
Mining the Royal Jelly Proteins: Combinatorial Hexapeptide Ligand Library Significantly Improves the MS-Based Proteomic Identification in Complex Biological Samples

 , , ,
, , ,
Abstract
:1. Introduction
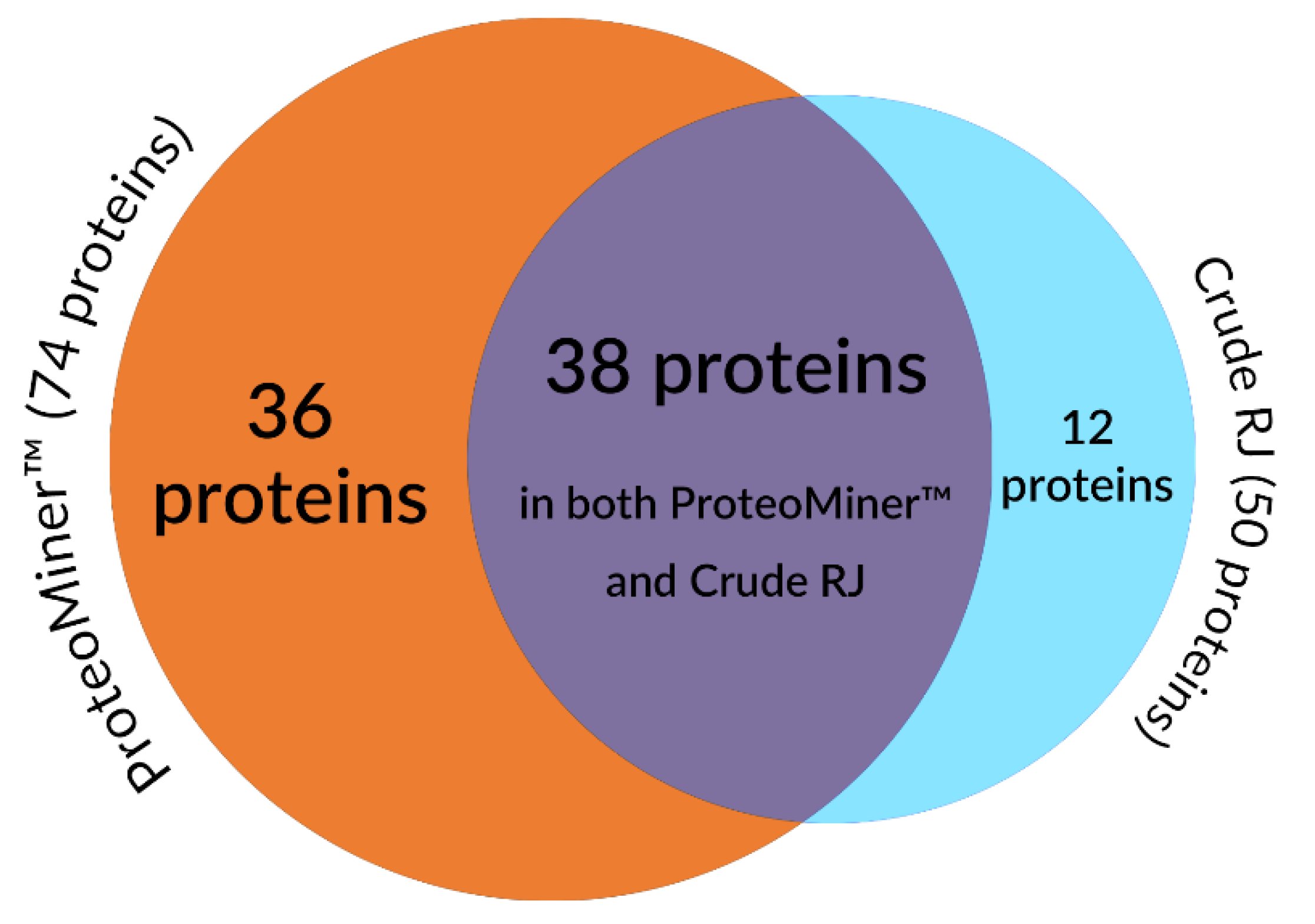
2. Results
3. Discussion
4. Materials and Methods
4.1. Royal Jelly
4.2. Pre-Treatment of the Royal Jelly Samples
4.3. Relative Protein Enrichment—ProteoMinerTM Hexapeptide Ligand Library
4.4. Trypsin Proteolytic Digestion and ZipTip Concentration and Purification
4.5. NanoLC-MALDI-TOF/TOF MS/MS Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Denisow, B.; Denisow-Pietrzyk, M. Biological and therapeutic properties of bee pollen: A review. J. Sci. Food Agric. 2016, 96, 4303–4309. [Google Scholar] [CrossRef]
- Fratini, F.; Cilia, G.; Mancini, S.; Felicioli, A. Royal Jelly: An ancient remedy with remarkable antibacterial properties. Microbiol. Res. 2016, 192, 130–141. [Google Scholar] [CrossRef]
- Taulavuori, K.; Julkunen-Tiitto, R.; Hyöky, V.; Taulavuori, E. Blue Mood for Superfood. Nat. Prod. Commun. 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Ramadan, M.F.; Al-Ghamdi, A. Bioactive compounds and health-promoting properties of royal jelly: A review. J. Funct. Foods 2012, 4, 39–52. [Google Scholar] [CrossRef]
- Wytrychowski, M.; Chenavas, S.; Daniele, G.; Casabianca, H.; Batteau, M.; Guibert, S.; Brion, B. Physicochemical characterisation of French royal jelly: Comparison with commercial royal jellies and royal jellies produced through artificial bee-feeding. J. Food Compos. Anal. 2013, 29, 126–133. [Google Scholar] [CrossRef]
- Sabatini, A.G. Quality and standardisation of Royal Jelly. J. ApiProd. ApiMed. Sci. 2009, 1, 16–21. [Google Scholar] [CrossRef]
- Garcia-Amoedo, L.H.; De Almeida-Muradian, L.B. Physicochemical composition of pure and adulterated royal jelly. Quim. Nova 2007, 30, 257–259. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M. Bee Products-Chemical and Biological Properties; Springer International Publishing: New York, NY, USA, 2017; pp. 1–306. [Google Scholar]
- Priomorac, L.; Bilić Rajs, B.; Puškadija, Z.; Kovačić, M.; Vukadin, I.; Flanjak, I. Physicochemical characteristics of Croatian royal jelly. Croat. J. Food Sci. Technol. 2019, 11, 266–271. [Google Scholar]
- Balkanska, R.; Crenguta, P. Comparison of physicochemical parameters in royal jelly from Romania and Bulgaria. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Anim. Sci. Biotechnol. 2013, 70, 117–121. [Google Scholar]
- Scarselli, R.; Donadio, E.; Giuffrida, M.G.; Fortunato, D.; Conti, A.; Balestreri, E.; Felicioli, R.; Pinzauti, M.; Sabatini, A.G.; Felicioli, A. Towards royal jelly proteome. Proteomics 2005, 5, 769–776. [Google Scholar] [CrossRef]
- Lercker, G.; Caboni, M.F.; Vecchi, M.A.; Sabatini, A.G.; Nanetti, A. Caratterizzazione dei principali costituenti della gelatina reale. Apicoltura 1992, 8, 27–37. [Google Scholar]
- Sano, O.; Kunikata, T.; Kohno, K.; Iwaki, K.; Ikeda, M.; Kurimoto, M. Characterization of Royal Jelly Proteins in both Africanized and European Honeybees (Apis mellifera) by Two-Dimensional Gel Electrophoresis. J. Agric. Food Chem. 2004, 52, 15–20. [Google Scholar] [CrossRef]
- Buttstedt, A.; Moritz, R.F.A.; Erler, S. Origin and function of the major royal jelly proteins of the honeybee (Apis mellifera) as members of the yellow gene family. Biol. Rev. 2014, 89, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Kamakura, M. Royalactin induces queen differentiation in honeybees. Nature 2011, 473, 478–483. [Google Scholar] [CrossRef]
- Buttstedt, A.; Ihling, C.H.; Pietzsch, M.; Moritz, R.F.A. Royalactin is not a royal making of a queen. Nature 2016, 537, E10–E12. [Google Scholar] [CrossRef]
