
The Regulating Effect of CII-3 and Its Active Components from Periplaneta americana on M1/M2 Macrophage Polarization

 , , and
, , and
Abstract
:1. Introduction
2. Results
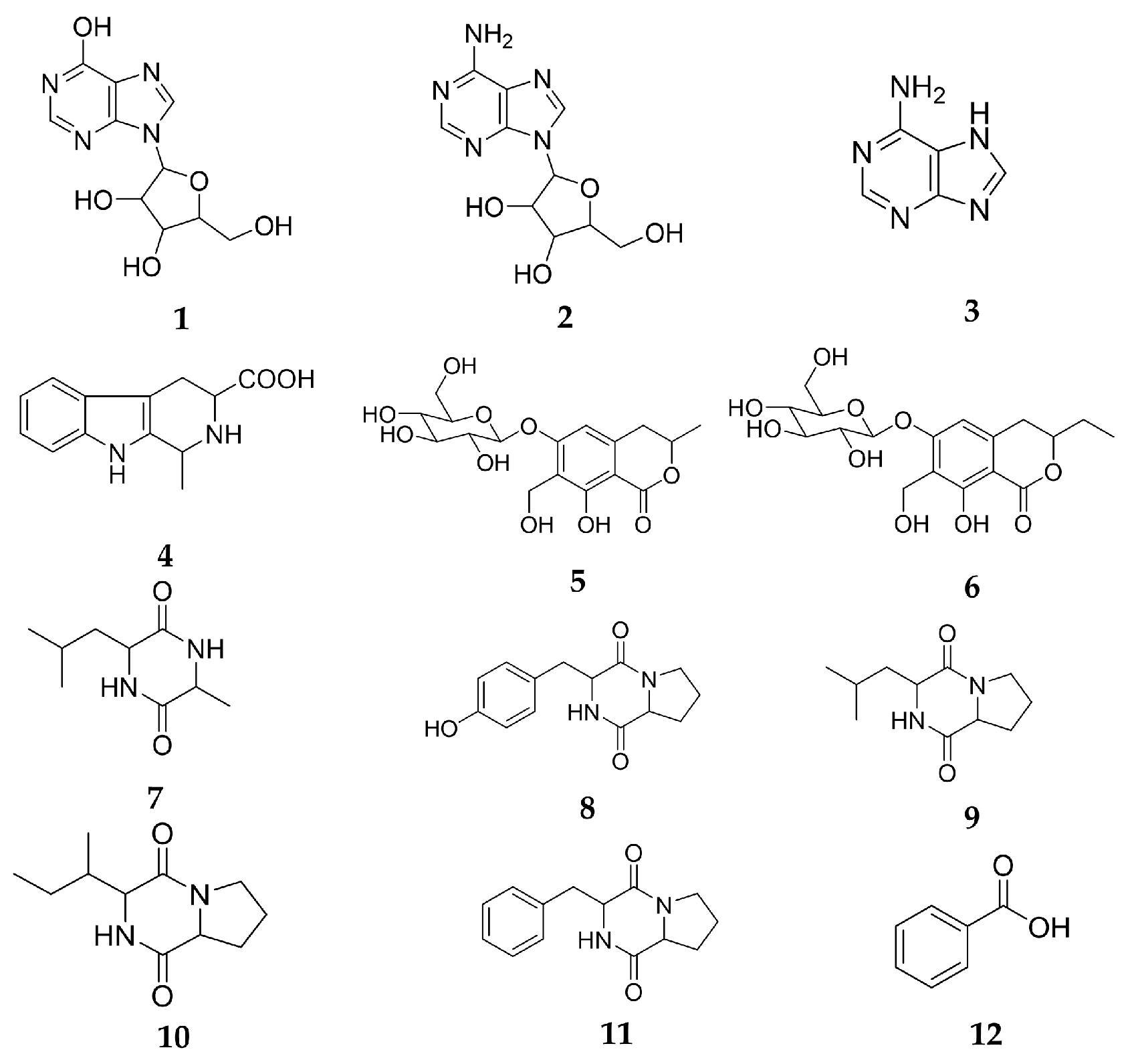
2.1. Main Chemical Components of CII-3
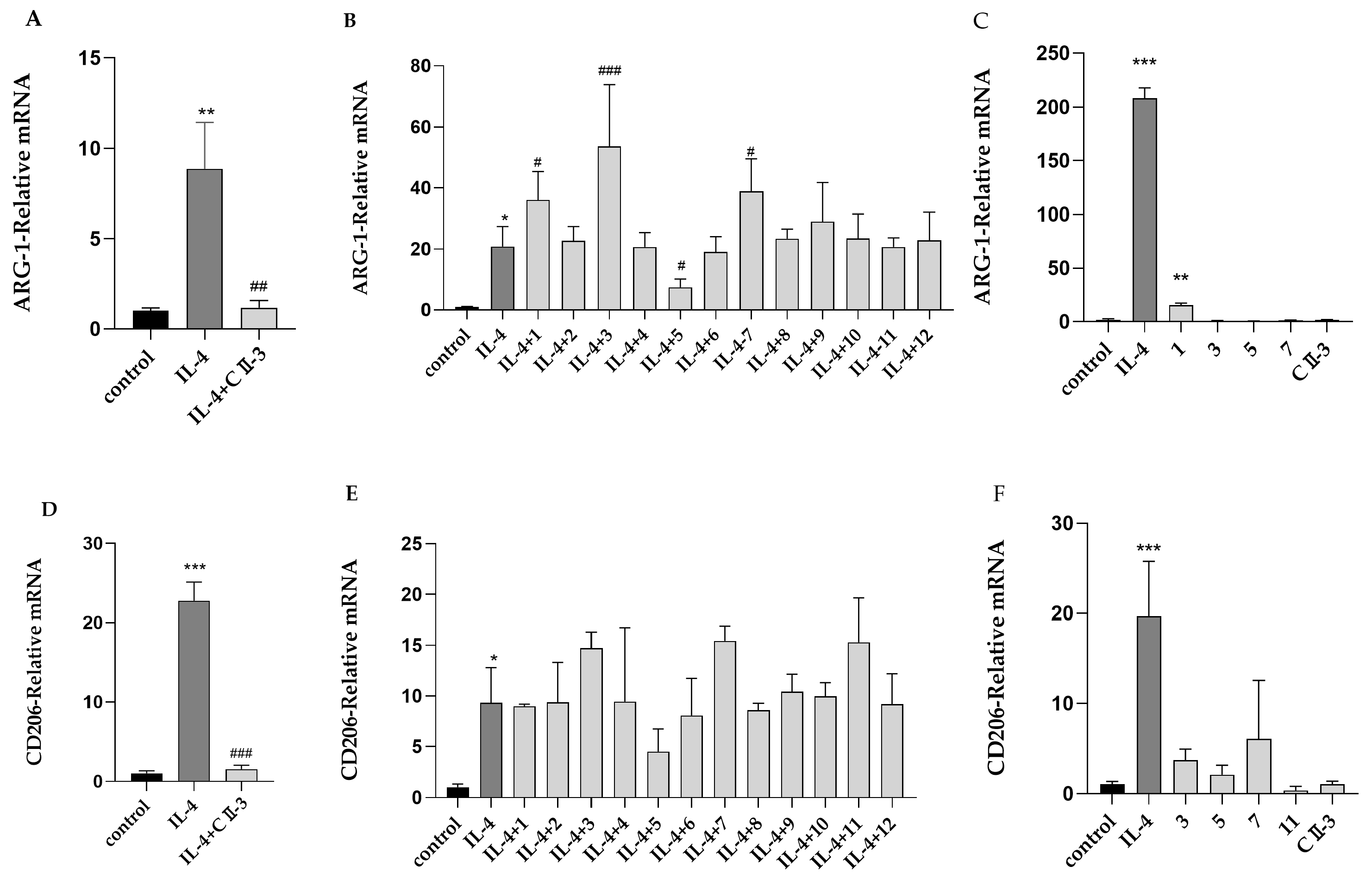
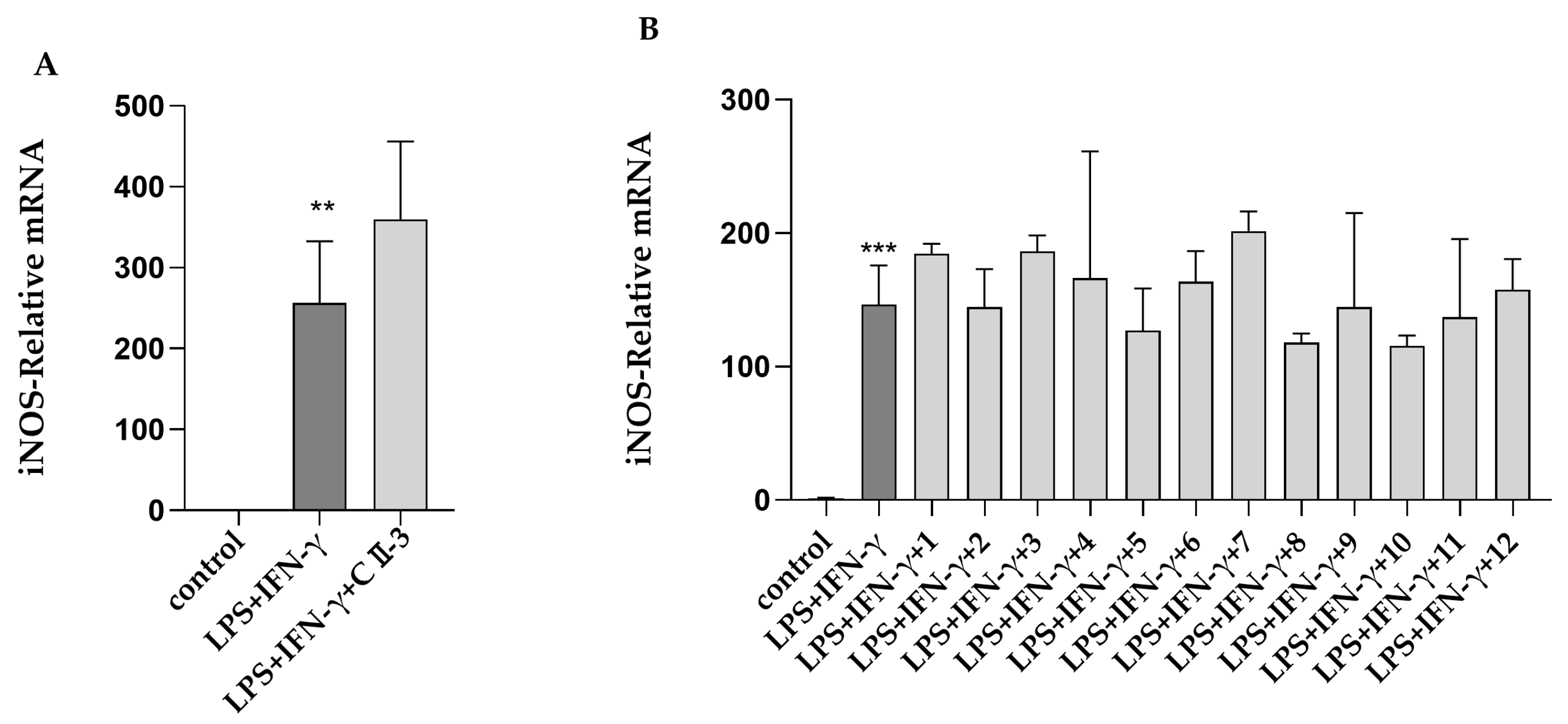
2.2. Primary Activity Screening of CII-3 and 12 Compounds on M1/M2 Macrophage Polarization
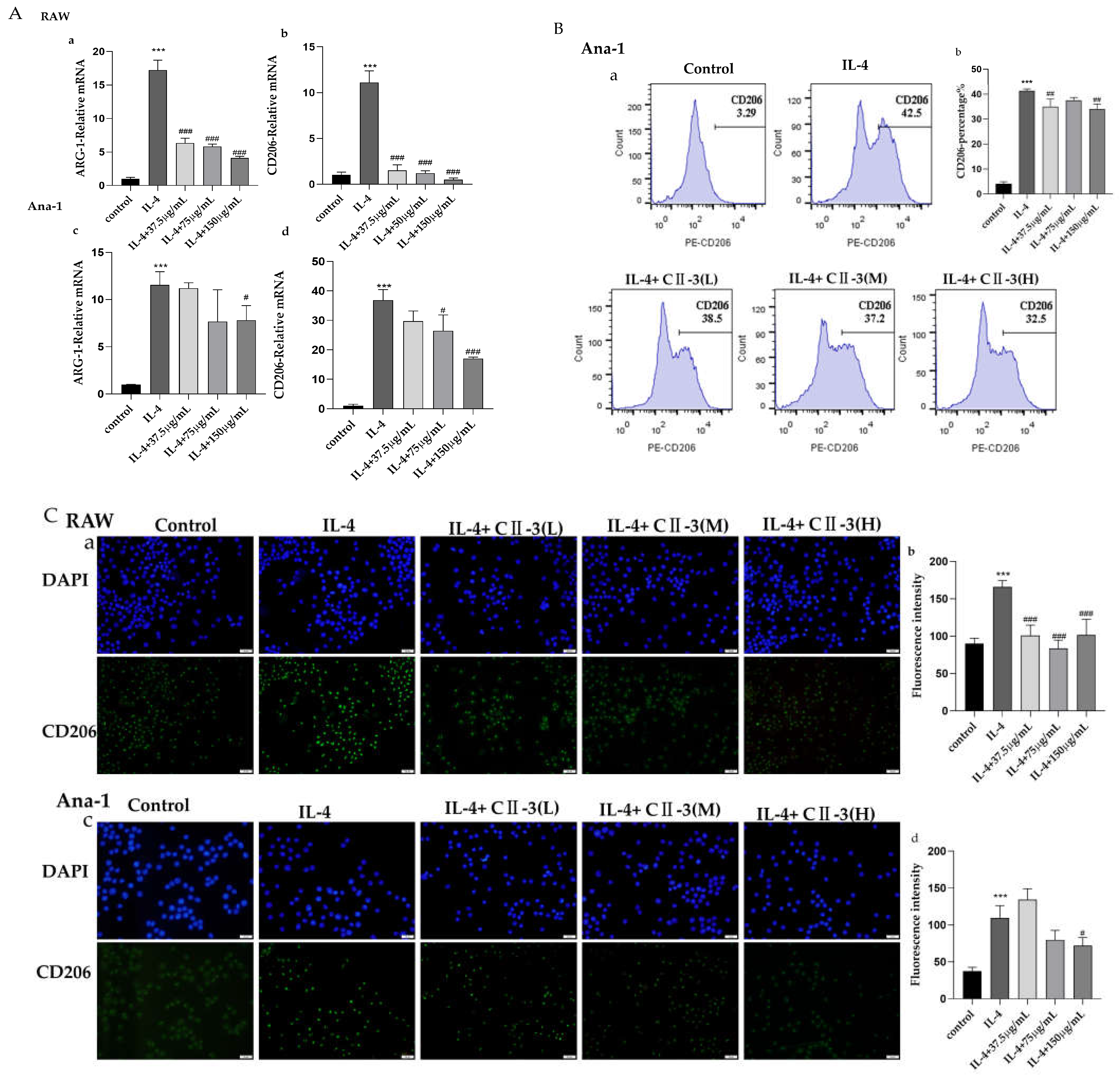
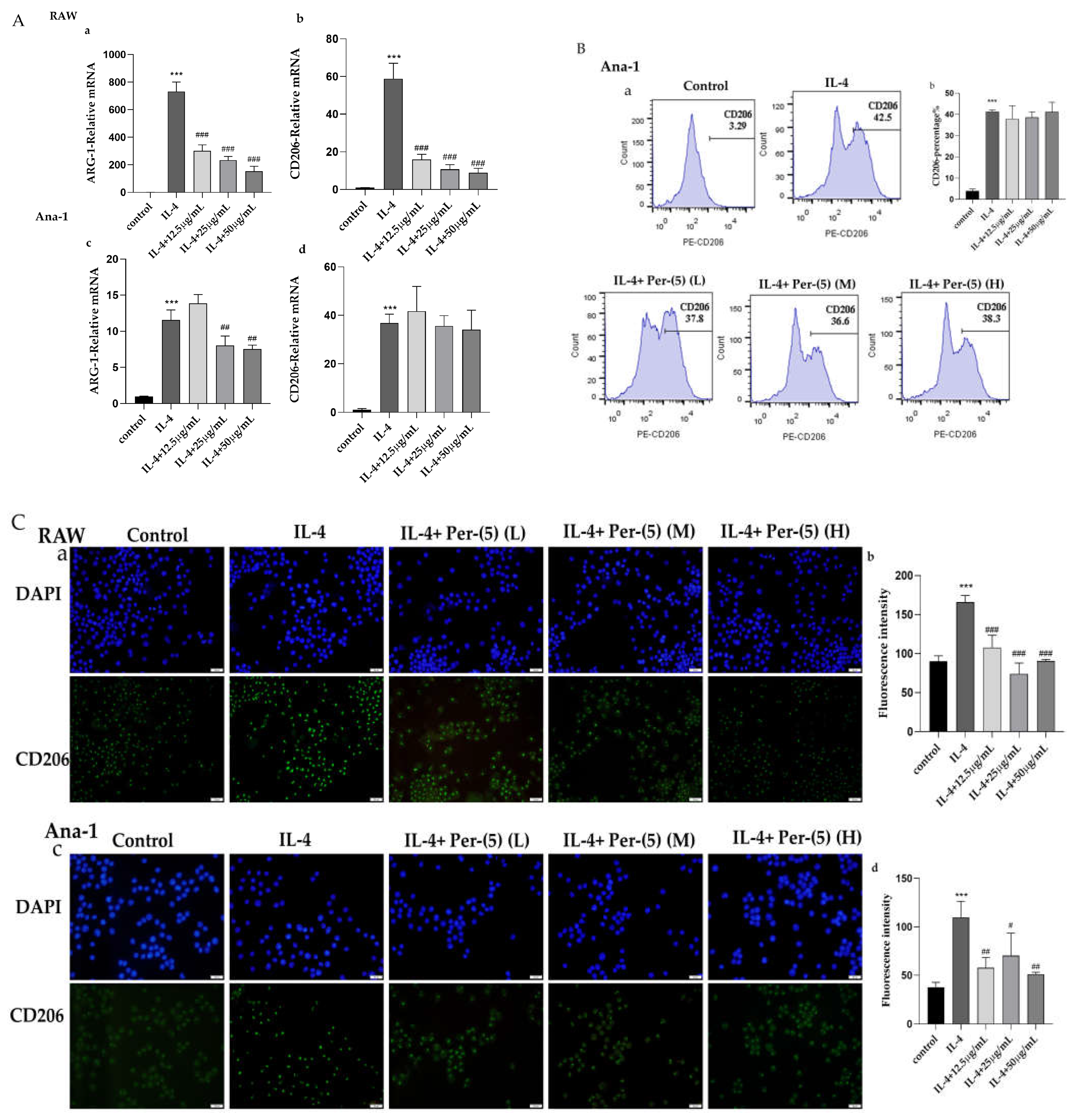
2.3. The Influence of CII-3 and Pericanaside (5) on M2 Markers ARG-1 and CD206
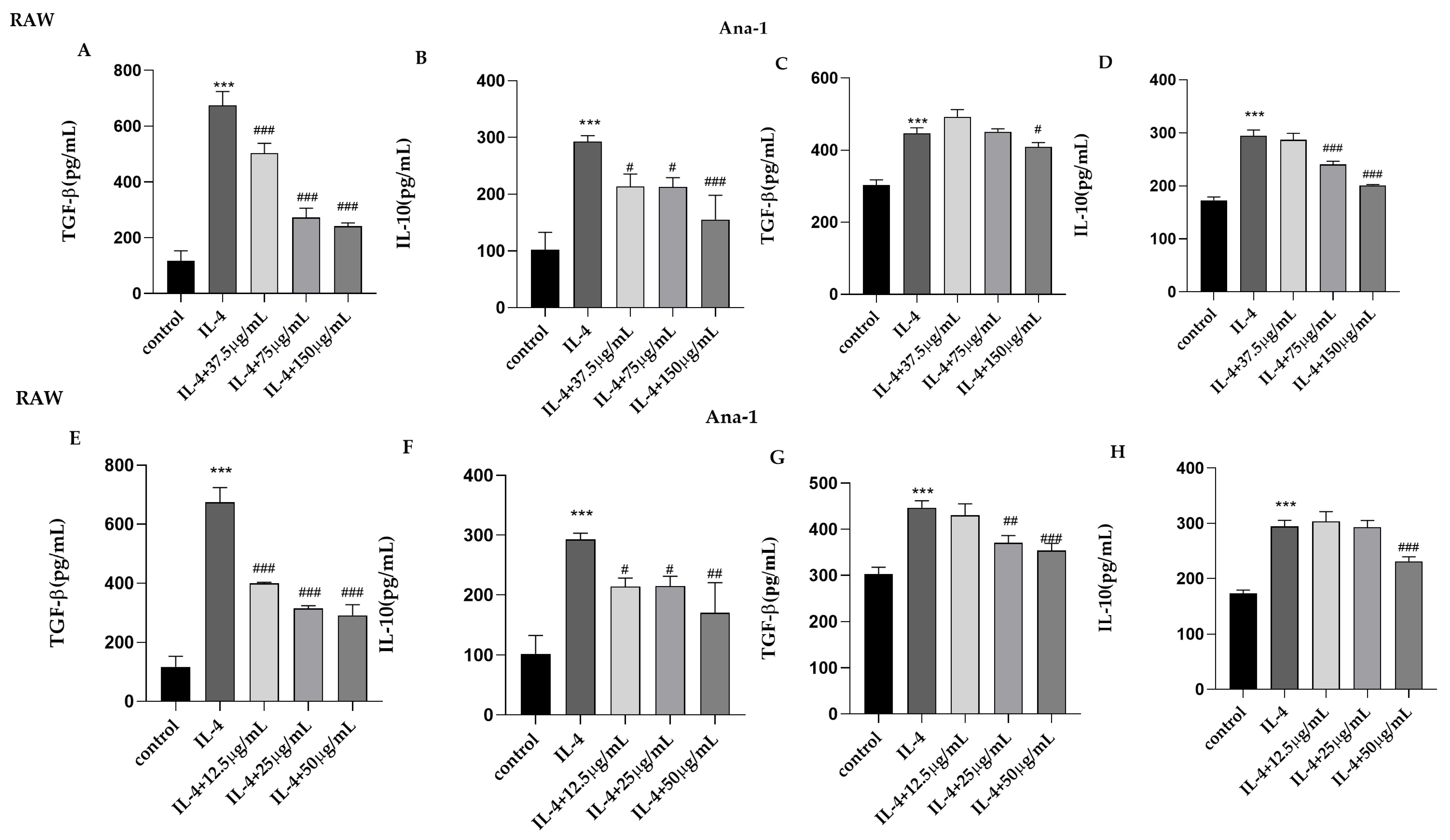
2.4. The Influence of CII-3 and Pericanaside (5) on TGF-β and IL-10
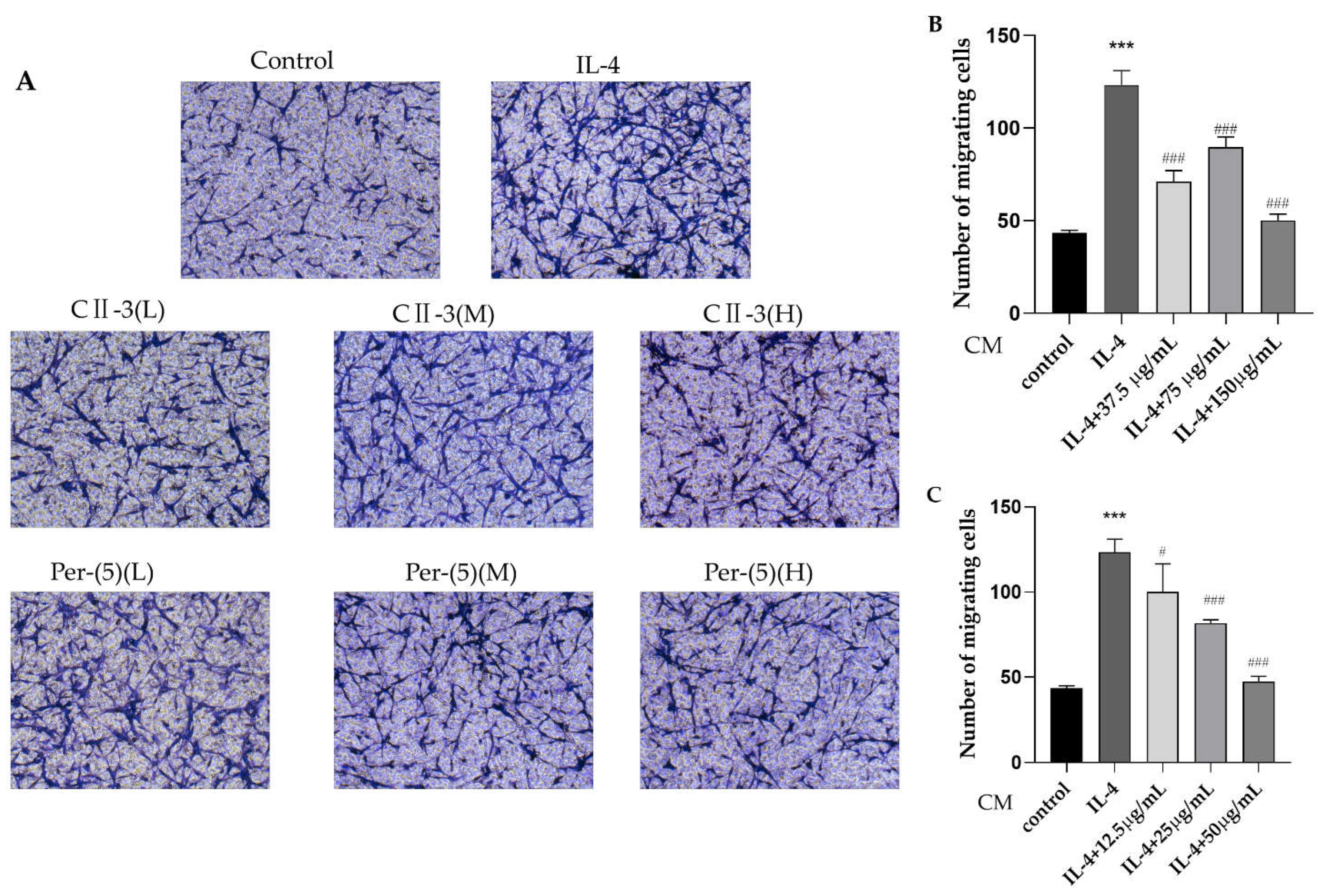
2.5. The Influence of Conditioned Medium on CT26.WT Cell Migration by Transwell Assays
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. Reagents
4.1.2. Device
4.2. Methods
4.2.1. Isolation and Identification
4.2.2. M1/M2 Polarization Models for RAW 264.7 and Ana-1 Cells
4.2.3. Cell Viability Assay
4.2.4. RT-PCR
4.2.5. Enzyme-Linked Immunosorbent Assay
4.2.6. Flow Cytometry
4.2.7. Immunofluorescence
4.2.8. Conditioned Medium
4.2.9. Transwell Migration Assay
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5-3) | GeneBank ID |
|---|---|---|
| iNOS | Forward: TTTGCTTCCATGCTAATGCGAAAG Reverse: GCTCTGTTGAGGTCTAAAGGCTCCG | NM_001313922.1 |
| CD206 | Forward: CATGAGGCTTCTCTTGCTTCTG Reverse: TTGCCGTCTGAACTGAGATGG | XM_021173919.1 |
| ARG-1 | Forward: CTCCAAGCCAAAGTCCTTAGAG Reverse: AGGAGCTGTCATTAGGGACATC | NM_008625.2 |
