
Health Benefits of Cereal Grain- and Pulse-Derived Proteins

Abstract
:1. Introduction
2. Protein Content and Amino Acid Composition
Challenges and Future Trends
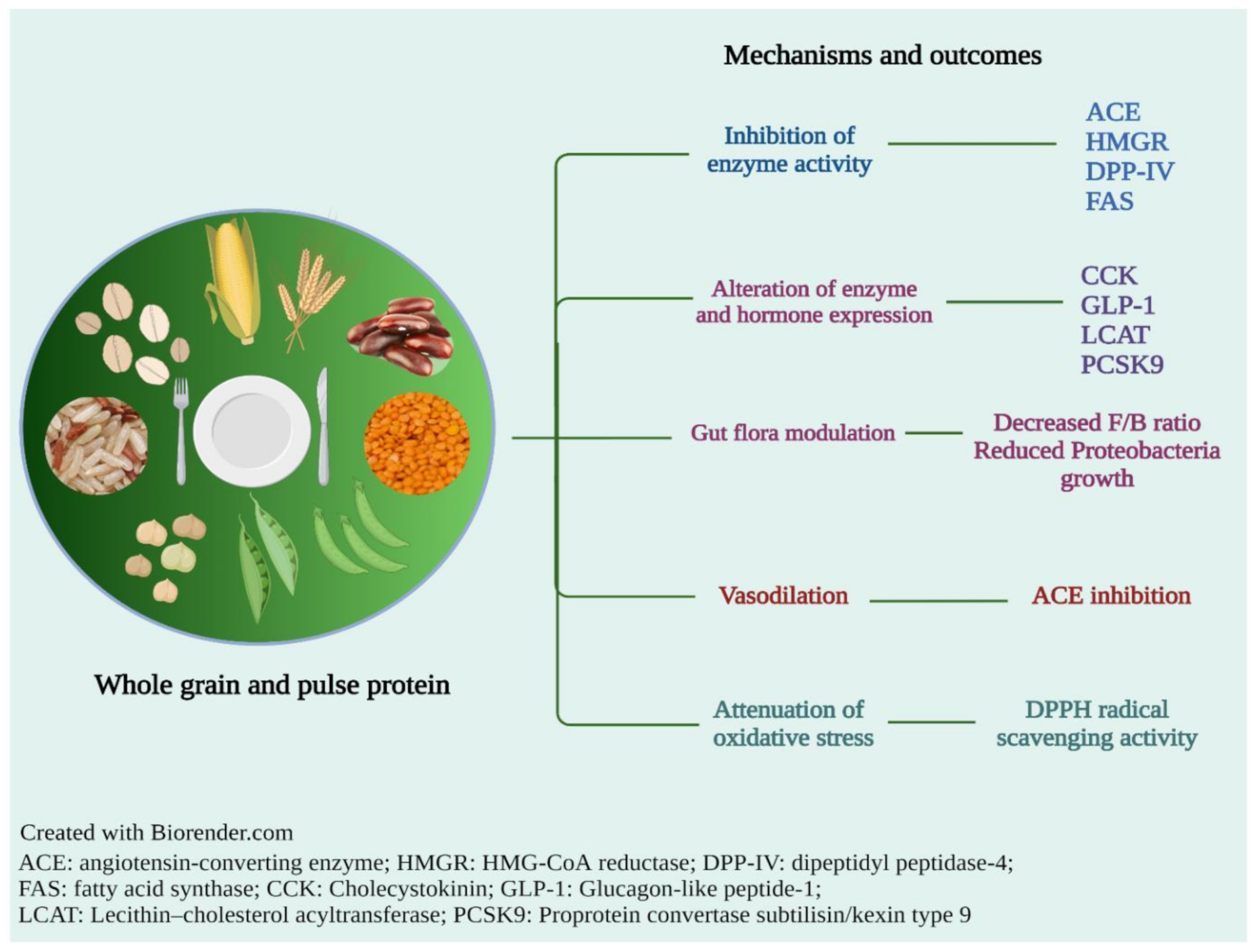
3. Evidence of Health Benefits of Cereal and Pulse Grain Proteins
3.1. Obesity
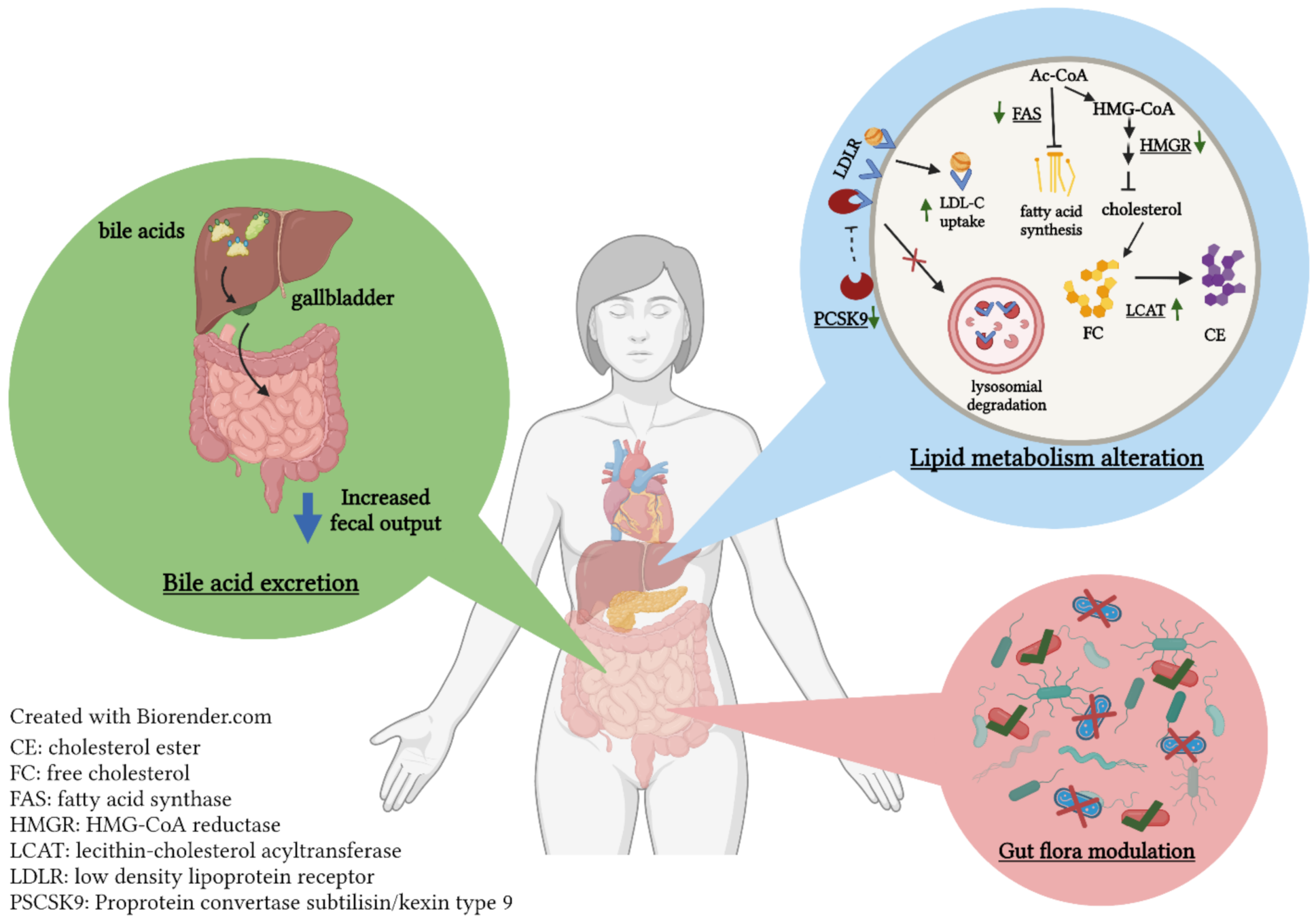
3.2. Hypercholesterolemia
3.3. Diabetes
3.4. Cardiovascular Disease
3.5. Cancer
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACE | Angiotensin I-converting enzyme |
| ANC | Antinutritional compound |
| BAD | Bcl-2-associated death promoter |
| Bcl-2 | B cell lymphoma 2 |
| BIRC7 | Baculoviral IAP repeat-containing protein 7 |
| CCK | Cholecystokinin |
| CDK4 | Cyclin-dependent kinase 4 |
| CE | Cholesterol ester |
| CPH | Chickpea protein hydrolysate |
| CVD | Cardiovascular disease |
| CYP7A1 | Cholesterol 7 alpha-hydroxylase |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| FRB | Fermented rice bran |
| GLP-1 | Glucagon-like peptide 1 |
| HDL-C | High-density lipoprotein-cholesterol |
| HMGR | 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase |
| LDL-C | Low-density lipoprotein-cholesterol |
| LPH | Lentil protein hydrolysate |
| oxLDL | Oxidative products of LDL |
| p27Kip | Cyclin-dependent kinase inhibitor 1B |
| PHA | Phytohaemagglutinin |
| PCSK9 | Proprotein convertase subtilisin/kexin type 9 |
| RPI | Rice protein isolate |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| SCFA | Short-chain fatty acids |
| SHR | Spontaneously hypertensive rats |
| SREBP | Sterol regulatory element-binding protein |
| SSP | Seed storage proteins |
| TC | Total cholesterol |
| TG | Triglyceride |
| T2D | Type 2 diabetes |
| TRB | Thermolysin-digested rice bran |
| VLDL-C | Very-low-density lipoprotein-cholesterol |
References
- Salas-Salvadó, J.; Guasch-Ferré, M.; Lee, C.-H.; Estruch, R.; Clish, C.B.; Ros, E. Protective Effects of the Mediterranean Diet on Type 2 Diabetes and Metabolic Syndrome. J. Nutr. 2016, 146, 920S–927S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRae, M.P. Health Benefits of Dietary Whole Grains: An Umbrella Review of Meta-analyses. J. Chiropr. Med. 2017, 16, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, C.; Hillen, C.; Garden Robinson, J. Composition, Nutritional Value, and Health Benefits of Pulses. Cereal Chem. 2017, 94, 11–31. [Google Scholar] [CrossRef]
- Jones, J.M.; García, C.G.; Braun, H.J. Perspective: Whole and Refined Grains and Health-Evidence Supporting “Make Half Your Grains Whole”. Adv. Nutr. 2020, 11, 492–506. [Google Scholar] [CrossRef] [Green Version]
- Marinangeli, C.P.; Jones, P.J. Pulse grain consumption and obesity: Effects on energy expenditure, substrate oxidation, body composition, fat deposition and satiety. Br. J. Nutr. 2012, 108 (Suppl. 1), S46–S51. [Google Scholar] [CrossRef]
- Jones, J.M.; Peña, R.J.; Korczak, R.; Braun, H.J. Wheat, Grains, and Health: Carbohydrates, Grains, and Whole Grains and Disease Prevention. Part I. Body Weight and Obesity. Cereal Foods World 2016, 61, 96–105. [Google Scholar] [CrossRef]
- Singhal, P.; Kaushik, G. Therapeutic Effect of Cereal Grains: A Review. Crit. Rev. Food Sci. Nutr. 2016, 56, 748–759. [Google Scholar] [CrossRef]
- Ramdath, D.; Renwick, S.; Duncan, A.M. The Role of Pulses in the Dietary Management of Diabetes. Can. J. Diabetes 2016, 40, 355–363. [Google Scholar] [CrossRef]
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Whole grain consumption and risk of cardiovascular disease, cancer, and all cause and cause specific mortality: Systematic review and dose-response meta-analysis of prospective studies. BMJ 2016, 353, i2716. [Google Scholar] [CrossRef] [Green Version]
- Padhi, E.M.T.; Ramdath, D.D. A review of the relationship between pulse consumption and reduction of cardiovascular disease risk factors. J. Funct. Foods 2017, 38, 635–643. [Google Scholar] [CrossRef]
- Rao, S.; Chinkwo, K.A.; Santhakumar, A.B.; Blanchard, C.L. Inhibitory Effects of Pulse Bioactive Compounds on Cancer Development Pathways. Diseases 2018, 6, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Yang, J.; Du, L.; Li, K.; Zhou, Y. Association of whole grain, refined grain, and cereal consumption with gastric cancer risk: A meta-analysis of observational studies. Food Sci. Nutr. 2019, 7, 256–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryngelsson, S.; Mannerstedt-Fogelfors, B.; Kamal-Eldin, A.; Andersson, R.; Dimberg, L.H. Lipids and antioxidants in groats and hulls of Swedish oats (Avena sativa L). J. Sci. Food Agric. 2002, 82, 606–614. [Google Scholar] [CrossRef]
- Seal, C.J. Whole Grains. In Encyclopedia of Human Nutrition, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 422–439. [Google Scholar]
- Evers, T.; Millar, S. Cereal Grain Structure and Development: Some Implications for Quality. J. Cereal Sci. 2002, 36, 261–284. [Google Scholar] [CrossRef]
- Saulnier, L.; Guillon, F.; Sado, P.E.; Rouau, X. Plant Cell Wall Polysaccharides in Storage Organs: Xylans (Food Applications). Compr. Glycosci. 2007, 2, 653–689. [Google Scholar]
- Zitterman, A. DIETARY FIBER|Bran. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 1844–1850. [Google Scholar]
- Pulse Canada. What Is a Pulse? Available online: http://www.pulsecanada.com/about-pulse-canada/what-is-a-pulse/ (accessed on 17 July 2020).
- Food and Agriculture Organization of the United Nations. International Year of Pulses 2016. Available online: http://www.fao.org/pulses-2016/en/ (accessed on 17 July 2020).
- Chibbar, R.N.; Ambigaipalan, P.; Hoover, R. REVIEW: Molecular Diversity in Pulse Seed Starch and Complex Carbohydrates and Its Role in Human Nutrition and Health. Cereal Chem. 2010, 87, 342–352. [Google Scholar] [CrossRef]
- Singh, N. Pulses: An overview. J. Food Sci. Technol. 2017, 54, 853–857. [Google Scholar] [CrossRef] [Green Version]
- Barker, B. Understanding Protein in Pulses; Growers, S.P., Ed.; Saskatchewan Pulse Growers: Saskatoon, SK, Canada, 2019. [Google Scholar]