- Fujita, T.; Kozuka-Hata, H.; Ao-Kondo, H.; Kunieda, T.; Oyama, M.; Kubo, T. Proteomic Analysis of the Royal Jelly and Characterization of the Functions of its Derivation Glands in the Honeybee. J. Proteome Res. 2013, 12, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Schönleben, S.; Sickmann, A.; Mueller, M.J.; Reinders, J. Proteome analysis of Apis mellifera royal jelly. Anal. Bioanal. Chem. 2007, 389, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Li, C.; Zhang, L.; Fang, Y.; Feng, M.; Li, J. Novel royal jelly proteins identified by gel-based and gel-free proteomics. J. Agric. Food Chem. 2011, 59, 10346–10355. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, T.; Rakwal, R.; Nam, H.W.; Shibato, J.; Agrawal, G.K.; Kim, Y.S.; Ogawa, Y.; Yoshida, Y.; Kouzuma, Y.; Masuo, Y.; et al. Comprehensive royal jelly (RJ) proteomics using one- and two-dimensional proteomics platforms reveals novel RJ proteins and potential phospho/glycoproteins. J. Proteome Res. 2008, 7, 3194–3229. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, T.; Zhang, Z.; Pan, Y. Proteomic analysis of royal jelly from three strains of western honeybees (Apis mellifera). J. Agric. Food Chem. 2007, 55, 8411–8422. [Google Scholar] [CrossRef]
- Hu, F.L.; Bíliková, K.; Casabianca, H.; Daniele, G.; Salmen Espindola, F.; Feng, M.; Guan, C.; Han, B.; Krištof Kraková, T.; Li, J.K.; et al. Standard methods for Apis mellifera royal jelly research. J. Apic. Res. 2017, 58, 1–68. [Google Scholar] [CrossRef] [Green Version]
- Collazo, N.; Carpena, M.; Nuñez-Estevez, B.; Otero, P.; Simal-Gandara, J.; Prieto, M.A. Health Promoting Properties of Bee Royal Jelly: Food of the Queens. Nutrients 2021, 13, 543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Han, B.; Li, R.; Lu, X.; Nie, A.; Guo, L.; Fang, Y.; Feng, M.; Li, J. Comprehensive identification of novel proteins and N-glycosylation sites in royal jelly. BMC Genom. 2014, 15, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Fang, Y.; Li, R.; Feng, M.; Han, B.; Zhou, T.; Li, J. Towards posttranslational modification proteome of royal jelly. J. Proteom. 2012, 75, 5327–5341. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Fang, Y.; Feng, M.; Lu, X.; Huo, X.; Meng, L.; Wu, B.; Li, J. In-depth phosphoproteomic analysis of royal jelly derived from western and eastern honeybee species. J. Proteome Res. 2014, 13, 5928–5943. [Google Scholar] [CrossRef] [PubMed]
- Greco, V.; Piras, C.; Pieroni, L.; Ronci, M.; Putignani, L.; Roncada, P.; Urbani, A. Applications of MALDI-TOF mass spectrometry in clinical proteomics. Expert Rev. Proteom. 2018, 15, 683–696. [Google Scholar] [CrossRef]
- Hajduk, J.; Matysiak, J.; Kokot, Z.J. Challenges in biomarker discovery with MALDI-TOF MS. Clin. Chim. Acta 2016, 458, 84–98. [Google Scholar] [CrossRef]
- Li, S.; He, Y.; Lin, Z.; Xu, S.; Zhou, R.; Liang, F.; Wang, J.; Yang, H.; Liu, S.; Ren, Y. Digging More Missing Proteins Using an Enrichment Approach with ProteoMiner. J. Proteome Res. 2017, 16, 4330–4339. [Google Scholar] [CrossRef]
- Righetti, P.G.; Boschetti, E.; Lomas, L.; Citterio, A. Protein EqualizerTM Technology: The quest for a “democratic proteome”. Proteomics 2006, 6, 3980–3992. [Google Scholar] [CrossRef]
- Boschetti, E.; Righetti, P.G. The ProteoMinerin the proteomic arena: A non-depleting tool for discovering low-abundance species. J. Proteom. 2008, 71, 255–264. [Google Scholar] [CrossRef]
- Drapeau, M.D.; Albert, S.; Kucharski, R.; Prusko, C.; Maleszka, R. Evolution of the Yellow/Major Royal Jelly Protein family and the emergence of social behavior in honey bees. Genome. Res. 2006, 16, 1385–1394. [Google Scholar] [CrossRef] [Green Version]
- Santos, K.S.; Delazari dos Santos, L.; Mendes, M.; Monson de Souza, B.; Malaspina, O.; Palma, M.S. Profiling the proteome complement of the secretion from hypopharyngeal gland of Africanized nurse-honeybees (Apis mellifera L.). Insect Biochem. Mol. Biol. 2005, 35, 85–91. [Google Scholar] [CrossRef]