| GAPDH | Forward: ATACGGCTACAGCAACAGGG Reverse: GCCTCTCTTGCTCAGTGTCC | NM_001289726.1 |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Fidler, M.M.; Bray, F.; Soerjomataram, I. The global cancer burden and human development: A review. Scand. J. Public Health 2017, 46, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De-Brito, N.M.; Duncan-Moretti, J.; Da-Costa, H.C.; Saldanha-Gama, R.; Paula-Neto, H.A.; Dorighello, G.; Simões, R.L.; Barja-Fidalgo, C. Aerobic glycolysis is a metabolic requirement to maintain the M2-like polarization of tumor-associated macrophages. Biochim. Biophys. Acta 2019, 1867, 118604. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-W.; Lee, B.; Liu, P.-S.; Wei, L.-N. Receptor-Interacting Protein 140 Orchestrates the Dynamics of Macrophage M1/M2 Polarization. J. Innate Immun. 2016, 8, 97–107. [Google Scholar] [CrossRef]
- Baek, S.H.; Lee, H.W.; Gangadaran, P.; Oh, J.M.; Zhu, L.; Rajendran, R.L.; Lee, J.; Ahn, B.-C. Role of M2-like macrophages in the progression of ovarian cancer. Exp. Cell Res. 2020, 395, 112211. [Google Scholar] [CrossRef]
- Wu, J.-Y.; Huang, T.-W.; Hsieh, Y.-T.; Wang, Y.-F.; Yen, C.-C.; Lee, G.-L.; Yeh, C.-C.; Peng, Y.-J.; Kuo, Y.-Y.; Wen, H.-T.; et al. Cancer-Derived Succinate Promotes Macrophage Polarization and Cancer Metastasis via Succinate Receptor. Mol. Cell 2020, 77, 213–227.e5. [Google Scholar] [CrossRef]