- Singh, B.; Singh, J.P.; Shevkani, K.; Singh, N.; Kaur, A. Bioactive constituents in pulses and their health benefits. J. Food Sci. Technol. 2017, 54, 858–870. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Singh, J.P.; Singh, N.; Kaur, A. Saponins in pulses and their health promoting activities: A review. Food Chem. 2017, 233, 540–549. [Google Scholar] [CrossRef]
- Rochfort, S.; Panozzo, J. Phytochemicals for Health, the Role of Pulses. J. Agric. Food Chem. 2007, 55, 7981–7994. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, J.; Ilic, S.; Jun Xue, S.; Kakuda, Y. Biological Properties and Characterization of Lectin from Red Kidney Bean (Phaseolus vulgaris). Food Rev. Int. 2008, 25, 12–27. [Google Scholar] [CrossRef]
- Mudryj, A.N.; Yu, N.; Aukema, H.M. Nutritional and health benefits of pulses. Appl. Physiol. Nutr. Metab. 2014, 39, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Joye, I. Protein Digestibility of Cereal Products. Foods 2019, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A. Proteins in Pulses. J. Nutr. Disord. Ther. 2017, 7, e129. [Google Scholar] [CrossRef]
- Clark, L.; Bogdan, A. Plant-based foods in Canada: Information, trust and closing the commercialization gap. Br. Food J. 2019, 121, 2535–2550. [Google Scholar] [CrossRef]
- Canada’s Food Guide, Healthy Food Choices. 2020. Available online: https://food-guide.canada.ca/en/healthy-food-choices/ (accessed on 20 May 2022).
- Gharibzahedi, S.M.T.; Smith, B. Legume proteins are smart carriers to encapsulate hydrophilic and hydrophobic bioactive compounds and probiotic bacteria: A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1250–1279. [Google Scholar] [CrossRef] [PubMed]
- Cavazos, A.; Gonzalez de Mejia, E. Identification of Bioactive Peptides from Cereal Storage Proteins and Their Potential Role in Prevention of Chronic Diseases. Compr. Rev. Food Sci. Food Saf. 2013, 12, 364–380. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive Peptides: Synthesis, Sources, Applications, and Proposed Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Zhuang, Y.; Li, Y.; Shi, P.; Tian, H.; Li, X.; Chen, X. Isolation of novel ACE-inhibitory peptide from naked oat globulin hydrolysates in silico approach: Molecular docking, in vivo antihypertension and effects on renin and intracellular endothelin-1. J. Food Sci. 2020, 85, 1328–1337. [Google Scholar] [CrossRef]
- Cheung, I.W.Y.; Nakayama, S.; Hsu, M.N.K.; Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Angiotensin-I Converting Enzyme Inhibitory Activity of Hydrolysates from Oat (Avena sativa) Proteins by In Silico and In Vitro Analyses. J. Agric. Food Chem. 2009, 57, 9234–9242. [Google Scholar] [CrossRef] [PubMed]
- Gangopadhyay, N.; Wynne, K.; O’Connor, P.; Gallagher, E.; Brunton, N.P.; Rai, D.K.; Hayes, M. In Silico and In Vitro analyses of the angiotensin-I converting enzyme inhibitory activity of hydrolysates generated from crude barley (Hordeum vulgare) protein concentrates. Food Chem. 2016, 203, 367–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, F.; Chen, L.; Liang, R.; Zhang, Z.; Wang, J.; Cai, M.; Li, Y. Pilot-scale production of low molecular weight peptides from corn wet milling byproducts and the antihypertensive effects in vivo and in vitro. Food Chem. 2011, 124, 801–807. [Google Scholar] [CrossRef]
- Suh, H.J.; Whang, J.H.; Lee, H. A peptide from corn gluten hydrolysate that is inhibitory toward angiotensin I converting enzyme. Biotechnol. Lett. 1999, 21, 1055–1058. [Google Scholar] [CrossRef]
- Karami, Z.; Peighambardoust, S.H.; Hesari, J.; Akbari-Adergani, B.; Andreu, D. Antioxidant, anticancer and ACE-inhibitory activities of bioactive peptides from wheat germ protein hydrolysates. Food Biosci. 2019, 32, 100450. [Google Scholar] [CrossRef]
- Motoi, H.; Kodama, T. Isolation and characterization of angiotensin I-converting enzyme inhibitory peptides from wheat gliadin hydrolysate. Food/Nahrung 2003, 47, 354–358. [Google Scholar] [CrossRef]
- Chen, J.; Liu, S.; Ye, R.; Cai, G.; Ji, B.; Wu, Y. Angiotensin-I converting enzyme (ACE) inhibitory tripeptides from rice protein hydrolysate: Purification and characterization. J. Funct. Foods 2013, 5, 1684–1692. [Google Scholar] [CrossRef]
- Wang, X.; Chen, H.; Fu, X.; Li, S.; Wei, J. A novel antioxidant and ACE inhibitory peptide from rice bran protein: Biochemical characterization and molecular docking study. LWT 2017, 75, 93–99. [Google Scholar] [CrossRef]
- Suwannapan, O.; Wachirattanapongmetee, K.; Thawornchinsombut, S.; Katekaew, S. Angiotensin-I-converting enzyme (ACE)-inhibitory peptides from Thai jasmine rice bran protein hydrolysates. Int. J. Food Sci. Technol. 2020, 55, 2441–2450. [Google Scholar] [CrossRef]
- Shobako, N.; Ogawa, Y.; Ishikado, A.; Harada, K.; Kobayashi, E.; Suido, H.; Kusakari, T.; Maeda, M.; Suwa, M.; Matsumoto, M.; et al. A Novel Antihypertensive Peptide Identified in Thermolysin-Digested Rice Bran. Mol. Nutr. Food Res. 2018, 62, 1700732. [Google Scholar] [CrossRef]
- Jeong, H.J.; Jeong, J.B.; Hsieh, C.C.; Hernández-Ledesma, B.; de Lumen, B.O. Lunasin Is Prevalent in Barley and Is Bioavailable and Bioactive in In Vivo and In Vitro Studies. Nutr. Cancer 2010, 62, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Connolly, A.; O’Keeffe, M.B.; Nongonierma, A.B.; Piggott, C.O.; FitzGerald, R.J. Isolation of peptides from a novel brewers spent grain protein isolate with potential to modulate glycaemic response. Int. J. Food Sci. Technol. 2017, 52, 146–153. [Google Scholar] [CrossRef]
- Ramírez Fuentes, L.; Richard, C.; Chen, L. Sequential alcalase and flavourzyme treatment for preparation of α-amylase, α-glucosidase, and dipeptidyl peptidase (DPP)-IV inhibitory peptides from oat protein. J. Funct. Foods 2021, 87, 104829. [Google Scholar] [CrossRef]
- Wang, F.; Yu, G.; Zhang, Y.; Zhang, B.; Fan, J. Dipeptidyl Peptidase IV Inhibitory Peptides Derived from Oat (Avena sativa L.), Buckwheat (Fagopyrum esculentum), and Highland Barley (Hordeum vulgare trifurcatum (L.) Trofim) Proteins. J. Agric. Food Chem. 2015, 63, 9543–9549. [Google Scholar] [CrossRef] [PubMed]