- Schmitzová, J.; Klaudiny, J.; Albert, Š.; Schröder, W.; Schreckengost, W.; Hanes, J.; Júdová, J.; Šimúth, J. A family of major royal jelly proteins of the honeybee Apis mellifera L. Cell. Mol. Life Sci. 1998, 54, 1020–1030. [Google Scholar] [CrossRef]
- Hanes, J.; Šimuth, J. Identification and partial characterisation of the major royal jelly protein of the honey bee (Apis mellifera L.). J. Apic. Res. 1992, 31, 22–26. [Google Scholar] [CrossRef]
- Ramanathan, A.N.K.G.; Nair, A.J.; Sugunan, V.S. A review on Royal Jelly proteins and peptides. J. Funct. Foods 2018, 44, 255–264. [Google Scholar] [CrossRef]
- Buttstedt, A.; Mureşan, C.I.; Lilie, H.; Hause, G.; Ihling, C.H.; Schulze, S.H.; Pietzsch, M.; Moritz, R.F.A. How Honeybees Defy Gravity with Royal Jelly to Raise Queens. Curr. Biol. 2018, 28, 1095–1100.e3. [Google Scholar] [CrossRef] [Green Version]
- Pirk, C.W.W. Honeybee Evolution: Royal Jelly Proteins Help Queen Larvae to Stay on Top. Curr. Biol. 2018, 28, R350–R351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mureşan, C.I.; Buttstedt, A. pH-dependent stability of honey bee (Apis mellifera) major royal jelly proteins. Sci. Rep. 2019, 9, 3–8. [Google Scholar] [CrossRef]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, e1006512. [Google Scholar] [CrossRef]
- Oliver, W.T.; Wells, J.E. Lysozyme as an alternative to growth promoting antibiotics in swine production. J. Anim. Sci. Biotechnol. 2015, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- McCleskey, C.S.; Melampy, R.M. Bactericidal Properties of Royal Jelly of the Honeybee. J. Econ. Entomol. 1939, 32, 581–587. [Google Scholar] [CrossRef]
- Bílikova, K.; Huang, S.-C.; Lin, I.-P.; Šimuth, J.; Peng, C.-C. Structure and antimicrobial activity relationship of royalisin, an antimicrobial peptide from royal jelly of Apis mellifera. Peptides 2015, 68, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, D.; Li, M.; Jin, F.; Din, M.; Parnell, L.D.; Lai, C.-Q. Mechanism of Action of Recombinant Acc-Royalisin from Royal Jelly of Asian Honeybee against Gram-Positive Bacteria. PLoS ONE 2012, 7, e47194. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Chou, W.M.; Widowati, D.A.; Lin, I.P.; Peng, C.C. 10-Hydroxy-2-Decenoic Acid of Royal Jelly Exhibits Bactericide and Anti-Inflammatory Activity in Human Colon Cancer Cells. BMC Complement. Altern. Med. 2018, 18, 202. [Google Scholar] [CrossRef] [PubMed]
- Kurek-Górecka, A.; Górecki, M.; Rzepecka-Stojko, A.; Balwierz, R.; Stojko, J. Bee Products in Dermatology and Skin Care. Molecules 2020, 25, 556. [Google Scholar] [CrossRef] [Green Version]
- Pavel, C.I.; Mărghitaş, L.A.; Bobiş, O.; Dezmirean, D.S.; Şapcaliu, A.; Radoi, I.; Mădaş, M.N. Biological Activities of Royal Jelly—Review. Sci. Pap. Anim. Sci. Biotechnol. 2011, 44, 108–118. [Google Scholar]
- Spillner, E.; Blank, S.; Jakob, T. Hymenoptera Allergens: From Venom to “Venome”. Front. Immunol. 2014, 5, 77. [Google Scholar] [CrossRef] [Green Version]
- Tomsitz, D.; Brockow, K. Component Resolved Diagnosis in Hymenoptera Anaphylaxis. Curr. Allergy Asthma Rep. 2017, 17, 38. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.R. Hymenoptera venom allergens. Clin. Rev. Allergy Immunol. 2006, 30, 109–128. [Google Scholar] [CrossRef]
- Takahama, H.; Shimazu, T. Food-induced anaphylaxis caused by ingestion of royal jelly. J. Dermatol. 2006, 33, 424–426. [Google Scholar] [CrossRef]
- Katayama, M.; Aoki, M.; Kawana, S. Case of anaphylaxis caused by ingestion of royal jelly. J. Dermatol. 2008, 35, 222–224. [Google Scholar] [CrossRef]
