
In Vitro Inhibitory Analysis of Rationally Designed siRNAs against MERS-CoV Replication in Huh7 Cells

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Sequence Isolation and Multiple Sequence Analysis
2.2. Prediction and Selection of Potential siRNAs
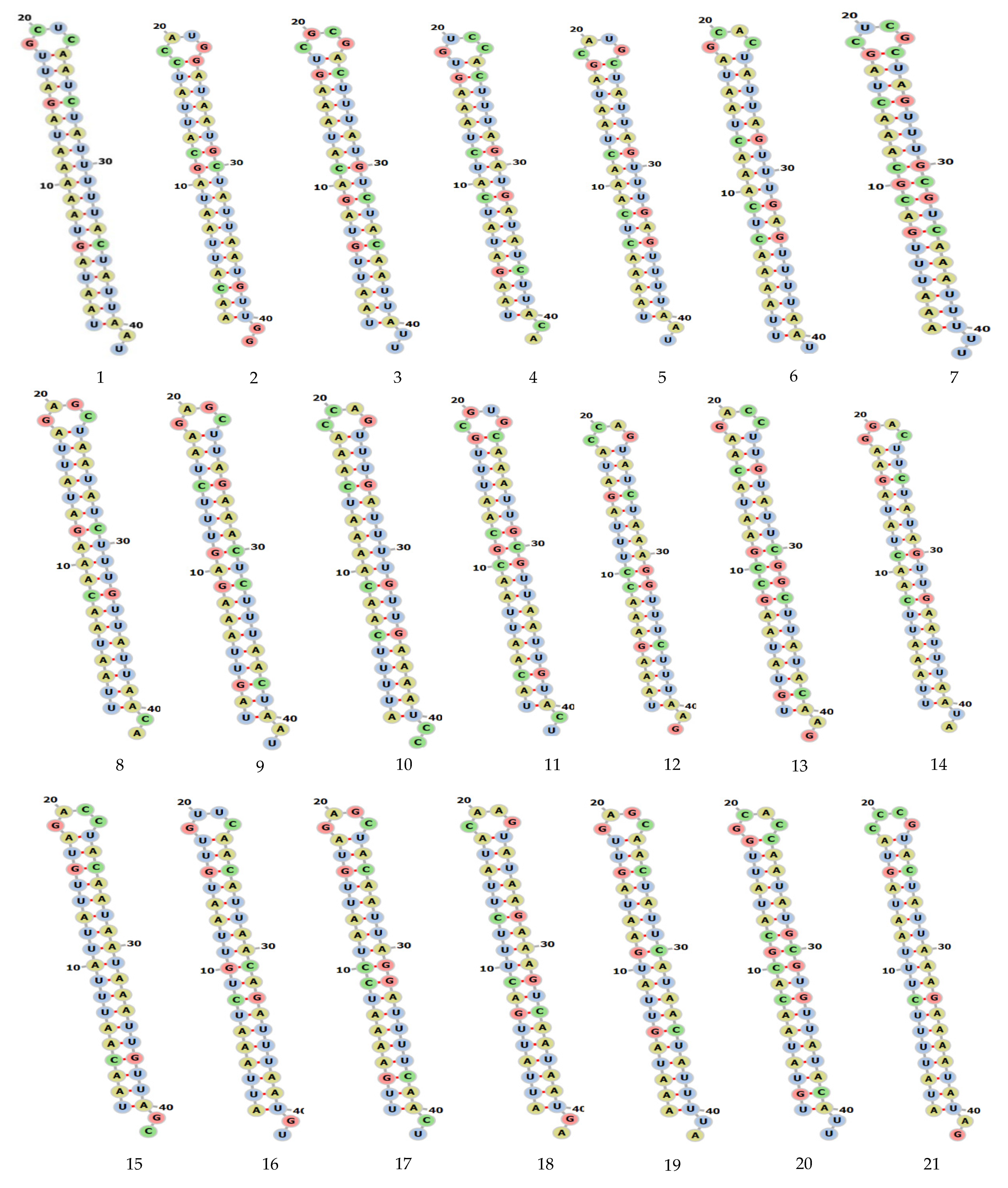
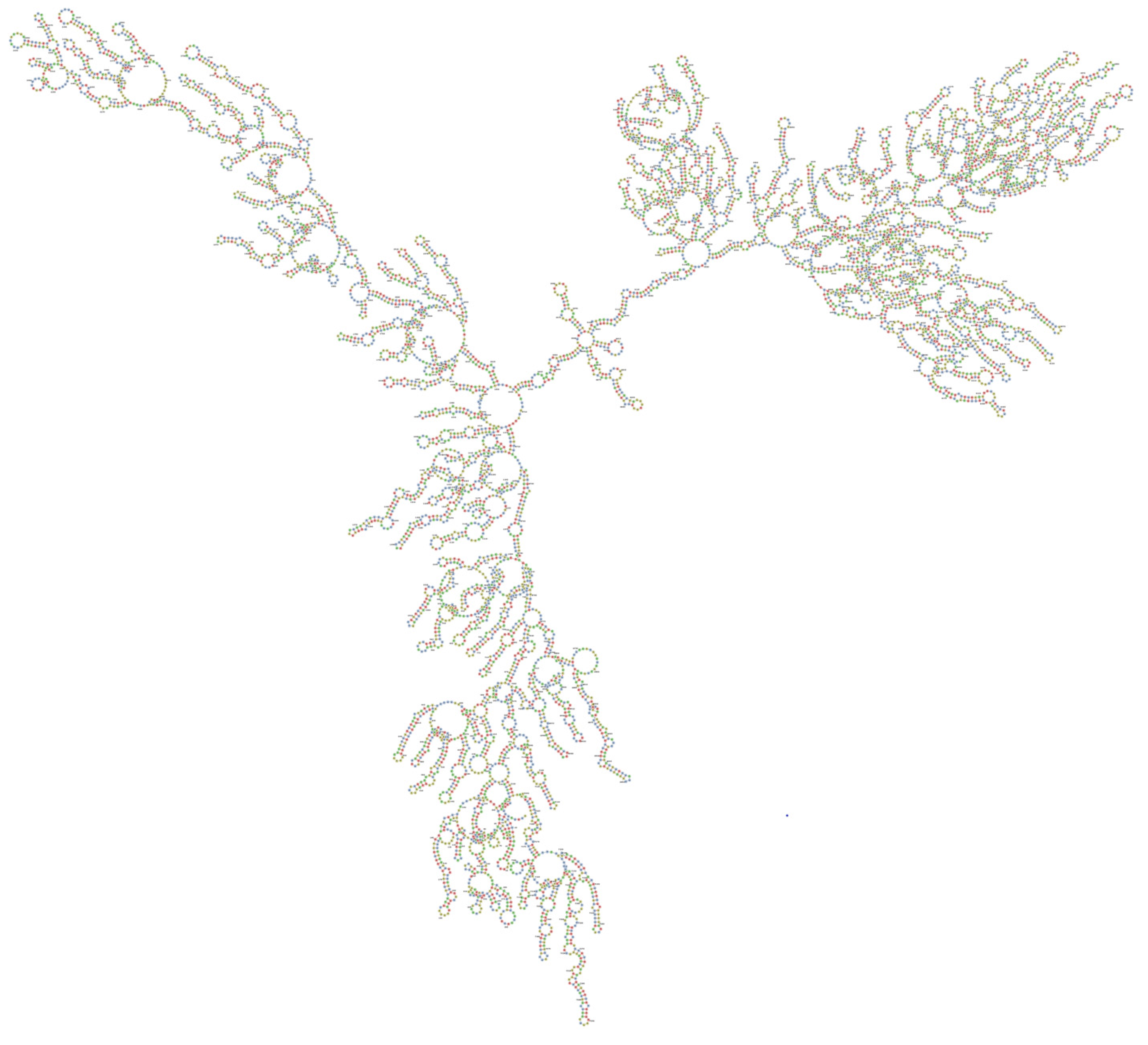