- Kannan, A.; Hettiarachchy, N.S.; Lay, J.O.; Liyanage, R. Human cancer cell proliferation inhibition by a pentapeptide isolated and characterized from rice bran. Peptides 2010, 31, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Kannan, A.; Hettiarachchy, N.S.; Mahedevan, M. Peptides Derived from Rice Bran Protects Cells from Obesity and Alzheimer’s Disease. Int. J. Biomed. Res. 2012, 3, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, H.; Joshi, R.; Gupta, M. Purification, identification and characterization of two novel antioxidant peptides from finger millet (Eleusine coracana) protein hydrolysate. Food Res. Int. 2019, 120, 697–707. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, H.; Wang, X.; Li, S.; Chen, Z.; Wang, J.; Liu, W. Isolation and identification of a novel peptide from zein with antioxidant and antihypertensive activities. Food Funct. 2015, 6, 3799–3806. [Google Scholar] [CrossRef]
- Yust Mdel, M.; Millán-Linares Mdel, C.; Alcaide-Hidalgo, J.M.; Millán, F.; Pedroche, J. Hypocholesterolaemic and antioxidant activities of chickpea (Cicer arietinum L.) protein hydrolysates. J. Sci. Food Agric. 2012, 92, 1994–2001. [Google Scholar] [CrossRef]
- Li, G.-H.; Wan, J.-Z.; Le, G.-W.; Shi, Y.-H. Novel angiotensin I-converting enzyme inhibitory peptides isolated from Alcalase hydrolysate of mung bean protein. J. Pept. Sci. 2006, 12, 509–514. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Baraniak, B. Activities and sequences of the angiotensin I-converting enzyme (ACE) inhibitory peptides obtained from the digested lentil (Lens culinaris) globulins. Int. J. Food Sci. Technol. 2013, 48, 2363–2369. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Baraniak, B. Angiotensin I converting enzyme inhibitory peptides obtained after in vitro hydrolysis of pea (Pisum sativum var. Bajka) globulins. BioMed Res. Int. 2014, 2014, 438459. [Google Scholar] [CrossRef] [Green Version]
- Mojica, L.; Luna-Vital, D.A.; González de Mejía, E. Characterization of peptides from common bean protein isolates and their potential to inhibit markers of type-2 diabetes, hypertension and oxidative stress. J. Sci. Food Agric. 2017, 97, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Luna Vital, D.A.; González de Mejía, E.; Dia, V.P.; Loarca-Piña, G. Peptides in common bean fractions inhibit human colorectal cancer cells. Food Chem. 2014, 157, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Alvarez, S.; Majumder, K. Transport of Dietary Anti-Inflammatory Peptide, γ-Glutamyl Valine (γ-EV), across the Intestinal Caco-2 Monolayer. Nutrients 2021, 13, 1448. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Expósito, S.; Martinez-Villaluenga, C.; Dueñas, M.; Silvan, J.; Frias, J.; Peñas, E. Combination of pH-controlled fermentation in mild acidic conditions and enzymatic hydrolysis by Savinase to improve metabolic health-promoting properties of lentil. J. Funct. Foods 2018, 48, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Finkina, E.I.; Shramova, E.I.; Tagaev, A.A.; Ovchinnikova, T.V. A novel defensin from the lentil Lens culinaris seeds. Biochem. Biophys. Res. Commun. 2008, 371, 860–865. [Google Scholar] [CrossRef]
- Xue, Z.; Wang, R.; Wen, H.; Yu, W.; Kou, X. Biochanin A and CPe-III Peptide Improved Hepatic Inflammation by Regulating the Hepatic Lipid Metabolic Pathways in Diet-Induced Obese Mice. Trans. Tianjin Univ. 2018, 24, 234–243. [Google Scholar] [CrossRef]
- Shi, W.; Hou, T.; Guo, D.; He, H. Evaluation of hypolipidemic peptide (Val-Phe-Val-Arg-Asn) virtual screened from chickpea peptides by pharmacophore model in high-fat diet-induced obese rat. J. Funct. Foods 2019, 54, 136–145. [Google Scholar] [CrossRef]
- Zanoni, C.; Aiello, G.; Arnoldi, A.; Lammi, C. Investigations on the hypocholesterolaemic activity of LILPKHSDAD and LTFPGSAED, two peptides from lupin β-conglutin: Focus on LDLR and PCSK9 pathways. J. Funct. Foods 2017, 32, 1–8. [Google Scholar] [CrossRef]
- Lammi, C.; Fassi, E.M.A.; Li, J.; Bartolomei, M.; Benigno, G.; Roda, G.; Arnoldi, A.; Grazioso, G. Computational Design and Biological Evaluation of Analogs of Lupin Peptide P5 Endowed with Dual PCSK9/HMG-CoAR Inhibiting Activity. Pharmaceutics 2022, 14, 665. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Tomar, M.; Potkule, J.; Verma, R.; Punia, S.; Mahapatra, A.; Belwal, T.; Dahuja, A.; Joshi, S.; Berwal, M.K.; et al. Advances in the plant protein extraction: Mechanism and recommendations. Food Hydrocoll. 2021, 115, 106595. [Google Scholar] [CrossRef]
- Sari, Y.W.; Mulder, W.J.; Sanders, J.P.M.; Bruins, M.E. Towards plant protein refinery: Review on protein extraction using alkali and potential enzymatic assistance. Biotechnol. J. 2015, 10, 1138–1157. [Google Scholar] [CrossRef] [PubMed]
- Osborne, T.B. The Vegetable Proteins, 2nd ed.; Longmans Green and Co.: London, UK, 1924. [Google Scholar]
- Boye, J.; Zare, F.; Pletch, A. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- González-Pérez, S.; Arellano, J.B. Vegetable protein isolates. In Handbook of Hydrocolloids, 2nd ed.; Phillips, G.O., Williams, P.A., Eds.; Woodhead Publishing: Sawston, UK, 2009; pp. 383–419. [Google Scholar]
- Krishnan, H.B.; Coe, E.H. Seed Storage Proteins. In Encyclopedia of Genetics; Brenner, S., Miller, J.H., Eds.; Academic Press: New York, NY, USA, 2001; pp. 1782–1787. [Google Scholar]
- Duranti, M. Grain legume proteins and nutraceutical properties. Fitoterapia 2006, 77, 67–82. [Google Scholar] [CrossRef] [PubMed]
- O’Kane, F.E.; Happe, R.P.; Vereijken, J.M.; Gruppen, H.; van Boekel, M.A.J.S. Characterization of Pea Vicilin. 1. Denoting Convicilin as the α-Subunit of the Pisum Vicilin Family. J. Agric. Food Chem. 2004, 52, 3141–3148. [Google Scholar] [CrossRef]
- Koehler, P.; Wieser, H. Chemistry of Cereal Grains. In Handbook on Sourdough Biotechnology; Gobbetti, M., Gänzle, M., Eds.; Springer: Boston, MA, USA, 2013; pp. 11–45. [Google Scholar]
- Yadav, D.; Singh, N. Wheat Triticin: A Potential Target for Nutritional Quality Improvement. Asian J. Biotechnol. 2011, 3, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Han, F.; Moughan, P.J.; Li, J.; Stroebinger, N.; Pang, S. The Complementarity of Amino Acids in Cooked Pulse/Cereal Blends and Effects on DIAAS. Plants 2021, 10, 1999. [Google Scholar] [CrossRef]
- Pulse Canada. PASTA & NOODLES. Available online: https://pulsecanada.com/applications/pasta-noodles (accessed on 17 May 2022).