- Mizutani, Y.; Shibuya, Y.; Takahashi, T.; Tsunoda, T.; Moriyama, T.; Seishima, M. Major royal jelly protein 3 as a possible allergen in royal jelly-induced anaphylaxis. J. Dermatol. 2011, 38, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Leung, R.; Lam, C.W.K.; Ho, A.; Chan, J.K.W.; Choy, D.; Lai, C.K.W. Allergic sensitisation to common environmental allergens in adult asthmatics in Hong Kong. Hong Kong Med. J. 1997, 3, 211–217. [Google Scholar]
- Hata, T.; Furusawa-Horie, T.; Arai, Y.; Takahashi, T.; Seishima, M.; Ichihara, K. Studies of royal jelly and associated cross-reactive allergens in atopic dermatitis patients. PLoS ONE 2020, 15, e0233707. [Google Scholar] [CrossRef] [PubMed]
- Matysiak, J.; Hajduk, J.; Światły, A.; Naskret, N.; Kokot, Z.J. Proteomic analysis of Apis mellifera venom determined by liquid chromatography (LC) coupled with nano-LC-MALDI-TOF/TOF MS. Acta. Pol. Pharm. Drug Res. 2017, 74, 53–65. [Google Scholar]
- Matysiak, J.; Hajduk, J.; Mayer, F.; Hebeler, R.; Kokot, Z.J. Hyphenated LC–MALDI–ToF/ToF and LC–ESI–QToF approach in proteomic characterisation of honeybee venom. J. Pharm. Biomed. Anal. 2016, 121, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Norton, R.S. Conotoxin gene superfamilies. Mar. Drugs 2014, 12, 6058–6101. [Google Scholar] [CrossRef] [Green Version]
- Daly, N.; Craik, D. Structural Studies of Conotoxins. IUBMB Life 2009, 61, 144–150. [Google Scholar] [CrossRef]
- Dao, F.-Y.; Yang, H.; Su, Z.-D.; Yang, W.; Wu, Y.; Hui, D.; Chen, W.; Tang, H.; Lin, H. Recent Advances in Conotoxin Classification by Using Machine Learning Methods. Molecules 2017, 22, 1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Campos, M.G.; Fratini, F.; Altaye, S.Z.; Li, J. New insights into the biological and pharmaceutical properties of royal jelly. Int. J. Mol. Sci. 2020, 21, 382. [Google Scholar] [CrossRef] [Green Version]
- Minami, A.; Matsushita, H.; Ieno, D.; Matsuda, Y.; Horii, Y.; Ishii, A.; Takahashi, T.; Kanazawa, H.; Wakatsuki, A.; Suzuki, T. Improvement of neurological disorders in postmenopausal model rats by administration of royal jelly. Climacteric 2016, 19, 568–573. [Google Scholar] [CrossRef]
- Mohamed, A.A.-R.; Galal, A.A.A.; Elewa, Y.H.A. Comparative protective effects of royal jelly and cod liver oil against neurotoxic impact of tartrazine on male rat pups brain. Acta Histochem. 2015, 117, 649–658. [Google Scholar] [CrossRef]
- Pan, Y.; Xu, J.; Jin, P.; Yang, Q.; Zhu, K.; You, M.; Hu, F.; Chen, M. Royal Jelly Ameliorates Behavioral Deficits, Cholinergic System Deficiency, and Autonomic Nervous Dysfunction in Ovariectomized Cholesterol-Fed Rabbits. Molecules 2019, 24, 1149. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, N.; Morpurgo, N.; Linial, M. Novel Families of Toxin-like Peptides in Insects and Mammals: A Computational Approach. J. Mol. Biol. 2007, 369, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Gätschenberger, H.; Gimple, O.; Tautz, J.; Beier, H. Honey bee drones maintain humoral immune competence throughout all life stages in the absence of vitellogenin production. J. Exp. Biol. 2012, 215, 1313–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrowski, L.A.; Hall, A.C.; Mekhail, K. Ataxin-2: From RNA Control to Human Health and Disease. Genes 2017, 8, 157. [Google Scholar] [CrossRef] [PubMed]
- Carmo-Silva, S.; Nobrega, C.; Pereira de Almeida, L.; Cavadas, C. Unraveling the Role of Ataxin-2 in Metabolism. Trends Endocrinol. Metab. 2017, 28, 309–318. [Google Scholar] [CrossRef]
- Zhang, Y.; Ling, J.; Yuan, C.; Dubruille, R.; Emery, P. A role for Drosophila ATX2 in activation of PER translation and circadian behavior. Science 2013, 340, 879–882. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.; Allada, R. ATAXIN-2 Activates PERIOD Translation to Sustain Circadian Rhythms in Drosophila. Science 2013, 340, 875–879. [Google Scholar] [CrossRef] [Green Version]