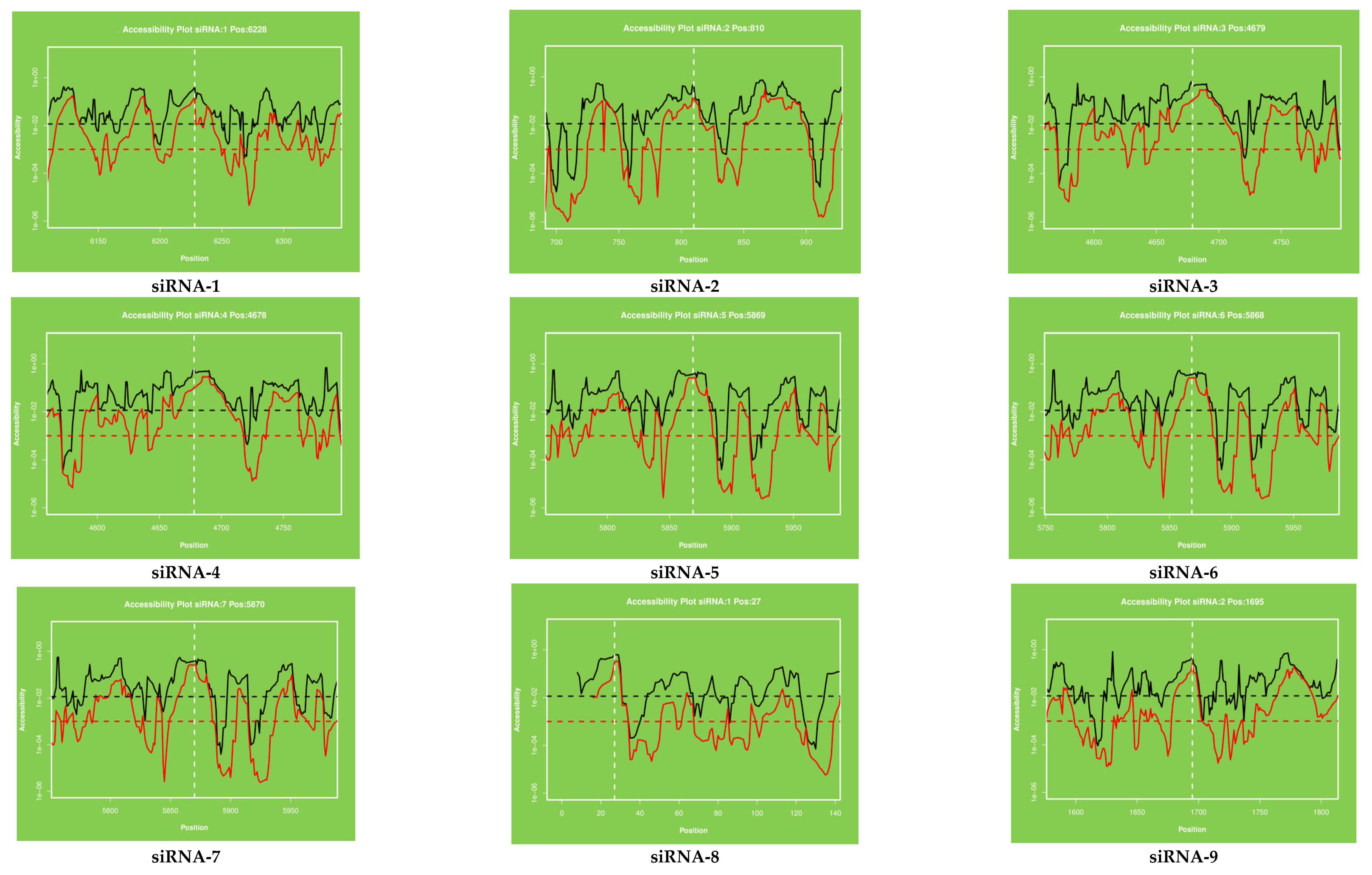
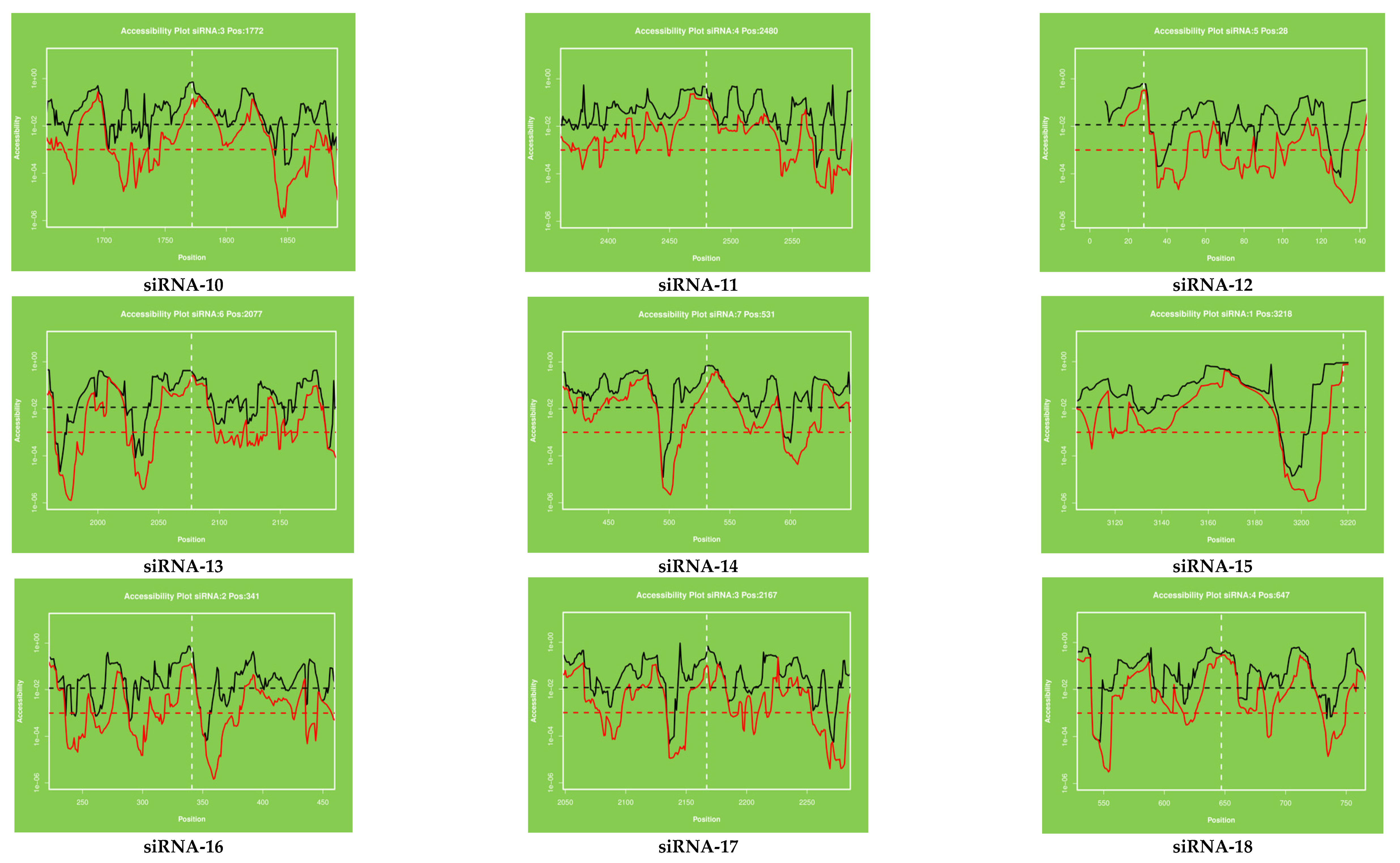
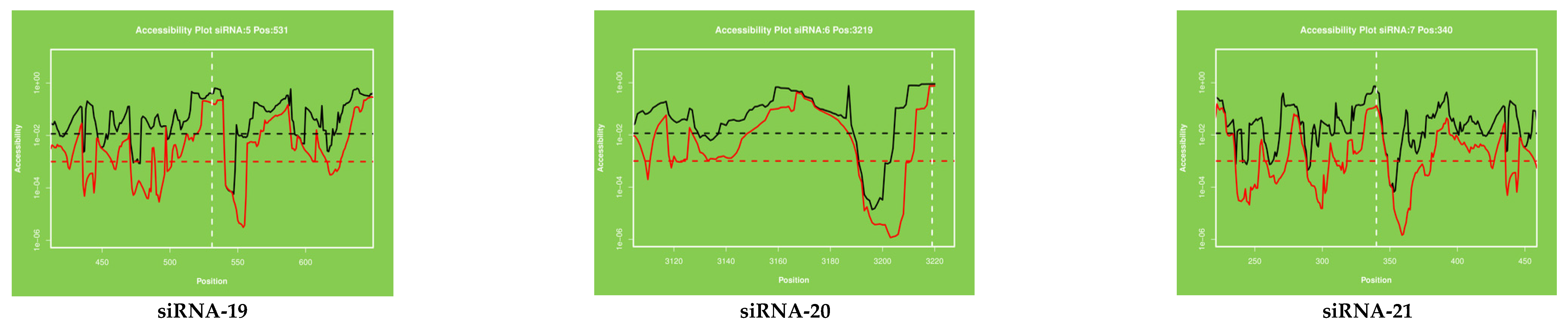
2.3. Target Accessibility and Secondary Structure Prediction
2.4. siRNA Transfection and Cytotoxicity Assay
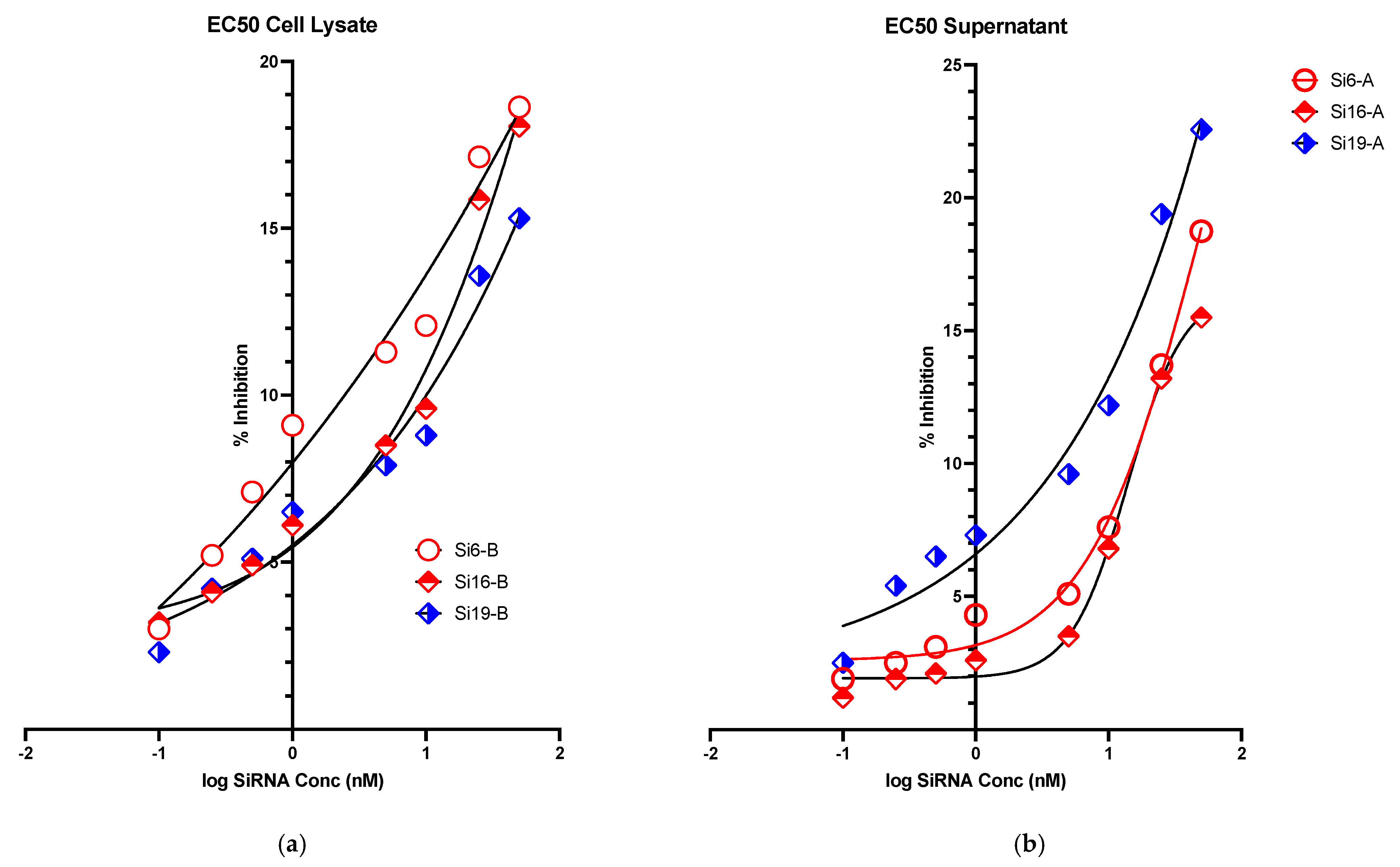
2.5. Evaluation of Virus Replication Inhibition
3. Discussion
4. Materials and Methods
4.1. Sequence Selection and Multiple Sequence Analysis
4.2. In Silico Prediction, Selection and Synthesis of siRNAs
4.3. Secondary Structure and Target Accessibility Prediction
4.4. Transfection of siRNA to Huh7 Cells
4.5. Cytotoxicity Assay
4.6. Analysis of MERS-CoV Replication Inhibition in Huh7 Cells
4.7. Inoculation of MERS-CoV to Huh7 Cells
4.8. Real-Time PCR Assay
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Middle East Respiratory Syndrome Coronavirus (MERS-CoV). Available online: http://www.emro.who.int/pandemic-epidemic-diseases/mers-cov/mers-situation-update-january-2020.html (accessed on 25 February 2021).
