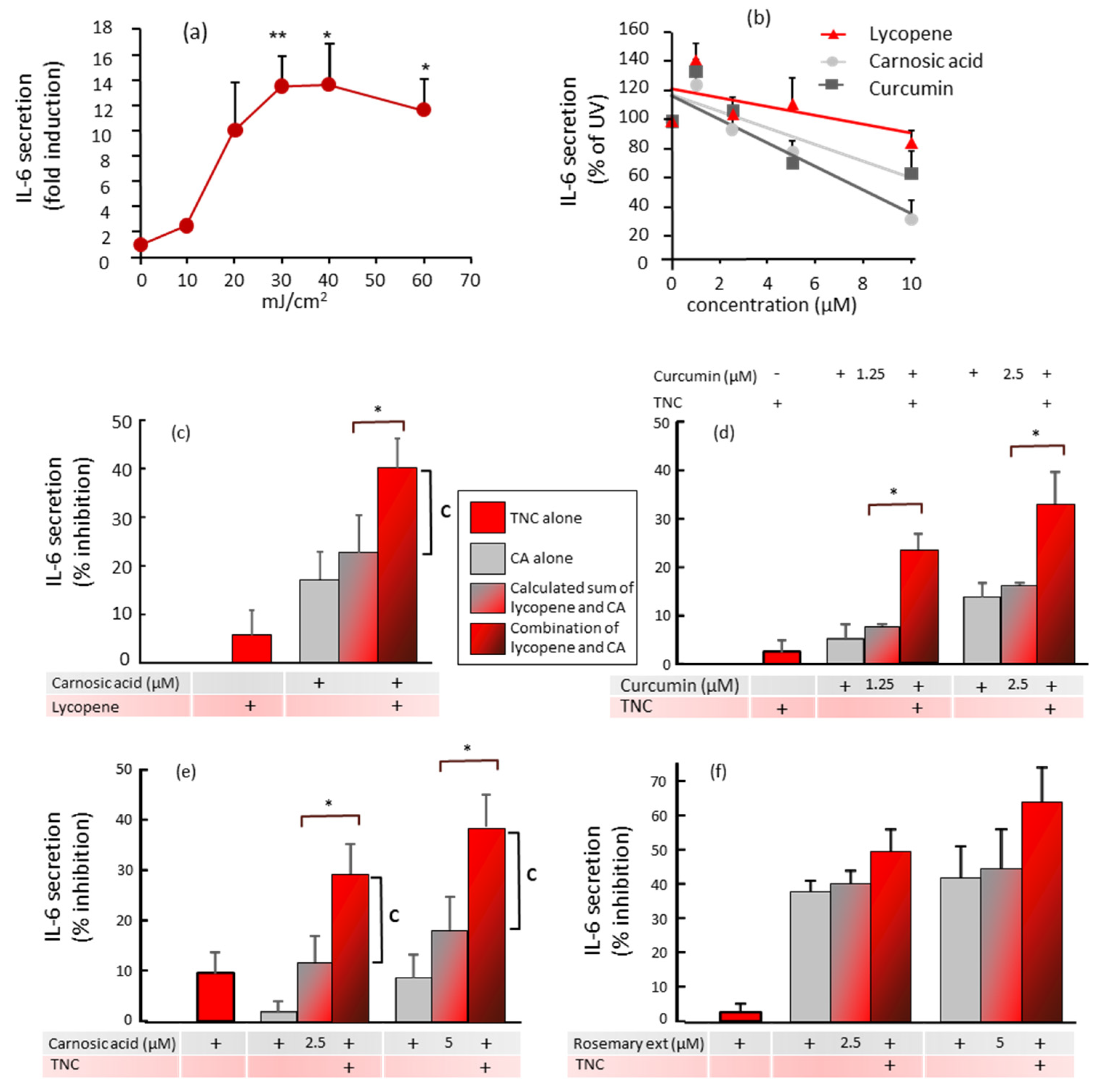
 ) and compared to the effects of the combinations of polyphenols and carotenoids (
) and compared to the effects of the combinations of polyphenols and carotenoids ( ). * p < 0.05 indicates that the combination is significantly different from the sum of each compound alone. The vertical line marked “C” indicates the percentage inhibition by the combination that is above the calculated sum of the percentage inhibition by the separate compounds. (c) Values are the means ± SEM of 4–7 experiments. Concentrations were: lycopene 5 µM and carnosic acid 10 µM; (d) Values are the means ± SEM of two experiments done with the indicated concentrations of curcumin. Concentrations of lycopene from the Tomato Nutrient Complex (TNC) were 10 µM; (e) Values are the means ± SEM of four experiments done with the indicated concentrations of carnosic acid. Concentrations of lycopene from the TNC were 10 µM; (f) Values are the means ± SEM of two experiments done with the indicated concentrations of carnosic acid from the rosemary extract. Concentrations of lycopene from the TNC were 10 µM.