- Vianna, M.C.B.; Poleto, D.C.; Gomes, P.F.; Valente, V.; Paçó-Larson, M.L. Drosophila ataxin-2 gene encodes two differentially expressed isoforms and its function in larval fat body is crucial for development of peripheral tissues. FEBS OpenBio 2016, 6, 1040–1453. [Google Scholar] [CrossRef]
- Venthur, H.; Zhou, J.-J. Odorant Receptors and Odorant-Binding Proteins as Insect Pest Control Targets: A Comparative Analysis. Front. Physiol. 2018, 9, 1163. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A. Biotechnology I Insect Biotechnologie in Drug Discovery, 1st ed.; Vilcinskas, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; p. 201. [Google Scholar]



{kind=link}
{kind=link}
| Accession | Protein Name | Function | MW [kDa] | pI | CRJ | F1 | F2 | F3 | F4 |
|---|---|---|---|---|---|---|---|---|---|
| gi|110749126 | Predicted: glucose dehydrogenase [acceptor] isoform 3 [Apis mellifera] | Enzyme | 70.1 | 6.7 | x | ||||
| gi|110751029 | Predicted: e3 ubiquitin-protein ligase IAP-3-like [Apis mellifera] | Enzyme | 43.2 | 6.4 | x | ||||
| gi|110756431 | Predicted: hypothetical protein LOC725074 [Apis mellifera] | 8.3 | 10.2 | x | |||||
| gi|110758964 | Predicted: regucalcin-like [Apis mellifera] | Binding | 10.2 | 9.4 | x | x | |||
| gi|110763647 | Predicted: hypothetical protein LOC726323 [Apis mellifera] | 18.5 | 7.9 | x | |||||
| gi|13184963 | defensin [Apis mellifera] | Defense | 6.3 | 6.5 | x | ||||
| gi|166795901 | apolipophorin-III-like protein precursor [Apis mellifera] | Binding | 21.3 | 5.4 | x | x | x | x | x |
| gi|202078658 | defensin [Apis cerana cerana] | Defense | 10.6 | 7.8 | x | ||||
| gi|202078660 | defensin [Apis cerana cerana] | Defense | 10.7 | 7.8 | x | ||||
| gi|254548151 | defensin precursor [Apis cerana] | Defense | 8.7 | 6.4 | x | x | x | x | x |
| gi|254548155 | defensin precursor [Apis mellifera] | Defense | 8.8 | 5.9 | x | x | x | x | x |
| gi|254910938 | defensin-1 preproprotein [Apis mellifera] | Defense | 10.7 | 6.4 | x | ||||
| gi|258678306 | MRJP9 [Apis cerana] | Honeybee nutrition and development | 19.4 | 9.2 | x | ||||
| gi|258678310 | MRJP5 [Apis dorsata] | Honeybee nutrition and development | 21.3 | 4.7 | x | ||||
| gi|258678314 | MRJP6 [Apis florea] | Honeybee nutrition and development | 21.5 | 4.4 | x | x | |||
| gi|258678316 | MRJP9 [Apis florea] | Honeybee nutrition and development | 21.0 | 5.5 | x | x | x | x | x |
| gi|283105164 | alpha-glucosidase III [Apis dorsata] | Enzyme | 65.5 | 5.0 | x | x | |||
| gi|284182838 | major royal jelly protein 4 [Apis mellifera] | Honeybee nutrition and development | 53.0 | 5.9 | x | x | x | x | |
| gi|284812514 | MRJP5 [Apis mellifera] | Honeybee nutrition and development | 70.1 | 6.1 | x | x | x | x | x |
| gi|288872651 | major royal jelly protein [Apis mellifera] | Honeybee nutrition and development | 61.6 | 6.7 | x | x | x | x | x |
| gi|28972896 | major royal jelly protein-like protein [Apis dorsata] | Honeybee nutrition and development | 9.2 | 9.8 | x | x | x | x | x |
| gi|328775853 | Predicted: DNA replication licensing factor MCM4-like [Apis mellifera] | Binding | 80.7 | 6.9 | x | ||||
| gi|328777366 | Predicted: hypothetical protein LOC100577348 [Apis mellifera] | 61.1 | 10.5 | x | |||||
| gi|328779534 | Predicted: hypothetical protein LOC552041 [Apis mellifera] | 79.3 | 4.2 | x | |||||
| gi|328782858 | Predicted: hypothetical protein LOC410515 [Apis mellifera] | 190.2 | 6.4 | x | |||||
| gi|328783362 | Predicted: hypothetical protein LOC725249 [Apis mellifera] | 6.3 | 4.9 | x | |||||
| gi|328783471 | Predicted: hypothetical protein LOC725114 isoform 1 [Apis mellifera] | 10.1 | 5.7 | x | |||||
| gi|328784821 | Predicted: hypothetical protein LOC100577210 [Apis mellifera] | 11.6 | 5.9 | x | x | x | x | ||
| gi|328789019 | Predicted: protein SERAC1-like [Apis mellifera] | Remodeling | 87.5 | 9.2 | x | ||||