- Assiri, A.; Al-Tawfiq, J.A.; Al-Rabeeah, A.A.; Al-Rabiah, F.A.; Al-Hajjar, S.; Al-Barrak, A.; Flemban, H.; Al-Nassir, W.N.; Balkhy, H.H.; Al-Hakeem, R.F. Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respira-tory syndrome coronavirus disease from Saudi Arabia: A descriptive study. Lancet Infect. Dis. 2013, 13, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wunderink, R.G. MERS, SARS and other coronaviruses as causes of pneumonia. Respirology 2017, 23, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S.; Wong, N.S. Probable transmission chains of Middle East respiratory syndrome coronavirus and the multiple generations of secondary infection in South Korea. Int. J. Infect. Dis. 2015, 38, 65–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for cam-el-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Oboho, I.K.; Tomczyk, S.M.; Al-Asmari, A.M.; Banjar, A.A.; Al-Mugti, H.; Aloraini, M.S.; Alkhaldi, K.Z.; Almohammadi, E.L.; Alraddadi, B.M.; Gerber, S.I.; et al. 2014 MERS-CoV outbreak in Jeddah--a link to health care facilities. N. Engl. J. Med. 2015, 372, 846–854. [Google Scholar] [CrossRef]
- Memish, Z.A.; Cotten, M.; Watson, S.J.; Kellam, P.; Zumla, A.; Alhakeem, R.F.; Assiri, A.; Al Rabeeah, A.A.; Al-Tawfiq, J.A. Community Case Clusters of Middle East Respiratory Syndrome Coronavirus in Hafr Al-Batin, Kingdom of Saudi Arabia: A Descriptive Genomic study. Int. J. Infect. Dis. 2014, 23, 63–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyong’a, A.N.; Cook, E.A.; Okba, N.; Kivali, V.; Reusken, C.; Haagmans, B.L.; Fèvre, E.M. Middle East respiratory syndrome coronavirus (MERS-CoV) seropositive camel handlers in Kenya. Viruses 2020, 12, 396. [Google Scholar] [CrossRef] [Green Version]
- Al Awaidy, S.T.; Khamis, F. Middle east respiratory syndrome coronavirus (Mers-cov) in Oman: Current situation and going forward. Oman Med. J. 2019, 34, 181. [Google Scholar] [CrossRef] [PubMed]
- Chafekar, A.; Fielding, B.C. MERS-CoV: Understanding the latest human coronavirus threat. Viruses 2018, 10, 93. [Google Scholar] [CrossRef]
- Al-Omari, A.; Rabaan, A.A.; Salih, S.; Al-Tawfiq, J.A.; Memish, Z.A. MERS coronavirus outbreak: Implications for emerging viral infections. Diagn. Microbiol. Infect. Dis. 2019, 93, 265–285. [Google Scholar] [CrossRef]
- Corman, V.M.; Ithete, N.L.; Richards, L.R.; Schoeman, M.C.; Preiser, W.; Drosten, C.; Drexler, J.F.; Raposo, R.A.S.; Abdel-Mohsen, M.; Deng, X.; et al. Rooting the Phylogenetic Tree of Middle East Respiratory Syndrome Coronavirus by Characterization of a Conspecific Virus from an African Bat. J. Virol. 2014, 88, 11297–11303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mubarak, A.; Alturaiki, W.; Hemida, M.G. Middle East Respiratory Syndrome Coronavirus (MERS-CoV): Infection, Immunological Response, and Vaccine Development. J. Immunol. Res. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yong, C.Y.; Ong, H.K.; Yeap, S.K.; Ho, K.L.; Tan, W.S. Recent Advances in the Vaccine Development Against Middle East Respiratory Syndrome-Coronavirus. Front. Microbiol. 2019, 10, 1781. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-D.; Chi, W.-Y.; Su, J.-H.; Ferrall, L.; Hung, C.-F.; Wu, T.-C. Coronavirus vaccine development: From SARS and MERS to COVID-19. J. Biomed. Sci. 2020, 27, 1–23. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.-S. Therapeutic miRNA and siRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol. Ther. Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levanova, A.; Poranen, M.M. RNA Interference as a Prospective Tool for the Control of Human Viral Infections. Front. Microbiol. 2018, 9, 2151. [Google Scholar] [CrossRef] [PubMed]
- Sohrab, S.S.; El-Kafrawy, S.A.; Mirza, Z.; Kamal, M.A.; Azhar, E.I. Design and Delivery of Therapeutic siRNAs: Application to MERS-Coronavirus. Curr. Pharm. Des. 2018, 24, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Setten, R.L.; Rossi, J.J.; Han, S.-P. The current state and future directions of RNAi-based therapeutics. Nat. Rev. Drug Discov. 2019, 18, 421–446. [Google Scholar] [CrossRef]
- Liu, C.; Liang, Z.; Kong, X. Efficacy Analysis of Combinatorial siRNAs against HIV Derived from One Double Hairpin RNA Precursor. Front. Microbiol. 2017, 8, 1651. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Christiaens, O.; Li, X.; Swevers, L.; Casteels, H.; Maes, M.; Smagghe, G. Engineered Flock House Virus for Targeted Gene Suppression Through RNAi in Fruit Flies (Drosophila melanogaster) in Vitro and in Vivo. Front. Physiol. 2018, 9, 805. [Google Scholar] [CrossRef] [Green Version]