- Li, L.-J.; Wang, M.-Z.; Yuan, T.-J.; Xu, X.-H.; Dad, H.A.; Yu, C.-L.; Hou, J.; Peng, L.-H. The crude ethanol extract of Periplaneta americana L. stimulates wound healing in vitro & in vivo. Chin. Med. 2019, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lou, Y.; Wang, J.; Yu, C.; Shen, W. Research Status and Molecular Mechanism of the Traditional Chinese Medicine and Antitumor Therapy Combined Strategy Based on Tumor Microenvironment. Front. Immunol. 2021, 11, 609705. [Google Scholar] [CrossRef]
- Xue, N.-N.; He, M.; Li, Y.; Wu, J.-Z.; Du, W.-W.; Wu, X.-M.; Yang, Z.-Z.; Zhang, C.-G.; Li, Q.-Y.; Xiao, H. Periplaneta americana extract promotes intestinal mucosa repair of ulcerative colitis in rat. Acta Cir. Bras. 2020, 35, e202001002. [Google Scholar] [CrossRef]
- Li, Q.-J.; Wang, Z.-G.; Liu, Q.; Xie, Y.; Hu, H.-L. Research status of Periplaneta americana with analyses and prospects of key issues. China J. Chin. Mater. Med. 2018, 43, 1507–1516. [Google Scholar]
- Zhao, Y.; Yang, A.; Tu, P.; Hu, Z. Anti-tumor effects of the American cockroach, Periplaneta americana. Chin. Med. 2017, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-J.; Xu, X.-H.; Yuan, T.-J.; Hou, J.; Yu, C.-L.; Peng, L.-H. Periplaneta Americana L. as a novel therapeutics accelerates wound repair and regeneration. Biomed. Pharmacother. 2019, 114, 108858. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Chen, X.; Chai, J.; Li, R.; Han, X.; Chen, X.; Liu, S.; Chen, M.; Xu, X. Antipyretic, anti-inflammatory and analgesic activities of Periplaneta americana extract and underlying mechanisms. Biomed. Pharmacother. 2019, 123, 109753. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Wang, Y.; Zhang, R.; Zhang, H.H.; Peng, F. Study on Anti-tumor Effects of Periplaneta americana Polypeptide PAP-2 on H22 Tumor-bearing Mice. J. Chin. Pharm. 2019, 30, 5. [Google Scholar] [CrossRef]