) and compared to the effects of the combinations of polyphenols and carotenoids (). * p < 0.05 indicates that the combination is significantly different from the sum of each compound alone. The vertical line marked “C” indicates the percentage inhibition by the combination that is above the calculated sum of the percentage inhibition by the separate compounds. (c) Values are the means ± SEM of 4–7 experiments. Concentrations were: lycopene 5 µM and carnosic acid 10 µM; (d) Values are the means ± SEM of two experiments done with the indicated concentrations of curcumin. Concentrations of lycopene from the Tomato Nutrient Complex (TNC) were 10 µM; (e) Values are the means ± SEM of four experiments done with the indicated concentrations of carnosic acid. Concentrations of lycopene from the TNC were 10 µM; (f) Values are the means ± SEM of two experiments done with the indicated concentrations of carnosic acid from the rosemary extract. Concentrations of lycopene from the TNC were 10 µM.
). * p < 0.05 indicates that the combination is significantly different from the sum of each compound alone. The vertical line marked “C” indicates the percentage inhibition by the combination that is above the calculated sum of the percentage inhibition by the separate compounds. (c) Values are the means ± SEM of 4–7 experiments. Concentrations were: lycopene 5 µM and carnosic acid 10 µM; (d) Values are the means ± SEM of two experiments done with the indicated concentrations of curcumin. Concentrations of lycopene from the Tomato Nutrient Complex (TNC) were 10 µM; (e) Values are the means ± SEM of four experiments done with the indicated concentrations of carnosic acid. Concentrations of lycopene from the TNC were 10 µM; (f) Values are the means ± SEM of two experiments done with the indicated concentrations of carnosic acid from the rosemary extract. Concentrations of lycopene from the TNC were 10 µM.
) and compared to the effects of the combinations of polyphenols and carotenoids (). * p < 0.05 indicates that the combination is significantly different from the sum of each compound alone. The vertical line marked “C” indicates the percentage inhibition by the combination that is above the calculated sum of the percentage inhibition by the separate compounds. (c) Values are the means ± SEM of 4–7 experiments. Concentrations were: lycopene 5 µM and carnosic acid 10 µM; (d) Values are the means ± SEM of two experiments done with the indicated concentrations of curcumin. Concentrations of lycopene from the Tomato Nutrient Complex (TNC) were 10 µM; (e) Values are the means ± SEM of four experiments done with the indicated concentrations of carnosic acid. Concentrations of lycopene from the TNC were 10 µM; (f) Values are the means ± SEM of two experiments done with the indicated concentrations of carnosic acid from the rosemary extract. Concentrations of lycopene from the TNC were 10 µM.

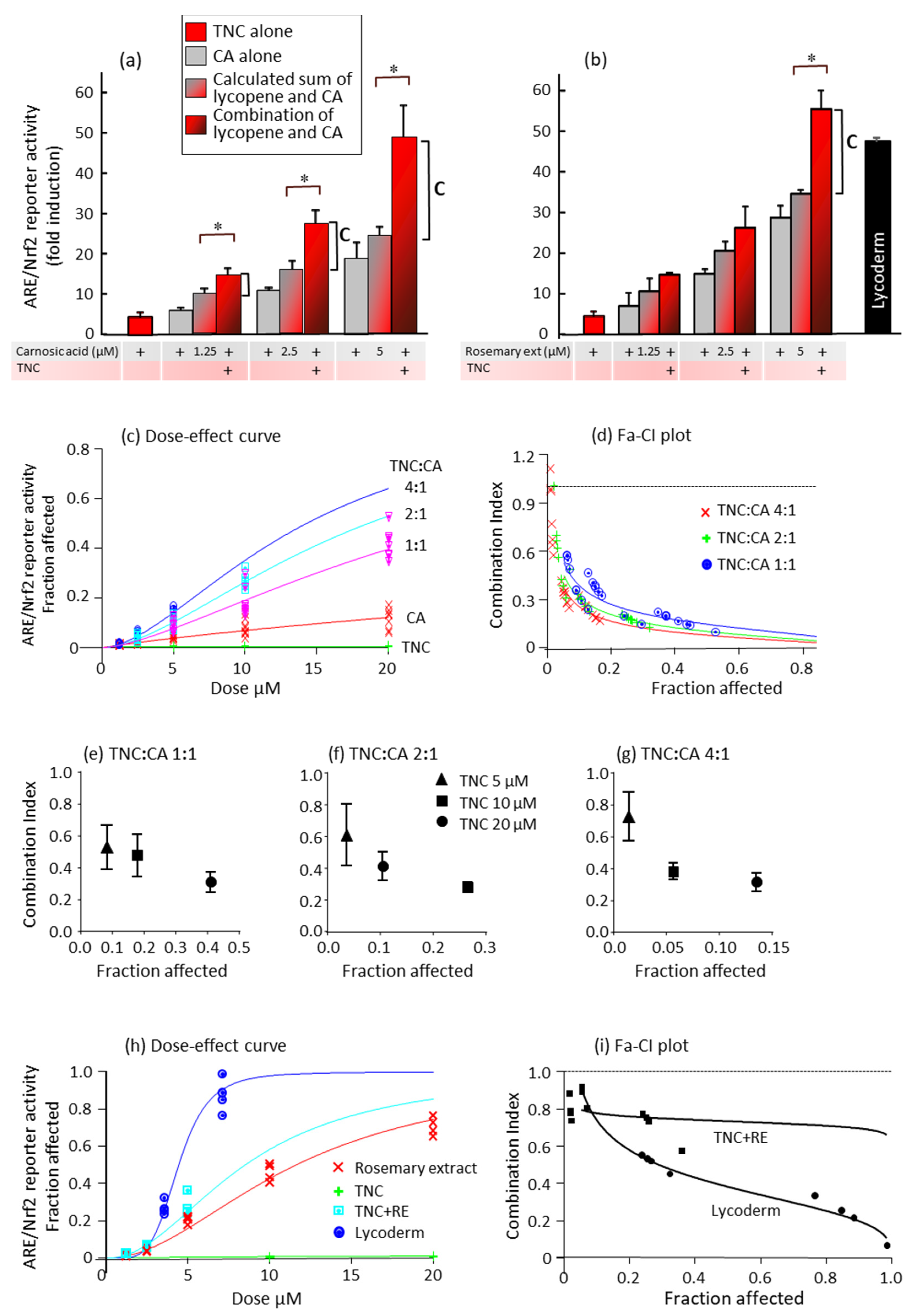
 ) and compared to the effects of the combinations of polyphenols and carotenoids (