- Idrees, S.; A Ashfaq, U. RNAi: Antiviral therapy against dengue virus. Asian Pac. J. Trop. Biomed. 2013, 3, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Shahid, I.; AlRabia, M.W.; Mukhtar, M.H.; Almalki, S.S.R.; Alkahtani, S.A.; Ashgar, S.S.; Faidah, H.S.; Hafeez, M.H.; Almalki, W.H. In vitro inhibitory analysis of consensus siRNAs against NS3 gene of hepatitis C virus 1a genotype. Asian Pac. J. Trop. Med. 2017, 10, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Szabat, M.; Lorent, D.; Czapik, T.; Tomaszewska, M.; Kierzek, E.; Kierzek, R. RNA Secondary Structure as a First Step for Rational Design of the Oligonucleotides towards Inhibition of Influenza A Virus Replication. Pathogens 2020, 9, 925. [Google Scholar] [CrossRef]
- Huang, D.T.-N.; Lu, C.-Y.; Shao, P.-L.; Chang, L.-Y.; Wang, J.-Y.; Chang, Y.-H.; Lai, M.-J.; Chi, Y.-H.; Huang, L.-M. In vivo inhibition of influenza A virus replication by RNA interference targeting the PB2 subunit via intratracheal delivery. PLoS ONE 2017, 12, e0174523. [Google Scholar] [CrossRef]
- Tsai, K.; Courtney, D.G.; Kennedy, E.M.; Cullen, B.R. Influenza A virus-derived siRNAs increase in the absence of NS1 yet fail to inhibit virus replication. RNA 2018, 24, 1172–1182. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Yang, L.; Zuo, J.-P. Recent developments in antivirals against hepatitis B virus. Virus Res. 2016, 213, 205–213. [Google Scholar] [CrossRef]
- Zeng, J.; Yang, S.; Wang, X.; Gao, Y.; Zhang, M. Effects of siRNA-mediated suppression of HPV-11 L1 expression on the proliferation and apoptosis of vaginal epithelial cells. Exp. Ther. Med. 2017, 13, 1561–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Jung, Y.-S.; Liang, P.-H. Anti-SARS coronavirus agents: A patent review (2008–present). Expert Opin. Ther. Patents 2013, 23, 1337–1348. [Google Scholar] [CrossRef] [Green Version]
- Sohrab, S.S.; El-Kafrawy, S.A.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. Designing and evaluation of MERS-CoV siRNAs in HEK-293 Cell line. J. Infect. Public Health 2020, 4, 238–243. [Google Scholar] [CrossRef]
- Chowdhury, U.F.; Shohan, M.U.S.; Hoque, K.I.; Beg, M.A.; Siam, M.K.S.; Moni, M.A. A computational approach to design potential siRNA molecules as a prospective tool for silencing nucleocapsid phosphoprotein and surface glycoprotein gene of SARS-CoV-2. Genomics 2020, 113, 331–343. [Google Scholar] [CrossRef]
- Sohrab, S.S.; El-Kafrawy, S.A.; Abbas, A.T.; Bajrai, L.H.; Azhar, E.I. In silico Prediction and Designing of Potential siRNAs to be Used as Antivirals Against SARS-CoV-2. Curr. Pharm. Des. 2021, 27, 1–12. [Google Scholar] [CrossRef]
- McKimm-Breschkin, J.L.; Jiang, S.; Hui, D.S.; Beigel, J.H.; Govorkova, E.A.; Lee, N. Prevention and treatment of respiratory viral infections: Presentations on antivirals, traditional therapies and host-directed interventions at the 5th ISIRV Antiviral Group conference. Antivir. Res. 2018, 149, 118–142. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Fakhr, E.; Zare, F.; Teimoori-Toolabi, L. Precise and efficient siRNA design: A key point in competent gene silencing. Cancer Gene Ther. 2016, 23, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lu, J. In Silico Design of siRNAs Targeting Existing and Future Respiratory Viruses with VirusSi. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sohrab, S.S.; Aly El-Kafrawy, S.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. In silico prediction and experimental vali-dation of siRNAs targeting ORF1ab of MERS-CoV in Vero cell line. Saudi J. Biol. Sci. 2020, 28, 1348–1355. [Google Scholar] [CrossRef]
- Eckerle, I.; Corman, V.M.; Müller, M.A.; Lenk, M.; Ulrich, R.G.; Drosten, C. Replicative Capacity of MERS Coronavirus in Livestock Cell Lines. Emerg. Infect. Dis. 2014, 20, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, M.M.; Akter, R.; Ullah, M.; Abedin, M.; Ullah, G.M.; Hossain, M. A computational approach for predicting role of human microRNAs in MERS-CoV genome. Adv. Bioinform. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Nur, S.M.; Hasan, M.A.; Amin, M.A.; Hossain, M.; Sharmin, T. Design of Potential RNAi (miRNA and siRNA) Molecules for Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Gene Silencing by Computational Method. Interdiscip. Sci. Comput. Life Sci. 2015, 7, 257–265. [Google Scholar] [CrossRef]
- Hashem, A.M.; Algaissi, A.; Agrawal, A.S.; Al-Amri, S.S.; Alhabbab, R.Y.; Sohrab, S.S.; Almasoud, A.S.; Alharbi, N.K.; Peng, B.-H.; Russell, M.; et al. A Highly Immunogenic, Protective, and Safe Adenovirus-Based Vaccine Expressing Middle East Respiratory Syndrome Coronavirus S1-CD40L Fusion Protein in a Transgenic Human Dipeptidyl Peptidase 4 Mouse Model. J. Infect. Dis. 2019, 220, 1558–1567. [Google Scholar] [CrossRef]
- Jeang, K.-T. RNAi in the regulation of mammalian viral infections. BMC Biol. 2012, 10, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Yang, Y.; Huang, J.; Jiang, S.; Du, L. Advances in MERS-CoV vaccines and therapeutics based on the recep-tor-binding domain. Viruses 2019, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Jia, W.; Wang, P.; Zhang, S.; Shi, X.; Wang, X.; Zhang, L. Antibodies and vaccines against Middle East respiratory syndrome coronavirus. Emerg. Microbes Infect. 2019, 8, 841–856. [Google Scholar] [CrossRef] [Green Version]