- Chang, X.; Wang, C.; Ou, H.; Zhang, W.; Wu, X.M.; Zhang, C.G.; Liu, G.M.; Bai, L. Preliminary study on the effect of Periplaneta americana peptide on immune of MFC tumor bearing mice. Immunol. J. 2017, 33, 6. [Google Scholar] [CrossRef]
- Ou, H.L.; Chang, X.; Wang, X.; Lv, Y.; Peng, F.; Wu, X.M.; Zhang, C.G.; Bai, L. Preliminary Study of Peptide from Periplaneta americana on Antitumor Immunity in L1210-Bearing Mice. Chin. Pharm. J. 2018, 53, 6. [Google Scholar] [CrossRef]
- Zeng, C.; Liao, Q.; Hu, Y.; Shen, Y.; Geng, F.; Chen, L. The Role of Periplaneta americana (Blattodea: Blattidae) in Modern Versus Traditional Chinese Medicine. J. Med. Entomol. 2019, 56, 1522–1526. [Google Scholar] [CrossRef]
- Wu, D.X.; Shao, W.L.; Yang, X.Y.; Chen, J.R.; Liu, G.M.; Chen, J.Y. Progress of studies on the Anti-tumor Activity and Immunomodulating Effcet progress of studies Periplaneta americana. Asia-Pac. Trad. Med. 2016, 12, 48–51. [Google Scholar] [CrossRef]
- Zhao, W. The Function of Periplaneta americana Extract in Anti-Tumor Immune Regulation; Dali University: Dali, China, 2015. [Google Scholar]
- Gao, Y.; Liang, L.C.; Wang, R.; Cheng, L.; Guo, F.J.; Li, Y.M. Chemical constituents from Periplaneta americana. Chin. Tradit. Pat. Med. 2018, 40, 4. [Google Scholar] [CrossRef]