| gi|328790726 | Predicted: venom acid phosphatase Acph-1-like [Apis mellifera] | Enzyme | 42.6 | 8.5 | x | ||||
| gi|328792767 | Predicted: hypothetical protein LOC724993 [Apis mellifera] | 43.8 | 5.4 | x | |||||
| gi|328793775 | Predicted: s-adenosylmethionine decarboxylase proenzyme-like, partial [Apis mellifera] | Enzyme | 32.1 | 4.6 | x | ||||
| gi|328794347 | Predicted: major royal jelly protein 3-like, partial [Apis mellifera] | Honeybee nutrition and development | 42.7 | 6.5 | x | x | |||
| gi|33358394 | major royal jelly protein MRJP1 [Apis cerana cerana] | Honeybee nutrition and development | 49.0 | 5.4 | x | x | x | x | x |
| gi|380011960 | Predicted: slit homolog 2 protein-like [Apis florea] | Binding | 155.3 | 6.0 | x | ||||
| gi|380012917 | Predicted: uncharacterized protein LOC100870850 [Apis florea] | 18.9 | 7.2 | x | |||||
| gi|380013532 | Predicted: Low Quality Protein: clathrin heavy chain-like [Apis florea] | Binding | 187.7 | 5.6 | x | ||||
| gi|380016522 | Predicted: probable bifunctional methylenetetrahydrofolate dehydrogenase/cyclohydrolase 2-like [Apis florea] | Enzyme | 39.9 | 9.6 | x | ||||
| gi|380017034 | Predicted: glucosylceramidase-like [Apis florea] | Enzyme | 60.8 | 5.5 | x | x | |||
| gi|380019073 | Predicted: lysozyme 1-like isoform 2 [Apis florea] | Enzyme | 13.8 | 4.6 | x | x | x | ||
| gi|380020436 | Predicted: regucalcin-like [Apis florea] | Binding | 37.7 | 5.3 | x | x | x | x | |
| gi|380022658 | Predicted: major royal jelly protein 3-like [Apis florea] | Honeybee nutrition and development | 62.7 | 7.8 | x | x | |||
| gi|380022660 | Predicted: major royal jelly protein 4-like [Apis florea] | Honeybee nutrition and development | 56.1 | 6.1 | x | ||||
| gi|380022665 | Predicted: major royal jelly protein 1-like [Apis florea] | Honeybee nutrition and development | 43.9 | 5.3 | x | x | x | x | x |
| gi|380022667 | Predicted: major royal jelly protein 2-like [Apis florea] | Honeybee nutrition and development | 49.1 | 5.7 | x | x | |||
| gi|380022669 | Predicted: major royal jelly protein 2-like [Apis florea] | Honeybee nutrition and development | 49.3 | 6.0 | x | x | x | x | x |
| gi|380022673 | Predicted: major royal jelly protein 5-like [Apis florea] | Honeybee nutrition and development | 34.2 | 4.7 | x | ||||
| gi|380022681 | Predicted: major royal jelly protein 5-like isoform 2 [Apis florea] | Honeybee nutrition and development | 47.5 | 9.2 | x | ||||
| gi|380023404 | Predicted: uncharacterized protein LOC100869599 [Apis florea] | 18.7 | 9.5 | x | x | x | |||
| gi|380024584 | Predicted: chymotrypsin inhibitor-like [Apis florea] | Enzyme inhibitor | 5.6 | 10.0 | x | ||||
| gi|380024588 | Predicted: chymotrypsin inhibitor-like [Apis florea] | Enzyme inhibitor | 8.0 | 5.0 | x | ||||
| gi|380025248 | Predicted: alkylated DNA repair protein alkB homolog 8-like [Apis florea] | Enzyme | 68.2 | 9.3 | x | x | |||
| gi|380025500 | Predicted: venom acid phosphatase Acph-1-like [Apis florea] | Enzyme | 39.9 | 4.9 | x | ||||
| gi|380025661 | Predicted: glucose dehydrogenase [acceptor]-like [Apis florea] | Enzyme | 67.9 | 6.4 | x | x | x | x | |
| gi|380026601 | Predicted: uncharacterized protein LOC100863702 [Apis florea] | 9.9 | 5.0 | x | x | ||||
| gi|380027252 | Predicted: uncharacterized protein LOC100864410 [Apis florea] | 33.4 | 9.7 | x | |||||
| gi|380028593 | Predicted: Low Quality Protein: delta-1-pyrroline-5-carboxylate synthase-like [Apis florea] | Enzyme | 84.9 | 9.1 | x | ||||
| gi|40218299 | major royal jelly protein MRJP5 [Apis cerana cerana] | Honeybee nutrition and development | 70.5 | 8.8 | x | ||||
| gi|40218301 | major royal jelly protein MRJP2 [Apis cerana cerana] | Honeybee nutrition and development | 53.0 | 9.1 | x | x | x | ||