- Folegatti, P.M.; Bittaye, M.; Flaxman, A.; Lopez, F.R.; Bellamy, D.; Kupke, A.; Mair, C.; Makinson, R.; Sheridan, J.; Rohde, C.; et al. Safety and immunogenicity of a candidate Middle East respiratory syndrome coronavirus viral-vectored vaccine: A dose-escalation, open-label, non-randomised, uncontrolled, phase 1 trial. Lancet Infect. Dis. 2020, 20, 816–826. [Google Scholar] [CrossRef]
- Stein, D.A.; Perry, S.T.; Buck, M.D.; Oehmen, C.S.; Fischer, M.A.; Poore, E.; Smith, J.L.; Lancaster, A.M.; Hirsch, A.J.; Slifka, M.K.; et al. Inhibition of Dengue Virus Infections in Cell Cultures and in AG129 Mice by a Small Interfering RNA Targeting a Highly Conserved Sequence. J. Virol. 2011, 85, 10154–10166. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, B.; Braga, A.C.S.; Batista, M.N.; Harris, M.; Rahal, P. Evaluation of Canonical siRNA and Dicer Substrate RNA for Inhibition of Hepatitis C Virus Genome Replication—A Comparative Study. PLoS ONE 2015, 10, e0117742. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.-S.; Lee, S.-H.; Kim, E.-J.; Cho, H.; Lee, W.; Kim, G.-W.; Park, H.-J.; Cho, S.-W.; Lee, C.; Oh, J.-W. Inhibition of Hepatitis C Virus in Mice by a Small Interfering RNA Targeting a Highly Conserved Sequence in Viral IRES Pseudoknot. PLoS ONE 2016, 11, e0146710. [Google Scholar] [CrossRef] [Green Version]
- Elhefnawi, M.; Kim, T.; Kamar, M.A.; Min, S.; Hassan, N.M.; El-Ahwany, E.; Kim, H.; Zada, S.; Amer, M.; Windisch, M.P. In Silico Design and Experimental Validation of siRNAs Targeting Conserved Regions of Multiple Hepatitis C Virus Genotypes. PLoS ONE 2016, 11, e0159211. [Google Scholar] [CrossRef]
- Ahmed, F.; Sharma, M.; Al-Ghamdi, A.A.; Al-Yami, S.M.; Al-Salami, A.M.; Refai, M.Y.; Warsi, M.K.; Howladar, S.M.; Baeshen, M.N. A Comprehensive Analysis of cis-Acting RNA Elements in the SARS-CoV-2 Genome by a Bioinformatics Approach. Front. Genet. 2020, 11, 572702. [Google Scholar] [CrossRef]
- Doench, J.G.; Petersen, C.P.; Sharp, P.A. siRNAs can function as miRNAs. Genes Dev. 2003, 17, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Shi, B.; Ding, X.; Liu, T.; Hu, X.; Yip, K.Y.; Yang, Z.R.; Mathews, D.H.; Lu, Z.J. Improved prediction of RNA secondary structure by integrating the free energy model with restraints derived from experimental probing data. Nucleic Acids Res. 2015, 43, 7247–7259. [Google Scholar] [CrossRef] [Green Version]
- Kiryu, H.; Terai, G.; Imamura, O.; Yoneyama, H.; Suzuki, K.; Asai, K. A detailed investigation of accessibilities around target sites of siRNAs and miRNAs. Bioinformatics 2011, 27, 1788–1797. [Google Scholar] [CrossRef]
- Ding, Y.; Chan, C.Y.; Lawrence, C.E. RNA secondary structure prediction by centroids in a Boltzmann weighted ensemble. RNA 2005, 11, 1157–1166. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.J.; Mathews, D.H. Efficient siRNA selection using hybridization thermodynamics. Nucleic Acids Res. 2007, 36, 640–647. [Google Scholar] [CrossRef] [Green Version]
- Petrova, N.S.; Zenkova, M.A.; Chernolovskaya, E.L. Structure—Functions Relations in Small Interfering RNAs. Pract. Appl. Biomed. Eng. 2013. [Google Scholar] [CrossRef] [Green Version]
- Walton, S.P.; Wu, M.; Gredell, J.A.; Chan, C. Designing highly active siRNAs for therapeutic applications. FEBS J. 2010, 277, 4806–4813. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Chan, C.Y.; Maliyekkel, A.; Lawrence, C.E.; Roninson, I.B.; Ding, Y. Effect of target secondary structure on RNAi efficiency. RNA 2007, 13, 1631–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajan, S.; Hutvagner, G. RNA-Based Therapeutics: From Antisense Oligonucleotides to miRNAs. Cells 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, Y.; Ui-Tei, K. siRNA Design Software for a Target Gene-Specific RNA Interference. Front. Genet. 2012, 3, 102. [Google Scholar] [CrossRef] [Green Version]
- Naito, Y.; Ui-Tei, K.; Nishikawa, T.; Takebe, Y.; Saigo, K. siVirus: Web-based antiviral siRNA design software for highly divergent viral sequences. Nucleic Acids Res. 2006, 34, W448–W450. [Google Scholar] [CrossRef] [Green Version]