- Sopiwnyk, E.; Bourré, L.; Young, G.; Borsuk, Y.; Lagassé, S.; Boyd, L.; Sarkar, A.; Jones, S.; Dyck, A.; Malcolmson, L. Flour and bread making properties of whole and split yellow peas treated with dry and steam heat used as premilling treatment. Cereal Chem. 2020, 97, 1290–1302. [Google Scholar] [CrossRef]
- Wu, Y.V.; Wall, J.S. Lysine content of protein increased by germination of normal and high-lysine sorghums. J. Agric. Food Chem. 1980, 28, 455–458. [Google Scholar] [CrossRef]
- Craddock, J.C.; Genoni, A.; Strutt, E.F.; Goldman, D.M. Limitations with the Digestible Indispensable Amino Acid Score (DIAAS) with Special Attention to Plant-Based Diets: A Review. Curr. Nutr. Rep. 2021, 10, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, N.S.; Bailey, H.M.; Guardiola, L.V.; Stein, H.H. Values for Digestible Indispensable Amino Acid Score (DIAAS) Determined in Pigs Are Greater for Milk Than for Breakfast Cereals, but DIAAS Values for Individual Ingredients Are Additive in Combined Meals. J. Nutr. 2021, 151, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-J.; Hao, J.; Kim, C.-H.; Hong, J.-S.; Ahn, H.-Y.; Lee, Y.-S. Quercetin Prevents Cardiac Hypertrophy Induced by Pressure Overload in Rats. J. Vet. Med. Sci. 2009, 71, 737–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herreman, L.; Nommensen, P.; Pennings, B.; Laus, M.C. Comprehensive overview of the quality of plant- And animal-sourced proteins based on the digestible indispensable amino acid score. Food Sci. Nutr. 2020, 8, 5379–5391. [Google Scholar] [CrossRef]
- Rutherfurd, S.M.; Fanning, A.C.; Miller, B.J.; Moughan, P.J. Protein Digestibility-Corrected Amino Acid Scores and Digestible Indispensable Amino Acid Scores Differentially Describe Protein Quality in Growing Male Rats. J. Nutr. 2015, 145, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Cervantes-Pahm, S.K.; Liu, Y.; Stein, H.H. Digestible indispensable amino acid score and digestible amino acids in eight cereal grains. Br. J. Nutr. 2014, 111, 1663–1672. [Google Scholar] [CrossRef]
- Nitrayová, S.; Brestenský, M.; Patráš, P. Comparison of two methods of protein quality evaluation in rice, rye and barley as food protein sources in human nutrition. Potravin. Slovak J. Food Sci. 2018, 12, 762–766. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.M. Current Concepts and Unresolved Questions in Dietary Protein Requirements and Supplements in Adults. Front. Nutr. 2017, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Mathai, J. Digestible Indispensable Amino Acid Scores for Food Proteins. Ph.D. Thesis, University of Illinois, Urbana, IL, USA, 2018. [Google Scholar]
- Nosworthy, M.G.; Neufeld, J.; Frohlich, P.; Young, G.; Malcolmson, L.; House, J.D. Determination of the protein quality of cooked Canadian pulses. Food Sci. Nutr. 2017, 5, 896–903. [Google Scholar] [CrossRef]
- Calderón de la Barca, A.M.; Martínez-Díaz, G.; Ibarra-Pastrana, É.N.; Devi, S.; Kurpad, A.V.; Valencia, M.E. Pinto Bean Amino Acid Digestibility and Score in a Mexican Dish with Corn Tortilla and Guacamole, Evaluated in Adults Using a Dual-Tracer Isotopic Method. J. Nutr. 2021, 151, 3151–3157. [Google Scholar] [CrossRef]
- Udenigwe, C.C. Bioinformatics approaches, prospects and challenges of food bioactive peptide research. Trends Food Sci. Technol. 2014, 36, 137–143. [Google Scholar] [CrossRef]
- FitzGerald, R.J.; O’Cuinn, G. Enzymatic debittering of food protein hydrolysates. Biotechnol. Adv. 2006, 24, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xie, N.; Li, B. Influence of peptide characteristics on their stability, intestinal transport, and in vitro bioavailability: A review. J. Food Biochem. 2019, 43, e12571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakhra, V.; Galappaththy, S.L.; Bulchandani, S.; Cabandugama, P.K. Obesity and the Western Diet: How We Got Here. Mo. Med. 2020, 117, 536–538. [Google Scholar]
- Kanoski, S.E.; Hsu, T.M.; Pennell, S. Chapter 5—Obesity, Western Diet Intake, and Cognitive Impairment. In Omega-3 Fatty Acids in Brain and Neurological Health; Watson, R.R., De Meester, F., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 57–62. [Google Scholar]
- Chaudhary, A.; Marinangeli, C.P.F.; Tremorin, D.; Mathys, A. Nutritional Combined Greenhouse Gas Life Cycle Analysis for Incorporating Canadian Yellow Pea into Cereal-Based Food Products. Nutrients 2018, 10, 490. [Google Scholar] [CrossRef] [Green Version]
- Rebello, C.J.; Greenway, F.L.; Finley, J.W. A review of the nutritional value of legumes and their effects on obesity and its related co-morbidities. Obes. Rev. 2014, 15, 392–407. [Google Scholar] [CrossRef]
- Geraedts, M.C.P.; Troost, F.J.; Tinnemans, R.; Söderholm, J.D.; Brummer, R.-J.; Saris, W.H.M. Release of Satiety Hormones in Response to Specific Dietary Proteins Is Different between Human and Murine Small Intestinal Mucosa. Ann. Nutr. Metab. 2010, 56, 308–313. [Google Scholar] [CrossRef]
- Abete, I.; Parra, D.; Martinez, J.A. Legume-, Fish-, or High-Protein-Based Hypocaloric Diets: Effects on Weight Loss and Mitochondrial Oxidation in Obese Men. J. Med. Food 2009, 12, 100–108. [Google Scholar] [CrossRef]
- Iwashita, S.; Mikus, C.; Baier, S.; Flakoll, P.J. Glutamine Supplementation Increases Postprandial Energy Expenditure and Fat Oxidation in Humans. J. Parenter. Enter. Nutr. 2006, 30, 76–80. [Google Scholar] [CrossRef]
- Pusztai, A.; Grant, G.; Buchan, W.C.; Bardocz, S.; de Carvalho, A.F.F.U.; Ewen, S.W.B. Lipid accumulation in obese Zucker rats is reduced by inclusion of raw kidney bean (Phaseolus vulgaris) in the diet. Br. J. Nutr. 1998, 79, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Herzig, K.H.; Bardocz, S.; Grant, G.; Nustede, R.; Fölsch, U.R.; Pusztai, A. Red kidney bean lectin is a potent cholecystokinin releasing stimulus in the rat inducing pancreatic growth. Gut 1997, 41, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Spadafranca, A.; Rinelli, S.; Riva, A.; Morazzoni, P.; Magni, P.; Bertoli, S.; Battezzati, A. Phaseolus vulgaris extract affects glycometabolic and appetite control in healthy human subjects. Br. J. Nutr. 2013, 109, 1789–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, M.L.; Udani, J.K. A proprietary alpha-amylase inhibitor from white bean (Phaseolus vulgaris): A review of clinical studies on weight loss and glycemic control. Nutr. J. 2011, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udani, J.; Tan, O.; Molina, J. Systematic Review and Meta-Analysis of a Proprietary Alpha-Amylase Inhibitor from White Bean (Phaseolus vulgaris L.) on Weight and Fat Loss in Humans. Foods 2018, 7, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Han, G.; Liu, Q.H.; Wu, Q.; He, H.J.; Cheng, C.Z.; Duan, Y.J. Rice protein exerts a hypocholesterolemic effect through regulating cholesterol metabolism-related gene expression and enzyme activity in adult rats fed a cholesterol-enriched diet. Int. J. Food Sci. Nutr. 2013, 64, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, Y.; Hosojima, M.; Kabasawa, H.; Kuwahara, S.; Goto, S.; Toba, K.; Kaseda, R.; Tanaka, T.; Kitamura, N.; Takihara, H.; et al. Rice Endosperm Protein Administration to Juvenile Mice Regulates Gut Microbiota and Suppresses the Development of High-Fat Diet-Induced Obesity and Related Disorders in Adulthood. Nutrients 2019, 11, 2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statistics Canada. Cholesterol Levels of Adults, 2016–2019; 2021. Available online: https://www150.statcan.gc.ca/n1/pub/82-625-x/2021001/article/00003-eng.htm (accessed on 29 June 2021).