- Yu, Y.; Song, W.X.; Guo, Q.L.; Lin, S.; Wang, S.J.; Yang, Y.C.; Shi, J.G. Studies on chemical constituents of aqueous extract of Lonicera japonica flower buds. China J. Chin. Mater. Medica 2015, 40, 3496–3504. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zeng, G.; Zhang, M.; Liu, Y. Chemical constituents from rhizoma of Anemone altaica. Chin. Tradit. Herbal. Drugs 2014, 45, 1219–1222. [Google Scholar] [CrossRef]
- Li, Y.; Wang, F.; Zhang, P.-Z.; Yang, M. Chemical Constituents from Periplaneta americana. J. Chin. Med. Mater. 2015, 38, 1219–1222. [Google Scholar]
- Yang, Y.-X.; Luo, Q.; Hou, B.; Yan, Y.-M.; Wang, Y.-H.; Tang, J.-J.; Dong, X.-P.; Ma, X.-Y.; Yang, T.-H.; Zuo, Z.-L.; et al. Periplanosides A–C: New insect-derived dihydroisocoumarin glucosides from Periplaneta americana stimulating collagen production in human dermal fibroblasts. J. Asian Nat. Prod. Res. 2015, 17, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.T.; Zhang, P.; LWang, G.; Wang, G.K.; Liu, J.S.; Xu, F.Q.; Liu, H.T.; Wu, P.Y. Secondary metabolites from endophytic fungus Fusarium oxysporum isolated from Paeonia ostii. Chin. Tradit. Herbal. Drugs. 2018, 49, 7. [Google Scholar] [CrossRef]
- Li, L.; Yan, Y.M.; Li, X.N.; Zhong, H.M.; Chen, Y.X. Nitrogenous Compounds from Holotrichia diomphalia Bates. Nat. Prod. Res. Dev. 2013, 25, 4. [Google Scholar] [CrossRef]
- Zhang, T.; Si, J.G.; Chen, J.H.; Zhang, Q.B.; Zou, Z.M. Antioxidant Constituents from Periplaneta americana. Nat. Prod. Res. Dev. 2017, 29, 7. [Google Scholar] [CrossRef]