) and compared to the effects of the combinations of polyphenols and carotenoids ( ). * p < 0.05 indicates that the combination is significantly different from the sum of each compound alone. The vertical line marked “C” indicates ARE/Nrf2 induction by the combination that is above the calculated sum of the induction by the separate compounds. (c–g) Combination index calculations of the synergy between TNC and carnosic acid. Different concentrations of TNC, carnosic acid, or their combinations at the ratios indicated in the figure were added 6 h after transfection. (c) Dose-effect curve at concentrations of 1.25–20 µM and at TNC:carnosic acid ratios of 1:1, 2:1, and 4:1. The µM concentrations shown on the x-axis are of each of the compounds alone or of carnosic acid in the combinations. Results are from two experiments, each done in triplicate. (d) Fraction affected-combination index plots of the data presented in (c). (e–g) Average CI values for the TNC–carnosic acid combinations. (h,i) Different concentrations of lycopene in TNC, carnosic acid in rosemary extract, or their combinations at a ratio of 4:1 were added 6 h after transfection. (h) Dose-effect curve at concentrations of 1.25–20 µM and at TNC:carnosic acid ratios of 4:1. The µM concentrations shown on the x-axis are of each of the compounds alone or of carnosic acid (from rosemary extract) in the combinations. LycodermTM containing lycopene:carnosic acid at a ratio of 1.8:1 was added at the indicated concentrations of carnosic acid. (i) Fraction affected-combination index plot of the data presented in (h).
) and compared to the effects of the combinations of polyphenols and carotenoids (). * p < 0.05 indicates that the combination is significantly different from the sum of each compound alone. The vertical line marked “C” indicates ARE/Nrf2 induction by the combination that is above the calculated sum of the induction by the separate compounds. (c–g) Combination index calculations of the synergy between TNC and carnosic acid. Different concentrations of TNC, carnosic acid, or their combinations at the ratios indicated in the figure were added 6 h after transfection. (c) Dose-effect curve at concentrations of 1.25–20 µM and at TNC:carnosic acid ratios of 1:1, 2:1, and 4:1. The µM concentrations shown on the x-axis are of each of the compounds alone or of carnosic acid in the combinations. Results are from two experiments, each done in triplicate. (d) Fraction affected-combination index plots of the data presented in (c). (e–g) Average CI values for the TNC–carnosic acid combinations. (h,i) Different concentrations of lycopene in TNC, carnosic acid in rosemary extract, or their combinations at a ratio of 4:1 were added 6 h after transfection. (h) Dose-effect curve at concentrations of 1.25–20 µM and at TNC:carnosic acid ratios of 4:1. The µM concentrations shown on the x-axis are of each of the compounds alone or of carnosic acid (from rosemary extract) in the combinations. LycodermTM containing lycopene:carnosic acid at a ratio of 1.8:1 was added at the indicated concentrations of carnosic acid. (i) Fraction affected-combination index plot of the data presented in (h).