| gi|40557703 | major royal jelly protein MRJP1 precursor [Apis cerana] | Honeybee nutrition and development | 48.9 | 5.4 | x | x | x | x | x |
| gi|40557705 | major royal jelly protein MRJP2 precursor [Apis cerana] | Honeybee nutrition and development | 52.5 | 8.9 | x | x | x | x | x |
| gi|42601246 | major royal jelly protein MRJP5 precursor [Apis cerana] | Honeybee nutrition and development | 68.2 | 9.3 | x | x | |||
| gi|46358503 | major royal jelly protein 2 [Apis cerana] | Honeybee nutrition and development | 52.4 | 8.9 | x | x | x | x | x |
| gi|48094573 | Predicted: hypothetical protein LOC408608 [Apis mellifera] | 19.4 | 7.5 | x | x | ||||
| gi|48101366 | Predicted: venom serine protease 34 [Apis mellifera] | Enzyme | 44.6 | 5.9 | x | ||||
| gi|562090 | defensin precursor [Apis mellifera] | Defense | 10.7 | 6.4 | x | x | |||
| gi|56422035 | major royal jelly protein 3 [Apis mellifera carnica] | Honeybee nutrition and development | 65.7 | 7.1 | x | x | x | x | x |
| gi|56422037 | major royal jelly protein 3 [Apis cerana] | Honeybee nutrition and development | 69.2 | 9.3 | x | x | x | ||
| gi|56422041 | major royal jelly protein 3 [Apis florea] | Honeybee nutrition and development | 59.3 | 6.4 | x | ||||
| gi|57546160 | major rojal jelly protein 7 [Apis cerana] | Honeybee nutrition and development | 24.9 | 5.5 | x | x | x | x | x |
| gi|58585090 | glucose oxidase [Apis mellifera] | Enzyme | 67.9 | 6.5 | x | x | |||
| gi|58585098 | major royal jelly protein 1 precursor [Apis mellifera] | Honeybee nutrition and development | 48.9 | 5.0 | x | x | x | x | x |
| gi|58585108 | major royal jelly protein 2 precursor [Apis mellifera] | Honeybee nutrition and development | 51.0 | 7.0 | x | x | x | x | x |
| gi|58585138 | major royal jelly protein 5 precursor [Apis mellifera] | Honeybee nutrition and development | 70.2 | 5.9 | x | ||||
| gi|58585142 | major royal jelly protein 3 precursor [Apis mellifera] | Honeybee nutrition and development | 61.6 | 6.5 | x | x | x | ||
| gi|58585164 | alpha-glucosidase precursor [Apis mellifera] | Enzyme | 65.5 | 4.9 | x | ||||
| gi|58585170 | major royal jelly protein 4 precursor [Apis mellifera] | Honeybee nutrition and development | 52.9 | 5.9 | x | x | x | x | x |
| gi|58585188 | major royal jelly protein 6 precursor [Apis mellifera] | Honeybee nutrition and development | 49.8 | 5.9 | x | x | x | x | x |
| gi|60115688 | icarapin-like precursor [Apis mellifera] | Venom carbohydrate-rich protein | 24.8 | 4.4 | x | x | x | x | |
| gi|62198227 | major royal jelly protein 7 precursor [Apis mellifera] | Honeybee nutrition and development | 50.5 | 4.8 | x | x | x | x | x |
| gi|66504790 | Predicted: integrator complex subunit 7-like isoform 1 [Apis mellifera] | snRNA processing | 105.9 | 9.5 | x | ||||
| gi|66511554 | Predicted: glucosylceramidase-like isoform 1 [Apis mellifera] | Enzyme | 59.3 | 5.2 | x | x | x | x | x |
| gi|66564326 | Predicted: plasma glutamate carboxypeptidase-like isoform 1 [Apis mellifera] | Enzyme | 52.9 | 5.1 | x | ||||
| gi|66565246 | Predicted: lysozyme isoform 1 [Apis mellifera] | Enzyme | 17.1 | 4.6 | x | x | |||
| gi|76496465 | major royal jelly protein 3 [Apis dorsata] | Honeybee nutrition and development | 66.9 | 6.6 | x | x | x | x | x |
| gi|94471624 | icarapin variant 2 precursor [Apis mellifera] | Venom carbohydrate-rich protein | 19.6 | 4.2 | x |
| Accession | Protein Name | Significant Alignment Sequence | Query Coverage | Percent Identity | E-Value |
|---|---|---|---|---|---|
| gi|110756431 | Predicted: hypothetical protein LOC725074 [Apis mellifera] | omega-conotoxin-like protein 1 [Apis mellifera] | 100% | 100.00% | 2.00 × 10−47 |
| gi|110763647 | Predicted: hypothetical protein LOC726323 [Apis mellifera] | uncharacterized protein LOC726323 isoform X1 [Apis mellifera] | 100% | 99.39% | 1.00 × 10−116 |