- Fu, Z.Q.; Huang, Z.H.; Lin, J.; He, W.D.; Ji, M.M.; Xu, W.; Fan, S.M. Chemical constituents in root tuber of Tetrastigma hemsleyanum and their anti-oxidative activities. Chin. Tradit. Herbal Drugs 2015, 46, 1583–1588. [Google Scholar] [CrossRef]
- Shu, Y.; Cheng, P. Targeting tumor-associated macrophages for cancer immunotherapy. Biochim. et Biophys. Acta 2020, 1874, 188434. [Google Scholar] [CrossRef]
- Yuan, C.; Yang, D.; Ma, J.; Yang, J.; Xue, J.; Song, F.; Liu, X. Modulation of Wnt/β-catenin signaling in IL-17A-mediated macrophage polarization of RAW264.7 cells. Braz. J. Med. Biol. Res. 2020, 53, e9488. [Google Scholar] [CrossRef]
- Funes, S.C.; Rios, M.; Escobar-Vera, J.; Kalergis, A.M. Implications of macrophage polarization in autoimmunity. Immunology 2018, 154, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Suarez-Lopez, L.; Sriram, G.; Kong, Y.W.; Morandell, S.; Merrick, K.A.; Hernandez, Y.; Haigis, K.M.; Yaffe, M.B. MK2 contributes to tumor progression by promoting M2 macrophage polarization and tumor angiogenesis. Proc. Natl. Acad. Sci. USA 2018, 115, E4236–E4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.T.; Pan, H.F.; Zhang, H.; Long, R.Y.; Luo, J.M.; Feng, J.H. Research progress of M2 macrophage marker CD163 and tumor. Chin. J. Immunol. 2018, 34, 5. [Google Scholar] [CrossRef]
- Hao, N.-B.; Lü, M.-H.; Fan, Y.-H.; Cao, Y.-L.; Zhang, Z.-R.; Yang, S.-M. Macrophages in Tumor Microenvironments and the Progression of Tumors. Clin. Dev. Immunol. 2012, 2012, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, R.; Shukla, N. Attributes of alternatively activated (M2) macrophages. Life Sci. 2019, 224, 222–231. [Google Scholar] [CrossRef]