). * p < 0.05 indicates that the combination is significantly different from the sum of each compound alone. The vertical line marked “C” indicates ARE/Nrf2 induction by the combination that is above the calculated sum of the induction by the separate compounds. (c–g) Combination index calculations of the synergy between TNC and carnosic acid. Different concentrations of TNC, carnosic acid, or their combinations at the ratios indicated in the figure were added 6 h after transfection. (c) Dose-effect curve at concentrations of 1.25–20 µM and at TNC:carnosic acid ratios of 1:1, 2:1, and 4:1. The µM concentrations shown on the x-axis are of each of the compounds alone or of carnosic acid in the combinations. Results are from two experiments, each done in triplicate. (d) Fraction affected-combination index plots of the data presented in (c). (e–g) Average CI values for the TNC–carnosic acid combinations. (h,i) Different concentrations of lycopene in TNC, carnosic acid in rosemary extract, or their combinations at a ratio of 4:1 were added 6 h after transfection. (h) Dose-effect curve at concentrations of 1.25–20 µM and at TNC:carnosic acid ratios of 4:1. The µM concentrations shown on the x-axis are of each of the compounds alone or of carnosic acid (from rosemary extract) in the combinations. LycodermTM containing lycopene:carnosic acid at a ratio of 1.8:1 was added at the indicated concentrations of carnosic acid. (i) Fraction affected-combination index plot of the data presented in (h).
) and compared to the effects of the combinations of polyphenols and carotenoids (). * p < 0.05 indicates that the combination is significantly different from the sum of each compound alone. The vertical line marked “C” indicates ARE/Nrf2 induction by the combination that is above the calculated sum of the induction by the separate compounds. (c–g) Combination index calculations of the synergy between TNC and carnosic acid. Different concentrations of TNC, carnosic acid, or their combinations at the ratios indicated in the figure were added 6 h after transfection. (c) Dose-effect curve at concentrations of 1.25–20 µM and at TNC:carnosic acid ratios of 1:1, 2:1, and 4:1. The µM concentrations shown on the x-axis are of each of the compounds alone or of carnosic acid in the combinations. Results are from two experiments, each done in triplicate. (d) Fraction affected-combination index plots of the data presented in (c). (e–g) Average CI values for the TNC–carnosic acid combinations. (h,i) Different concentrations of lycopene in TNC, carnosic acid in rosemary extract, or their combinations at a ratio of 4:1 were added 6 h after transfection. (h) Dose-effect curve at concentrations of 1.25–20 µM and at TNC:carnosic acid ratios of 4:1. The µM concentrations shown on the x-axis are of each of the compounds alone or of carnosic acid (from rosemary extract) in the combinations. LycodermTM containing lycopene:carnosic acid at a ratio of 1.8:1 was added at the indicated concentrations of carnosic acid. (i) Fraction affected-combination index plot of the data presented in (h).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calniquer, G.; Khanin, M.; Ovadia, H.; Linnewiel-Hermoni, K.; Stepensky, D.; Trachtenberg, A.; Sedlov, T.; Braverman, O.; Levy, J.; Sharoni, Y. Combined Effects of Carotenoids and Polyphenols in Balancing the Response of Skin Cells to UV Irradiation. Molecules 2021, 26, 1931. https://doi.org/10.3390/molecules26071931
Calniquer G, Khanin M, Ovadia H, Linnewiel-Hermoni K, Stepensky D, Trachtenberg A, Sedlov T, Braverman O, Levy J, Sharoni Y. Combined Effects of Carotenoids and Polyphenols in Balancing the Response of Skin Cells to UV Irradiation. Molecules. 2021; 26(7):1931. https://doi.org/10.3390/molecules26071931
Chicago/Turabian StyleCalniquer, Glenda, Marina Khanin, Hilla Ovadia, Karin Linnewiel-Hermoni, David Stepensky, Aviram Trachtenberg, Tanya Sedlov, Oleg Braverman, Joseph Levy, and Yoav Sharoni. 2021. "Combined Effects of Carotenoids and Polyphenols in Balancing the Response of Skin Cells to UV Irradiation" Molecules 26, no. 7: 1931. https://doi.org/10.3390/molecules26071931
APA StyleCalniquer, G., Khanin, M., Ovadia, H., Linnewiel-Hermoni, K., Stepensky, D., Trachtenberg, A., Sedlov, T., Braverman, O., Levy, J., & Sharoni, Y. (2021). Combined Effects of Carotenoids and Polyphenols in Balancing the Response of Skin Cells to UV Irradiation. Molecules, 26(7), 1931. https://doi.org/10.3390/molecules26071931