- Naito, Y.; Yoshimura, J.; Morishita, S.; Ui-Tei, K. siDirect 2.0: Updated software for designing functional siRNA with reduced seed-dependent off-target effect. BMC Bioinform. 2009, 10, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, A.; Leake, D.; Boese, Q.; Scaringe, S.; Marshall, W.S.; Khvorova, A. Rational siRNA design for RNA interference. Nat. Biotechnol. 2004, 22, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Amarzguioui, M.; Prydz, H. An algorithm for selection of functional siRNA sequences. Biochem. Biophys. Res. Commun. 2004, 316, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, M.; Murakumo, Y.; Masuda, A.; Matsuura, T.; Asai, N.; Jijiwa, M.; Ishida, M.; Shinmi, J.; Yatsuya, H.; Qiao, S.; et al. Thermodynamic instability of siRNA duplex is a prerequisite for dependable prediction of siRNA activities. Nucleic Acids Res. 2007, 35, e123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.S.; Ahn, J.; Jun, E.J.; Yang, S.; Joo, C.H.; Kim, Y.K.; Lee, H. A novel program to design siRNAs simultaneously effective to highly variable virus genomes. Biochem. Biophys. Res. Commun. 2009, 384, 431–435. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.N. | Position of siRNA in the Genome (Start–End) | Target Sequence | Predicted RNA Oligo Sequences (5′→3′) | Minimum Free Energy (MFE- kcal/mol) and Frequency of Thermodynamic Ensemble (%) |
|---|---|---|---|---|
| 1 | 791–813 | agcaatctatttttactattaat | UAAUAGUAAAAAUAGAUUGCU CAAUCUAUUUUUACUAUUAAU | −17.96, 65.21 |
| 2 | 1615–1637 | atggataatgctattaatgttgg | AACAUUAAUAGCAUUAUCCAU GGAUAAUGCUAUUAAUGUUGG | −21.80, 77.37 |
| 3 | 1910–1932 | gcgactttatgtctacaattatt | UAAUUGUAGACAUAAAGUCGC GACUUUAUGUCUACAAUUAUU | −22.02,69.74 |
| 4 | 4018–4040 | gacactttagatgatatcttaca | UAAGAUAUCAUCUAAAGUGUC CACUUUAGAUGAUAUCUUACA | −22.62,69.54 |
| 5 | 5597–5619 | atgctattagtttgagttttaat | UAAAACUCAAACUAAUAGCAU GCUAUUAGUUUGAGUUUUAAU | −21.51, 83.64 |
| 6 | 5598–5620 | tgctattagtttgagttttaata | UUAAAACUCAAACUAAUAGCA CUAUUAGUUUGAGUUUUAAUA | −19.74, 57.91 |
| 7 | 5819–5841 | gagctagtttgcgtcaaattttt | AAAUUUGACGCAAACUAGCUC GCUAGUUUGCGUCAAAUUUUU | −24.28, 53.63 |
| 8 | 9495–9517 | ctctaatatctttgttattaaca | UUAAUAACAAAGAUAUUAGAG CUAAUAUCUUUGUUAUUAACA | −17.97, 54.45 |
| 9 | 9533–9555 | ctcttagaaactctttaactaat | UAGUUAAAGAGUUUCUAAGAG CUUAGAAACUCUUUAACUAAU | −22.37, 64.54 |
| 10 | 13,605–13,627 | tggtttgattttgttgaaaatcc | AUUUUCAACAAAAUCAAACCA GUUUGAUUUUGUUGAAAAUCC | −18.34, 35.22 |
| 11 | 14,005–14,027 | acgcaaattgcgttaattgtact | UACAAUUAACGCAAUUUGCGU GCAAAUUGCGUUAAUUGUACU | −22.34, 79.46 |
| 12 | 14,389–14,411 | tggtatctaaaggtttctttaag | UAAAGAAACCUUUAGAUACCA GUAUCUAAAGGUUUCUUUAAG | −22.04, 67.95 |
| 13 | 16,177–16,199 | gtcttgtattcggcttatacaag | UGUAUAAGCCGAAUACAAGAC CUUGUAUUCGGCUUAUACAAG | −26.53, 58.68 |
| 14 | 16,217–16,239 | tccttctatagttgaatttaata | UUAAAUUCAACUAUAGAAGGA CUUCUAUAGUUGAAUUUAAUA | −20.24, 48.81 |
| 15 | 17,283–17,305 | gtctacaataataaattgttagc | UAACAAUUUAUUAUUGUAGAC CUACAAUAAUAAAUUGUUAGC | −17.87, 75.42 |
| 16 | 17,583–17,605 | aacaacattaacagatttaatgt | AUUAAAUCUGUUAAUGUUGUU CAACAUUAACAGAUUUAAUGU | −19.59, 62.23 |
| 17 | 18,028–18,050 | ctctacaattaggattttcaact | UUGAAAAUCCUAAUUGUAGAG CUACAAUUAGGAUUUUCAACU | −22.08, 53.94 |
| 18 | 19,806–19,828 | ttgtataagaaagtcaataatga | AUUAUUGACUUUCUUAUACAA GUAUAAGAAAGUCAAUAAUGA | −19.97, 64.53 |
| 19 | 20,090–20,112 | ctcaactattcataactatttta | AAAUAGUUAUGAAUAGUUGAG CAACUAUUCAUAACUAUUUUA | −19.63, 42.01 |
| 20 | 20,498–20,520 | tgccaatatgcgtgttatacatt | UGUAUAACACGCAUAUUGGCA CCAAUAUGCGUGUUAUACAUU | −25.98, 74.21 |
| 21 | 20,948–20,970 | gggtactattaaagaaaatatag | AUAUUUUCUUUAAUAGUACCC GUACUAUUAAAGAAAAUAUAG | −17.65, 66.76 |
| siRNAs | CC50 (nM) | IC50 (nM) | |
|---|---|---|---|
| Supernatant | Lysate | ||
| siRNA-1 | >100 | ND | ND |
| siRNA-2 | >100 | ND | ND |
| siRNA-3 | >100 | ND | ND |
| siRNA-4 | >100 | ND | ND |
| siRNA-5 | >100 | ND | ND |
| siRNA-6 | >100 | 26.22 | 17.56 |
| siRNA-7 | >100 | ND | ND |
| siRNA-8 | >100 | ND | ND |
| siRNA-9 | >100 | ND | ND |
| siRNA-10 | >100 | ND | ND |
| siRNA-11 | >100 | ND | ND |
| siRNA-12 | >100 | ND | ND |
| siRNA-13 | >100 | ND | ND |
| siRNA-14 | >100 | ND | ND |
| siRNA-15 | >100 | ND | ND |
| siRNA-16 | >100 | 13.87 | 16.25 |
| siRNA-17 | >100 | ND | ND |
| siRNA-18 | >100 | ND | ND |
| siRNA-19 | >100 | 11.12 | 5.17 |
| siRNA-20 | >100 | ND | ND |
| siRNA-21 | >100 | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Kafrawy, S.A.; Sohrab, S.S.; Mirza, Z.; Hassan, A.M.; Alsaqaf, F.; Azhar, E.I. In Vitro Inhibitory Analysis of Rationally Designed siRNAs against MERS-CoV Replication in Huh7 Cells. Molecules 2021, 26, 2610. https://doi.org/10.3390/molecules26092610
El-Kafrawy SA, Sohrab SS, Mirza Z, Hassan AM, Alsaqaf F, Azhar EI. In Vitro Inhibitory Analysis of Rationally Designed siRNAs against MERS-CoV Replication in Huh7 Cells. Molecules. 2021; 26(9):2610. https://doi.org/10.3390/molecules26092610
Chicago/Turabian StyleEl-Kafrawy, Sherif Aly, Sayed Sartaj Sohrab, Zeenat Mirza, Ahmed M. Hassan, Fatima Alsaqaf, and Esam Ibraheem Azhar. 2021. "In Vitro Inhibitory Analysis of Rationally Designed siRNAs against MERS-CoV Replication in Huh7 Cells" Molecules 26, no. 9: 2610. https://doi.org/10.3390/molecules26092610
APA StyleEl-Kafrawy, S. A., Sohrab, S. S., Mirza, Z., Hassan, A. M., Alsaqaf, F., & Azhar, E. I. (2021). In Vitro Inhibitory Analysis of Rationally Designed siRNAs against MERS-CoV Replication in Huh7 Cells. Molecules, 26(9), 2610. https://doi.org/10.3390/molecules26092610