- Hollænder, P.L.B.; Ross, A.B.; Kristensen, M. Whole-grain and blood lipid changes in apparently healthy adults: A systematic review and meta-analysis of randomized controlled studies 1–3. Am. J. Clin. Nutr. 2015, 102, 556–572. [Google Scholar] [CrossRef] [Green Version]
- Health Canada Oat Products and Blood Cholesterol Lowering. Available online: https://www.canada.ca/en/health-canada/services/food-nutrition/food-labelling/health-claims/assessments/products-blood-cholesterol-lowering-summary-assessment-health-claim-about-products-blood-cholesterol-lowering.html (accessed on 27 August 2020).
- Health Canada Summary of Health Canada’s Assessment of a Health Claim about Barley Products and Blood Cholesterol Lowering. Available online: https://www.canada.ca/en/health-canada/services/food-nutrition/food-labelling/health-claims/assessments/assessment-health-claim-about-barley-products-blood-cholesterol-lowering.html (accessed on 27 August 2020).
- Liang, T.T.; Tong, L.T.; Geng, D.H.; Wang, L.L.; Zhou, X.R.; Pu, H.Y.; Jia, W.; Wu, Q.P.; Huang, J.R. Wheat Gluten Regulates Cholesterol Metabolism by Modulating Gut Microbiota in Hamsters with Hyperlipidemia. J. Oleo Sci. 2019, 68, 909–922. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, J.; Liu, Y.; Gong, L.; Sun, B. Rice bran proteins and their hydrolysates modulate cholesterol metabolism in mice on hypercholesterolemic diets. Food Funct. 2016, 7, 2747–2753. [Google Scholar] [CrossRef]
- Boualga, A.; Prost, J.; Taleb-Senouci, D.; Krouf, D.; Kharoubi, O.; Lamri-Senhadji, M.; Belleville, J.; Bouchenak, M. Purified chickpea or lentil proteins impair VLDL metabolism and lipoprotein lipase activity in epididymal fat, but not in muscle, compared to casein, in growing rats. Eur. J. Nutr. 2009, 48, 162–169. [Google Scholar] [CrossRef]
- Barbana, C.; Boucher, A.C.; Boye, J.I. In vitro binding of bile salts by lentil flours, lentil protein concentrates and lentil protein hydrolysates. Food Res. Int. 2011, 44, 174–180. [Google Scholar] [CrossRef]
- Martínez, R.; Kapravelou, G.; Donaire, A.; Lopez-Chaves, C.; Arrebola, F.; Galisteo, M.; Cantarero, S.; Aranda, P.; Porres, J.M.; López-Jurado, M. Effects of a combined intervention with a lentil protein hydrolysate and a mixed training protocol on the lipid metabolism and hepatic markers of NAFLD in Zucker rats. Food Funct. 2018, 9, 830–850. [Google Scholar] [CrossRef] [PubMed]
- Yahia, S.; Benomar, S.; Dehiba, F.; Allaoui, A.; Guillén, N.; Rodríguez-Yoldi, M.; Osada, J.; Boualga, A.J.N.; Science, F. Hypocholesterolaemic and antioxidant efficiency of chickpea (Cicer arietinum) protein hydrolysates depend on its degree of hydrolysis in cholesterol-fed rat. Nutr. Food Sci. 2017, 47, 254–269. [Google Scholar] [CrossRef]
- Amaral, A.; Ferreira, E.; Neves, V.; Demonte, A. Legumin from chickpea: Hypolipidemic effect in the liver of hypercholesterolemic rats. Nutr. Food Sci. 2014, 44, 378–388. [Google Scholar] [CrossRef]
- Shrestha, S.; Hag, L.V.T.; Haritos, V.S.; Dhital, S. Lupin proteins: Structure, isolation and application. Trends Food Sci. Technol. 2021, 116, 928–939. [Google Scholar] [CrossRef]
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; Álvarez-Ríos, A.I.; Santos-Sánchez, G.; Pedroche, J.; Millán, F.; Carrera Sánchez, C.; Fernández-Pachón, M.S.; Millán-Linares, M.C.; Martínez-López, A.; et al. Safety and Efficacy of a Beverage Containing Lupine Protein Hydrolysates on the Immune, Oxidative and Lipid Status in Healthy Subjects: An Intervention Study (the Lupine-1 Trial). Mol. Nutr. Food Res. 2021, 65, e2100139. [Google Scholar] [CrossRef] [PubMed]
- Bähr, M.; Fechner, A.; Kiehntopf, M.; Jahreis, G. Consuming a mixed diet enriched with lupin protein beneficially affects plasma lipids in hypercholesterolemic subjects: A randomized controlled trial. Clin. Nutr. 2015, 34, 7–14. [Google Scholar] [CrossRef]
- Bähr, M.; Fechner, A.; Krämer, J.; Kiehntopf, M.; Jahreis, G. Lupin protein positively affects plasma LDL cholesterol and LDL:HDL cholesterol ratio in hypercholesterolemic adults after four weeks of supplementation: A randomized, controlled crossover study. Nutr. J. 2013, 12, 107. [Google Scholar] [CrossRef] [Green Version]
- Pavanello, C.; Lammi, C.; Ruscica, M.; Bosisio, R.; Mombelli, G.; Zanoni, C.; Calabresi, L.; Sirtori, C.R.; Magni, P.; Arnoldi, A. Effects of a lupin protein concentrate on lipids, blood pressure and insulin resistance in moderately dyslipidaemic patients: A randomised controlled trial. J. Funct. Foods 2017, 37, 8–15. [Google Scholar] [CrossRef]
- Druce, I.; Abujrad, H.; Ooi, T.C. PCSK9 and triglyceride-rich lipoprotein metabolism. J. Biomed. Res. 2015, 29, 429–436. [Google Scholar]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Aiello, G. YDFYPSSTKDQQS (P3), a peptide from lupin protein, absorbed by Caco-2 cells, modulates cholesterol metabolism in HepG2 cells via SREBP-1 activation. J. Food Biochem. 2019, 43, e12757. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Chen, J.-H.; Lv, J.; Wu, Q.; Xu, T.; Zhang, H.; Liu, Q.-H.; Yang, H.-K. Rice protein improves adiposity, body weight and reduces lipids level in rats through modification of triglyceride metabolism. Lipids Health Dis. 2012, 11, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronis, M.J.; Badeaux, J.; Chen, Y.; Badger, T.M. Rice protein isolate improves lipid and glucose homeostasis in rats fed high fat/high cholesterol diets. Exp. Biol. Med. 2010, 235, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Um, M.Y.; Ahn, J.; Jung, C.H.; Ha, T.Y. Cholesterol-lowering Effect of Rice Protein by Enhancing Fecal Excretion of Lipids in Rats. Prev. Nutr. Food Sci. 2013, 18, 210–213. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Yang, L.; He, H.J.; Xu, T.; Liu, H.B.; Wu, Q.; Ma, Y.; Liu, Q.H.; Nie, M.H. Antioxidant capacity responsible for a hypocholesterolemia is independent of dietary cholesterol in adult rats fed rice protein. Gene 2014, 533, 57–66. [Google Scholar] [CrossRef]
- Hosojima, M.; Kaseda, R.; Kondo, H.; Fujii, M.; Kubota, M.; Watanabe, R.; Tanabe, N.; Kadowaki, M.; Suzuki, Y.; Saito, A. Beneficial effects of rice endosperm protein intake in Japanese men with risk factors for metabolic syndrome: A randomized, crossover clinical trial. BMC Nutr. 2016, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, D.J.; Kendall, C.W.; Vuksan, V.; Augustin, L.S.; Mehling, C.; Parker, T.; Vidgen, E.; Lee, B.; Faulkner, D.; Seyler, H.; et al. Effect of wheat bran on serum lipids: Influence of particle size and wheat protein. J. Am. Coll. Nutr. 1999, 18, 159–165. [Google Scholar] [CrossRef]
- Guo, L.; Tong, L.T.; Liu, L.; Zhong, K.; Qiu, J.; Zhou, S. The cholesterol-lowering effects of oat varieties based on their difference in the composition of proteins and lipids. Lipids Health Dis. 2014, 13, 182. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.T.; Guo, L.; Zhou, X.; Qiu, J.; Liu, L.; Zhong, K.; Zhou, S. Effects of dietary oat proteins on cholesterol metabolism of hypercholesterolaemic hamsters. J. Sci. Food Agric. 2016, 96, 1396–1401. [Google Scholar] [CrossRef]
- Deshpande, A.; Harris-Hayes, M.; Schootman, M. Epidemiology of Diabetes and Diabetes-Related Complications. Phys. Ther. 2008, 88, 1254–1264. [Google Scholar] [CrossRef] [Green Version]
- Aune, D.; Norat, T.; Romundstad, P.; Vatten, L.J. Whole grain and refined grain consumption and the risk of type 2 diabetes: A systematic review and dose–response meta-analysis of cohort studies. Eur. J. Epidemiol. 2013, 28, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, T.; Inoue, Y.; Arima, J.; Kumagai, Y.; Usuki, H.; Kawakami, K.; Kimura, M.; Mukaihara, T. Production of dipeptidyl peptidase IV inhibitory peptides from defatted rice bran. Food Chem. 2012, 134, 797–802. [Google Scholar] [CrossRef]
- Zhu, B.; He, H.; Hou, T. A Comprehensive Review of Corn Protein-derived Bioactive Peptides: Production, Characterization, Bioactivities, and Transport Pathways. Compr. Rev. Food Sci. Food Saf. 2019, 18, 329–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, S.; Luna-Vital, D.; de Mejia, E.G. Identification and Comparison of Peptides from Chickpea Protein Hydrolysates Using Either Bromelain or Gastrointestinal Enzymes and Their Relationship with Markers of Type 2 Diabetes and Bitterness. Nutrients 2020, 12, 3843. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Peptides Derived from Soy and Lupin Protein as Dipeptidyl-Peptidase IV Inhibitors: In Vitro Biochemical Screening and in Silico Molecular Modeling Study. J. Agric. Food Chem. 2016, 64, 9601–9606. [Google Scholar] [CrossRef] [Green Version]