- Li, Y.; Gao, L.; Li, H.C.; Yu, H.J.; Zhang, Y. M2 macrophage marker CD206 and tumor. J. Int. Oncol. 2019, 46, 4. [Google Scholar] [CrossRef]
- Xue, N.; Zhou, Q.; Ji, M.; Jin, J.; Lai, F.; Chen, J.; Zhang, M.; Jia, J.; Yang, H.; Zhang, J.; et al. Chlorogenic acid inhibits glioblastoma growth through repolarizating macrophage from M2 to M1 phenotype. Sci. Rep. 2017, 7, 39011. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Fang, Y.; Yuan, W.F.; Li, L.; Huang, W.J. Comparison between two kinds of murine macrophages.Ana-1 and RAW264.7 cell in appearance and cytokines expression. Int. J. Respir. 2009, 5. [Google Scholar]
- Liu, G.M.; He, Z.C.; Wang, X.Y. American cockroaches effective fraction for anti-tumor prepared by macroporous adsorption resin and use. CN101214262B, 5 September 2012. [Google Scholar]
- Sui, H.; Tan, H.; Fu, J.; Song, Q.; Jia, R.; Han, L.; Lv, Y.; Zhang, H.; Zheng, D.; Dong, L.; et al. The active fraction of Garcinia yunnanensis suppresses the progression of colorectal carcinoma by interfering with tumorassociated macrophage-associated M2 macrophage polarization in vivo and in vitro. FASEB J. 2020, 34, 7387–7403. [Google Scholar] [CrossRef]
- Biswas, S.K.; Mantovani, A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef]
- Ye, Y.; Xu, Y.; Lai, Y.; He, W.; Li, Y.; Wang, R.; Luo, X.; Chen, R.; Chen, T. Long non-coding RNA cox-2 prevents immune evasion and metastasis of hepatocellular carcinoma by altering M1/M2 macrophage polarization. J. Cell. Biochem. 2017, 119, 2951–2963. [Google Scholar] [CrossRef]