| gi|328777366 | Predicted: hypothetical protein LOC100577348 [Apis mellifera] | uncharacterized protein LOC100577348 isoform X2 [Apis mellifera] | 98% | 98.37% | 0.0 |
| gi|328779534 | Predicted: hypothetical protein LOC552041 [Apis mellifera] | uncharacterized protein LOC102655185 isoform X1 [Apis mellifera] | 100% | 71.55% | 5.00 × 10−92 |
| gi|328782858 | Predicted: hypothetical protein LOC410515 [Apis mellifera] | uncharacterized protein LOC410515 [Apis mellifera] | 93% | 100.00% | 0.0 |
| chorion peroxidase [Habropoda laboriosa] | 93% | 62.86% | 0.0 | ||
| gi|328783362 | Predicted: hypothetical protein LOC725249 [Apis mellifera] | chymotrypsin inhibitor-like [Apis mellifera] | 84% | 100.00% | 1.00 × 10−25 |
| gi|328783471 | Predicted: hypothetical protein LOC725114 isoform 1 [Apis mellifera] | chymotrypsin inhibitor [Apis mellifera] | 100% | 100.00% | 2.00 × 10−59 |
| gi|328784821 | Predicted: hypothetical protein LOC100577210 [Apis mellifera] | uncharacterized protein LOC413627 [Apis mellifera] | 77% | 100.00% | 6.00 × 10−45 |
| regucalcin [Apis cerana cerana] | 77% | 94.81% | 4.00 × 10−45 | ||
| gi|328792767 | Predicted: hypothetical protein LOC724993 [Apis mellifera] | uncharacterized protein LOC724993 [Apis mellifera] | 83% | 90.37% | 0.00 × 10+00 |
| Predicted: kielin/chordin-like protein [Trachymyrmex zeteki] | 82% | 45.91% | 2.00 × 10−85 | ||
| kielin/chordin-like protein [Nasonia vitripennis] | 81% | 47.06% | 3.00 × 10−89 | ||
| gi|380012917 | Predicted: uncharacterized protein LOC100870850 [Apis florea] | uncharacterized protein LOC408608 [Apis mellifera] | 99% | 73.33% | 2.00 × 10−84 |
| gi|380023404 | Predicted: uncharacterized protein LOC100869599 [Apis florea] | uncharacterized protein LOC102654257 [Apis mellifera] | 100% | 94.71% | 7.00 × 10−116 |
| low-density lipoprotein receptor 1-like isoform X3 [Vespa mandarinia] | 98% | 55.88% | 7.00 × 10−57 | ||
| gi|380026601 | Predicted: uncharacterized protein LOC100863702 [Apis florea] | ataxin-2 [Apis cerana cerana] | 77% | 88.41% | 5.00 × 10−30 |
| gi|380027252 | Predicted: uncharacterized protein LOC100864410 [Apis florea] | odorant receptor Or1 [Apis mellifera] | 98% | 89.68% | 0.0 |
| gi|48094573 | Predicted: hypothetical protein LOC408608 [Apis mellifera] | uncharacterized protein LOC408608 [Apis mellifera] | 100% | 100.00% | 3.00 × 10−124 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matuszewska, E.; Matysiak, J.; Rosiński, G.; Kędzia, E.; Ząbek, W.; Zawadziński, J.; Matysiak, J. Mining the Royal Jelly Proteins: Combinatorial Hexapeptide Ligand Library Significantly Improves the MS-Based Proteomic Identification in Complex Biological Samples. Molecules 2021, 26, 2762. https://doi.org/10.3390/molecules26092762
Matuszewska E, Matysiak J, Rosiński G, Kędzia E, Ząbek W, Zawadziński J, Matysiak J. Mining the Royal Jelly Proteins: Combinatorial Hexapeptide Ligand Library Significantly Improves the MS-Based Proteomic Identification in Complex Biological Samples. Molecules. 2021; 26(9):2762. https://doi.org/10.3390/molecules26092762
Chicago/Turabian StyleMatuszewska, Eliza, Joanna Matysiak, Grzegorz Rosiński, Elżbieta Kędzia, Weronika Ząbek, Jarosław Zawadziński, and Jan Matysiak. 2021. "Mining the Royal Jelly Proteins: Combinatorial Hexapeptide Ligand Library Significantly Improves the MS-Based Proteomic Identification in Complex Biological Samples" Molecules 26, no. 9: 2762. https://doi.org/10.3390/molecules26092762
APA StyleMatuszewska, E., Matysiak, J., Rosiński, G., Kędzia, E., Ząbek, W., Zawadziński, J., & Matysiak, J. (2021). Mining the Royal Jelly Proteins: Combinatorial Hexapeptide Ligand Library Significantly Improves the MS-Based Proteomic Identification in Complex Biological Samples. Molecules, 26(9), 2762. https://doi.org/10.3390/molecules26092762