- Mojica, L.; Chen, K.; de Mejía, E.G. Impact of Commercial Precooking of Common Bean (Phaseolus vulgaris) on the Generation of Peptides, After Pepsin–Pancreatin Hydrolysis, Capable to Inhibit Dipeptidyl Peptidase-IV. J. Food Sci. 2015, 80, H188–H198. [Google Scholar] [CrossRef]
- Magni, C.; Sessa, F.; Accardo, E.; Vanoni, M.; Morazzoni, P.; Scarafoni, A.; Duranti, M. Conglutin γ, a lupin seed protein, binds insulin in vitro and reduces plasma glucose levels of hyperglycemic rats. J. Nutr. Biochem. 2004, 15, 646–650. [Google Scholar] [CrossRef]
- Vargas-Guerrero, B.; García-López, P.M.; Martínez-Ayala, A.L.; Domínguez-Rosales, J.A.; Gurrola-Díaz, C.M. Administration of Lupinus albus Gamma Conglutin (Cγ) to n5 STZ Rats Augmented Ins-1 Gene Expression and Pancreatic Insulin Content. Plant Foods Hum. Nutr. 2014, 69, 241–247. [Google Scholar] [CrossRef]
- Terruzzi, I.; Senesi, P.; Magni, C.; Montesano, A.; Scarafoni, A.; Luzi, L.; Duranti, M. Insulin-mimetic action of conglutin-γ, a lupin seed protein, in mouse myoblasts. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 197–205. [Google Scholar] [CrossRef]
- Bertoglio, J.C.; Calvo, M.A.; Hancke, J.L.; Burgos, R.A.; Riva, A.; Morazzoni, P.; Ponzone, C.; Magni, C.; Duranti, M. Hypoglycemic effect of lupin seed γ-conglutin in experimental animals and healthy human subjects. Fitoterapia 2011, 82, 933–938. [Google Scholar] [CrossRef]
- Dove, E.R.; Mori, T.A.; Chew, G.T.; Barden, A.E.; Woodman, R.J.; Puddey, I.B.; Sipsas, S.; Hodgson, J.M. Lupin and soya reduce glycaemia acutely in type 2 diabetes. Br. J. Nutr. 2011, 106, 1045–1051. [Google Scholar] [CrossRef]
- Yu, Y. The anti-cancer activity and potential clinical application of rice bran extracts and fermentation products. RSC Adv. 2019, 9, 18060–18069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, M.; Watanabe, R.; Kabasawa, H.; Iino, N.; Saito, A.; Kumagai, T.; Fujimura, S.; Kadowaki, M. Rice protein ameliorates the progression of diabetic nephropathy in Goto–Kakizaki rats with high-sucrose feeding. Br. J. Nutr. 2013, 110, 1211–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, Y.; Hira, T.; Inoue, D.; Harada, Y.; Hashimoto, H.; Fujii, M.; Kadowaki, M.; Hara, H. Rice protein hydrolysates stimulate GLP-1 secretion, reduce GLP-1 degradation, and lower the glycemic response in rats. Food Funct. 2015, 6, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xia, J.; Yu, S.; Yan, J.; He, F.; Zhang, M.; Fan, Q.; Yang, R.; Zhao, W. Natural edible materials made of protein-functionalized aerogel particles for postprandial hyperglycemia management. Int. J. Biol. Macromol. 2021, 167, 279–288. [Google Scholar] [CrossRef]
- Esfandi, R.; Seidu, I.; Willmore, W.; Tsopmo, A. Antioxidant, pancreatic lipase, and α-amylase inhibitory properties of oat bran hydrolyzed proteins and peptides. J. Food Biochem. 2021, 46, e13762. [Google Scholar] [CrossRef]
- Wang, J.B.; Liu, X.R.; Liu, S.Q.; Mao, R.X.; Hou, C.; Zhu, N.; Liu, R.; Ma, H.J.; Li, Y. Hypoglycemic Effects of Oat Oligopeptides in High-Calorie Diet/STZ-Induced Diabetic Rats. Molecules 2019, 24, 558. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.Y.; Siow, P.C.; Peh, E.; Henry, C.J. Influence of rice, pea and oat proteins in attenuating glycemic response of sugar-sweetened beverages. Eur. J. Nutr. 2018, 57, 2795–2803. [Google Scholar] [CrossRef]
- Public Health Agency of Canada. Heart Disease in Canada. Available online: https://www.canada.ca/en/public-health/services/publications/diseases-conditions/heart-disease-canada.html (accessed on 25 August 2020).
- Whelton Paul, K.; Carey Robert, M.; Aronow Wilbert, S.; Casey Donald, E.; Collins Karen, J.; Dennison Himmelfarb, C.; DePalma Sondra, M.; Gidding, S.; Jamerson Kenneth, A.; Jones Daniel, W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults. J. Am. Coll. Cardiol. 2018, 71, e127–e248. [Google Scholar] [CrossRef]
- Messerli, F.H.; Williams, B.; Ritz, E. Essential hypertension. Lancet 2007, 370, 591–603. [Google Scholar] [CrossRef]
- Hansson, G.K.; Hermansson, A. The immune system in atherosclerosis. Nat. Immunol. 2011, 12, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Vermeirssen, V.; Augustijns, P.; Morel, N.; Van Camp, J.; Opsomer, A.; Verstraete, W. In vitro intestinal transport and antihypertensive activity of ACE inhibitory pea and whey digests. Int. J. Food Sci. Nutr. 2005, 56, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Pedroche, J.; Yust, M.M.; Girón-Calle, J.; Alaiz, M.; Millán, F.; Vioque, J. Utilisation of chickpea protein isolates for production of peptides with angiotensin I-converting enzyme (ACE)-inhibitory activity. J. Sci. Food Agric. 2002, 82, 960–965. [Google Scholar] [CrossRef]
- Aluko, R. Determination of Nutritional and Bioactive Properties of Peptides in Enzymatic Pea, Chickpea, and Mung Bean Protein Hydrolysates. J. AOAC Int. 2008, 91, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Alu’datt, M.H.; Ereifej, K.; Abu-Zaiton, A.; Alrababah, M.; Almajwal, A.; Rababah, T.; Yang, W. Anti-Oxidant, Anti-Diabetic, and Anti-Hypertensive Effects of Extracted Phenolics and Hydrolyzed Peptides from Barley Protein Fractions. Int. J. Food Prop. 2012, 15, 781–795. [Google Scholar] [CrossRef]
- Roy, F.; Boye, J.I.; Simpson, B.K. Bioactive proteins and peptides in pulse crops: Pea, chickpea and lentil. Food Res. Int. 2010, 43, 432–442. [Google Scholar] [CrossRef]
- Gomes, M.; Lima, S.; Alves, A.; Moreira, M.; Toledo, R.; Rosa, C.; Teixeira, O.; Bassinello, P.; De Mejia, E.; Martino, H. Common bean protein hydrolysate modulates lipid metabolism and prevents endothelial dysfunction in BALB/c mice fed an atherogenic diet. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Porres, J.M.; Constantino, J.; Kapravelou, G.; Lopez-Chaves, C.; Galisteo, M.; Aranda, P.; López-Jurado, M.; Martínez, R. The combined treatment with lentil protein hydrolysate and a mixed training protocol is an efficient lifestyle intervention to manage cardiovascular and renal alterations in obese Zucker rats. Eur. J. Nutr. 2020, 59, 3473–3490. [Google Scholar] [CrossRef]
- Belski, R.; Mori, T.A.; Puddey, I.B.; Sipsas, S.; Woodman, R.J.; Ackland, T.R.; Beilin, L.J.; Dove, E.R.; Carlyon, N.B.; Jayaseena, V.; et al. Effects of lupin-enriched foods on body composition and cardiovascular disease risk factors: A 12-month randomized controlled weight loss trial. Int. J. Obes. 2011, 35, 810–819. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Duan, W.; Ren, X.; Wang, C.; Pan, Z.; Diao, X.; Shen, Q. Effect of foxtail millet protein hydrolysates on lowering blood pressure in spontaneously hypertensive rats. Eur. J. Nutr. 2017, 56, 2129–2138. [Google Scholar] [CrossRef]
- Yang, T.; Zhu, H.; Zhou, H.; Lin, Q.; Li, W.; Liu, J. Rice protein hydrolysate attenuates hydrogen peroxide induced apoptosis of myocardiocytes H9c2 through the Bcl-2/Bax pathway. Food Res. Int. 2012, 48, 736–741. [Google Scholar] [CrossRef]
- Ni, W.; Tsuda, Y.; Takashima, S.; Sato, H.; Sato, M.; Imaizumi, K. Anti-atherogenic effect of soya and rice-protein isolate, compared with casein, in apolipoprotein E-deficient mice. Br. J. Nutr. 2003, 90, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burris, R.L.; Xie, C.H.; Thampi, P.; Wu, X.; Melnyk, S.B.; Nagarajan, S. Dietary rice protein isolate attenuates atherosclerosis in apoE-deficient mice by upregulating antioxidant enzymes. Atherosclerosis 2010, 212, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Shobako, N.; Fukuhara, I.; Satoh, H.; Kobayashi, E.; Kusakari, T.; Suwa, M.; Matsumoto, M.; Ishikado, A. Rice Bran Supplement Containing A Functional Substance, the Novel Peptide Leu-Arg-Ala, has Anti-Hypertensive Effects: A Double-Blind, Randomized, Placebo-Controlled Study. Nutrients 2019, 11, 726. [Google Scholar] [CrossRef] [Green Version]
- Alauddin, M.; Shirakawa, H.; Koseki, T.; Kijima, N.; Ardiansyah; Budijanto, S.; Islam, J.; Goto, T.; Komai, M. Fermented rice bran supplementation mitigates metabolic syndrome in stroke-prone spontaneously hypertensive rats. BMC Complementary Altern. Med. 2016, 16, 442. [Google Scholar] [CrossRef] [Green Version]
- National Institutes of Health (US); Biological Sciences Curriculum Study. NIH Curriculum Supplement Series [Internet]; Understanding Cancer; National Institutes of Health (US): Bethesda, MD, USA, 2007. Available online: https://www.ncbi.nlm.nih.gov/books/NBK20362/ (accessed on 17 May 2022).