- Cao, J.; Dong, R.; Jiang, L.; Gong, Y.; Yuan, M.; You, J.; Meng, W.; Chen, Z.; Zhang, N.; Weng, Q.; et al. LncRNA-MM2P Identified as a Modulator of Macrophage M2 Polarization. Cancer Immunol. Res. 2019, 7, 292–305. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Che, Y.; Liu, X.; Liu, C.; Meng, D.; Pang, X.; He, M.; Liu, G.; Zhang, C.; Yang, D.; et al. The Regulating Effect of CII-3 and Its Active Components from Periplaneta americana on M1/M2 Macrophage Polarization. Molecules 2022, 27, 4416. https://doi.org/10.3390/molecules27144416
Xu J, Che Y, Liu X, Liu C, Meng D, Pang X, He M, Liu G, Zhang C, Yang D, et al. The Regulating Effect of CII-3 and Its Active Components from Periplaneta americana on M1/M2 Macrophage Polarization. Molecules. 2022; 27(14):4416. https://doi.org/10.3390/molecules27144416
Chicago/Turabian StyleXu, Jinglei, Yihao Che, Xinyue Liu, Chaohe Liu, Di Meng, Xiuqin Pang, Miao He, Guangming Liu, Chenggui Zhang, Dasong Yang, and et al. 2022. "The Regulating Effect of CII-3 and Its Active Components from Periplaneta americana on M1/M2 Macrophage Polarization" Molecules 27, no. 14: 4416. https://doi.org/10.3390/molecules27144416
APA StyleXu, J., Che, Y., Liu, X., Liu, C., Meng, D., Pang, X., He, M., Liu, G., Zhang, C., Yang, D., & Xiao, H. (2022). The Regulating Effect of CII-3 and Its Active Components from Periplaneta americana on M1/M2 Macrophage Polarization. Molecules, 27(14), 4416. https://doi.org/10.3390/molecules27144416