- Ganesh, K.; Massagué, J. Targeting metastatic cancer. Nat. Med. 2021, 27, 34–44. [Google Scholar] [CrossRef]
- Canadian Cancer Society. Cancer Statistics at a Glance. Available online: https://www.cancer.ca/sk/cancer-information/cancer-101/cancer-statistics-at-a-glance/?region=nu (accessed on 18 August 2021).
- Molina-Montes, E.; Salamanca-Fernández, E.; Garcia-Villanova, B.; Sánchez, M.J. The Impact of Plant-Based Dietary Patterns on Cancer-Related Outcomes: A Rapid Review and Meta-Analysis. Nutrients 2020, 12, 2010. [Google Scholar] [CrossRef]
- Hellinger, R.; Gruber, C.W. Peptide-based protease inhibitors from plants. Drug Discov. Today 2019, 24, 1877–1889. [Google Scholar] [CrossRef]
- Clemente, A.; Jimenez, E.; Marin-Manzano, M.C.; Rubio, L.A. Active Bowman–Birk inhibitors survive gastrointestinal digestion at the terminal ileum of pigs fed chickpea-based diets. J. Sci. Food Agric. 2008, 88, 513–521. [Google Scholar] [CrossRef]
- Gitlin-Domagalska, A.; Maciejewska, A.; Dębowski, D.J.P. Bowman-Birk Inhibitors: Insights into Family of Multifunctional Proteins and Peptides with Potential Therapeutical Applications. Pharmaceuticals 2020, 13, 421. [Google Scholar] [CrossRef]
- Ivanova, E.V.; Kondakova, I.V.; Spirina, L.V.; Afanas’ev, S.G.; Avgustinovich, A.V.; Cheremisina, O.V. Chymotrypsin-Like Activity of Proteasomes and Total Calpain Activity in Gastric and Colorectal Cancer. Bull. Exp. Biol. Med. 2014, 157, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, S.; Chen, Z. Plant Protease Inhibitors in Therapeutics-Focus on Cancer Therapy. Front. Pharmacol. 2016, 7, 470. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N. Biochemical characterisation of lectin from wild chickpea (Cicer reticulatum L.) with potential inhibitory action against human cancer cells. J. Food Biochem. 2019, 43, e12712. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Bhagyawant, S.S. Enzymatic treatment improves ACE-I inhibiton and antiproliferative potential of chickpea. Vegetos 2019, 32, 363–369. [Google Scholar] [CrossRef]
- Gupta, N.; Bhagyawant, S.S. Bioactive peptide of Cicer arietinum L. induces apoptosis in human endometrial cancer via DNA fragmentation and cell cycle arrest. 3 Biotech 2021, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Mukhamedov, N.; Wubulikasimu, A.; Rustamova, N.; Nuerxiati, R.; Mirzaakhmedov, S.; Ishimov, U.; Ziyavitdinov, J.; Yili, A.; Aisa, H.A. Synthesis and Characterization of Novel Chickpea Protein Hydrolysate-Vanadium Complexes Having Cell Inhibitory Effects on Lung Cancer A549 Cells Lines. Protein J. 2021, 40, 721–730. [Google Scholar] [CrossRef]
- Sánchez-Chino, X.M. Protective Effect of Chickpea Protein Hydrolysates on Colon Carcinogenesis Associated With a Hypercaloric Diet. J. Am. Coll. Nutr. 2019, 38, 162–170. [Google Scholar] [CrossRef]
- Debatin, K.M. Apoptosis pathways in cancer and cancer therapy. Cancer Immunol. Immunother. 2004, 53, 153–159. [Google Scholar] [CrossRef]
- Luna Vital, D.A.; Loarca-Piña, G.; Dia, V.P.; de Mejía, E.G. Peptides extracted from common bean (Phaseolus vulgaris L.) non-digestible fraction caused differential gene expression of HCT116 and RKO human colorectal cancer cells. Food Res. Int. 2014, 62, 193–204. [Google Scholar] [CrossRef]
- Kannan, A.; Hettiarachchy, N.; Narayan, S. Colon and Breast Anti-cancer Effects of Peptide Hydrolysates Derived from Rice Bran. The Open Bioact. Compd. J. 2009, 2, 17–20. [Google Scholar] [CrossRef] [Green Version]
- Gasymov, O.K.; Celik, S.; Agaeva, G.; Akyuz, S.; Kecel-Gunduz, S.; Qocayev, N.M.; Ozel, A.E.; Agaeva, U.; Bakhishova, M.; Aliyev, J.A. Evaluation of anti-cancer and anti-COVID-19 properties of cationic pentapeptide Glu-Gln-Arg-Pro-Arg, from rice bran protein and its d-isomer analogs through molecular docking simulations. J. Mol. Graph. Model. 2021, 108, 107999. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Fang, N.; Li, Q.; Zhang, J.; Luo, H.; Ronis, M.; Badger, T.M. In vitro actions on human cancer cells and the liquid chromatography-mass spectrometry/mass spectrometry fingerprint of phytochemicals in rice protein isolate. J. Agric. Food Chem. 2006, 54, 4482–4492. [Google Scholar] [CrossRef]
- Liao, H.-F.; Chen, Y.-J.; Chen, Y.-Y. Abstract 3806: Stimulation of anti-leukemia immune response by a rice protein prolamin. Cancer Res. 2010, 70 (Suppl. 8), 3806. [Google Scholar]
- Morita, T.; Kiriyama, S. A rice protein isolate alters 7,12-dimethylbenz[alpha]anthracene-induced mammary tumor development in female rats. J. Nutr. Sci. Vitaminol. 1996, 42, 325–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, H.J.; Jeong, J.B.; Kim, D.S.; Park, J.H.; Lee, J.B.; Kweon, D.-H.; Chung, G.Y.; Seo, E.W.; de Lumen, B.O. The cancer preventive peptide lunasin from wheat inhibits core histone acetylation. Cancer Lett. 2007, 255, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Lam, Y.; de Lumen, B.O. Barley lunasin suppresses ras-induced colony formation and inhibits core histone acetylation in mammalian cells. J. Agric. Food Chem. 2002, 50, 5903–5908. [Google Scholar] [CrossRef]
- Jeong, H.J.; Lee, J.R.; Jeong, J.B.; Park, J.H.; Cheong, Y.-k.; de Lumen, B.O. The Cancer Preventive Seed Peptide Lunasin From Rye Is Bioavailable and Bioactive. Nutr. Cancer 2009, 61, 680–686. [Google Scholar] [CrossRef]
- Nakurte, I.; Kirhnere, I.; Namniece, J.; Saleniece, K.; Krigere, L.; Mekss, P.; Vicupe, Z.; Bleidere, M.; Legzdina, L.; Muceniece, R. Detection of the lunasin peptide in oats (Avena sativa L). J. Cereal Sci. 2013, 57, 319–324. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Martínez-Villaluenga, C.; de Lumen, B.O.; Hernández-Ledesma, B. Updating the research on the chemopreventive and therapeutic role of the peptide lunasin. J. Sci. Food Agric. 2018, 98, 2070–2079. [Google Scholar] [CrossRef] [Green Version]
- McConnell, E.J.; Devapatla, B.; Yaddanapudi, K.; Davis, K.R. The soybean-derived peptide lunasin inhibits non-small cell lung cancer cell proliferation by suppressing phosphorylation of the retinoblastoma protein. Oncotarget 2015, 6, 4649–4662. [Google Scholar] [CrossRef] [Green Version]
- Dia, V.P.; Gonzalez de Mejia, E. Lunasin induces apoptosis and modifies the expression of genes associated with extracellular matrix and cell adhesion in human metastatic colon cancer cells. Mol. Nutr. Food Res. 2011, 55, 623–634. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Activity | Type of Protein | Bioactive Peptide Sequence | IC50 | Description | Reference |
|---|---|---|---|---|---|
| Antihypertensive | Naked oat globulin | SSYYPFK | 91.82 µM | ACE inhibitor; alcalase, flavourzyme, pepsin and trypsin digestion | [36] |
| Antihypertensive | Oat 11S and 12S globulin | GQ, QC, GL, PQ, AG | 30–50 µg/mL | ACE inhibitor; thermolysin digestion | [37] |
| Antihypertensive | Barley protein concentrate | FQLPKF, GFPTLKIF, ALRYFM, NFLARF | 28.2, 41.2, 200, 100 µM | ACE inhibitor; papain digestion | [38] |
| Antihypertensive | Corn gluten hydrolysate | AY | 0.037 mg/mL | ACE inhibitor; antihypertensive effect in rats | [39] |
| Antihypertensive | Corn gluten hydrolysate | PSGQYY | 100 µM | ACE inhibitor; antihypertensive effect in rats; pescalase digestion | [40] |
| Antihypertensive; anticancer | Wheat germ protein hydrolysate | SGGSYADELVSTAK, MDATALHYENQK | 0.09, 0.21 µM | ACE inhibitor; inhibited A549 lung cancer cell growth; proteinase K digestion | [41] |
| Antihypertensive | Wheat gliadin hydrolysate | IAP | 2.7 µM | ACE inhibitor; acid protease digestion | [42] |
| Antihypertensive; antioxidant | Rice protein hydrolysate | VNP, VWP | 6.4, 4.5 µM | ACE inhibitor; alcalase and trypsin digestion | [43] |
| Antihypertensive; antioxidant | Rice bran protein hydrolysate | YSK | 76 µM | ACE inhibitor; high DPPH radical scavenging activity; trypsin digestion | [44] |
| Antihypertensive | Rice bran protein hydrolysate | GSGYF | 3.98 µM | ACE inhibitor; pepsin and trypsin digestion | [45] |
| Antihypertensive | Rice bran vicilin-like protein | LRA, YY | 0.25, 0.5 mg/kg | ACE inhibitor; antihypertensive effect in SHRs; thermolysin digestion | [46] |
| Anticancer | Barley lunasin peptide | SKWQHQQDSCRKQKQGVNLTP CEKHIMEKIQGRGDDDDDDDDD | – | Inhibits histone acetyltransferase and Rb hyperphosphorylation; increases expression of tumour suppressors | [47] |
| Antihyperglycemic | Wheat gluten | ILDL, ILLPGAQDGL | 1121.1, 145.5 µM | DPP-IV inhibitor; alcalase digestion | [48] |
| Antihyperglycemic | Oat globulin | GDVVALPA, DVVALPAG | – | DPP-IV inhibitor; alcalase and flavourzyme digestion | [49] |
| Antihyperglycemic | Oat globulin | LQAFEPLR | 103.5 µM | DPP-IV inhibitor; alcalase digestion | [50] |
| Anti-obesity; anti-Alzheimer’s; anticancer | Rice bran protein hydrolysate | EQRPR | – | Insulin-like differentiation of preadipocytes; reduction in cytotoxicity of amyloid-induced neuroblastoma cells; antiproliferative effects on colon, breast, lung and liver cancer cell lines; alcalase digestion | [51,52] |
| Antioxidant | Finger millet protein | TSSSLNMAVRGGLTR and STTVGLGISMRSASVR | – | DPPH radical scavenging activity; trypsin digestion | [53] |
| Antioxidant | Sorghum kafirin | YLRQ, AQVAQ, AMCGVV | – | DPPH radical scavenging activity; papain digestion | [12] |
| Antioxidant; antihypertensive | Corn prolamin | MI/LPP | 220 µg/mL (antioxidant), 70.32 µg/mL (ACE) | DPPH radical scavenging activity; ACE inhibitor; alcalase digestion | [54] |
| Activity | Type of Protein | Bioactive Peptide Sequence | IC50 | Description | Reference |
|---|---|---|---|---|---|
| Antihypertensive | Chickpea legumin | MDLA, MDFLI, MFDL | 0.01–0.02 mg/mL | ACE inhibitor; alcalase digestion | [55] |
| Antihypertensive | Mungbean protein isolate | KDYRL, VTPARLR, KLPAGTLF | 26.5, 82.4, 13.4 µM | ACE inhibitor; alcalase digestion | [56] |
| Antihypertensive | Lentil globulin | KLRT, TLHGMV, VNRLM | 0.13 mg/mL | ACE inhibitor; α-amylase, pepsin and pancreatin digestion | [57] |
| Antihypertensive | Pea globulin | GGSGNY, DLKLP, GSSDNR, MRDLK, HNTPSR | 0.07 mg/mL | ACE inhibitor; α-amylase, pepsin and pancreatin digestion | [58] |
| Antihypertensive; antihyperglycemic | Common bean protein hydrolysate | KTYGL, KKSSG | 0.09 and 0.20 mg DW/mL (ACE), 0.03 and 0.64 mg DW/mL (DPP-IV) | ACE and DPP-IV inhibitor; pepsin and pancreatin digestion | [59] |
| Anticancer | Common bean protein hydrolysate | GLTSK, LSGNK, GEGSGA, MPACGSS, MTEEY | - | Antiproliferative effects in HCT116 and RKO cell lines; sequential enzyme digestion | [60] |
| Anti-inflammatory | Common bean protein hydrolysate | γ-EV | - | Anti-inflammatory activity in intestinal epithelial Caco-2 cells; synthetic peptide | [61] |
| Antiglycemic; antihypertensive; antioxidant | Lentil protein | SDQENPFIFK, HGDPEER, ATAFGLMK | 0.39 mg/mL (ACE) | ACE, α-glycosidase and maltase inhibitor; antioxidant activities; savinase digestion | [62] |
| Antimicrobial | Lentil defensin peptide | KTCENLSDSFKGPCIPDGNCNKHCKEKEHLLSGRCRDDFRCWCTRNC | - | Inhibits growth of Aspergillus niger | [63] |
| Cholesterol-lowering; anti-inflammatory | Chickpea protein hydrolysate | RQSHFANAQP (CPe-III) | - | Antihyperlipidemic and anti-inflammatory effects in Kunming mice; synthetic peptide | [64] |
| Cholesterol-lowering | Chickpea protein hydrolysate | VFVRN | - | HMGR inhibitor, decreases TC synthesis; alcalase digestion | [65] |
| Cholesterol-lowering | Lupin protein | LILPKHSDAD, LTFPGSAED | 147.2, 68.4 µM | HMGR inhibitor, synthetic peptides | [66] |
| Cholesterol-lowering | Lupin protein | LILPHKSDAD | 1.6 μM | PCSK9 inhibitor, HMGR inhibitor, synthetic peptide | [67] |
| Item | DIAAS Score (%) | Reference |
|---|---|---|
| Oats | 43–57 (Lys) | [84,85,86] |
| Dehulled oats | 77 (Lys) | [87] |
| Wheat | 43–48 (Lys) | [80,81,87] |
| Dehulled barley | 51–77 (Lys) | [81,87] |
| Rye | 47–56 (Lys) | [81,87,88] |
| Rice protein concentrate | 37 (Lys) | [86,89] |
| Brown rice (cooked) | 42 (Lys) | [79] |
| Polished white rice | 37–64 (Lys) | [79,81,87] |
| Foxtail millet (cooked) | 10–22 (Lys) | [78,79] |
| Corn | 36–48 (Lys) | [79,87] |
| Sorghum | 29–45 (Lys) | [81,90] |
| Split green peas (cooked) | 46 (SAAs) | [91] |
| Split yellow peas (cooked) | 73 (SAAs) | [84] |
| Chickpeas | 83–89 (SAAs) | [82,83] |
| Peas | 58–70 (SAAs) | [80,82,86] |
| Pea protein concentrate | 62–82 (SAAs) | [82,83,86] |
| Pigeon peas | 57 (SAAs) | [83] |
| Fava beans | 55 (SAAs) | [81] |
| Kidney beans (cooked) | 51–58 (SAAs) | [84,86] |
| Black beans (cooked) | 43–49 (SAAs) | [83,84] |
| Pinto beans (cooked) | 60–83 (SAAs) | [84,92] |
| Navy beans (cooked) | 65 (SAAs) | [84] |
| Mung beans (cooked) | 93 (Val) | [78] |
| Whole green lentils (cooked) | 49–58 (SAA) | [84,91] |
| Split red lentils (cooked) | 50–54 (SAA) | [84,91] |
| Lupins | 68 (SAA) | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouchard, J.; Malalgoda, M.; Storsley, J.; Malunga, L.; Netticadan, T.; Thandapilly, S.J. Health Benefits of Cereal Grain- and Pulse-Derived Proteins. Molecules 2022, 27, 3746. https://doi.org/10.3390/molecules27123746
Bouchard J, Malalgoda M, Storsley J, Malunga L, Netticadan T, Thandapilly SJ. Health Benefits of Cereal Grain- and Pulse-Derived Proteins. Molecules. 2022; 27(12):3746. https://doi.org/10.3390/molecules27123746
Chicago/Turabian StyleBouchard, Jenny, Maneka Malalgoda, Joanne Storsley, Lovemore Malunga, Thomas Netticadan, and Sijo Joseph Thandapilly. 2022. "Health Benefits of Cereal Grain- and Pulse-Derived Proteins" Molecules 27, no. 12: 3746. https://doi.org/10.3390/molecules27123746
APA StyleBouchard, J., Malalgoda, M., Storsley, J., Malunga, L., Netticadan, T., & Thandapilly, S. J. (2022). Health Benefits of Cereal Grain- and Pulse-Derived Proteins. Molecules, 27(12), 3746. https://doi.org/10.3390/molecules27123746