
Sequential Extraction of Proanthocyanidin Fractions from Ficus Species and Their Effects on Rumen Enzyme Activities In Vitro

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
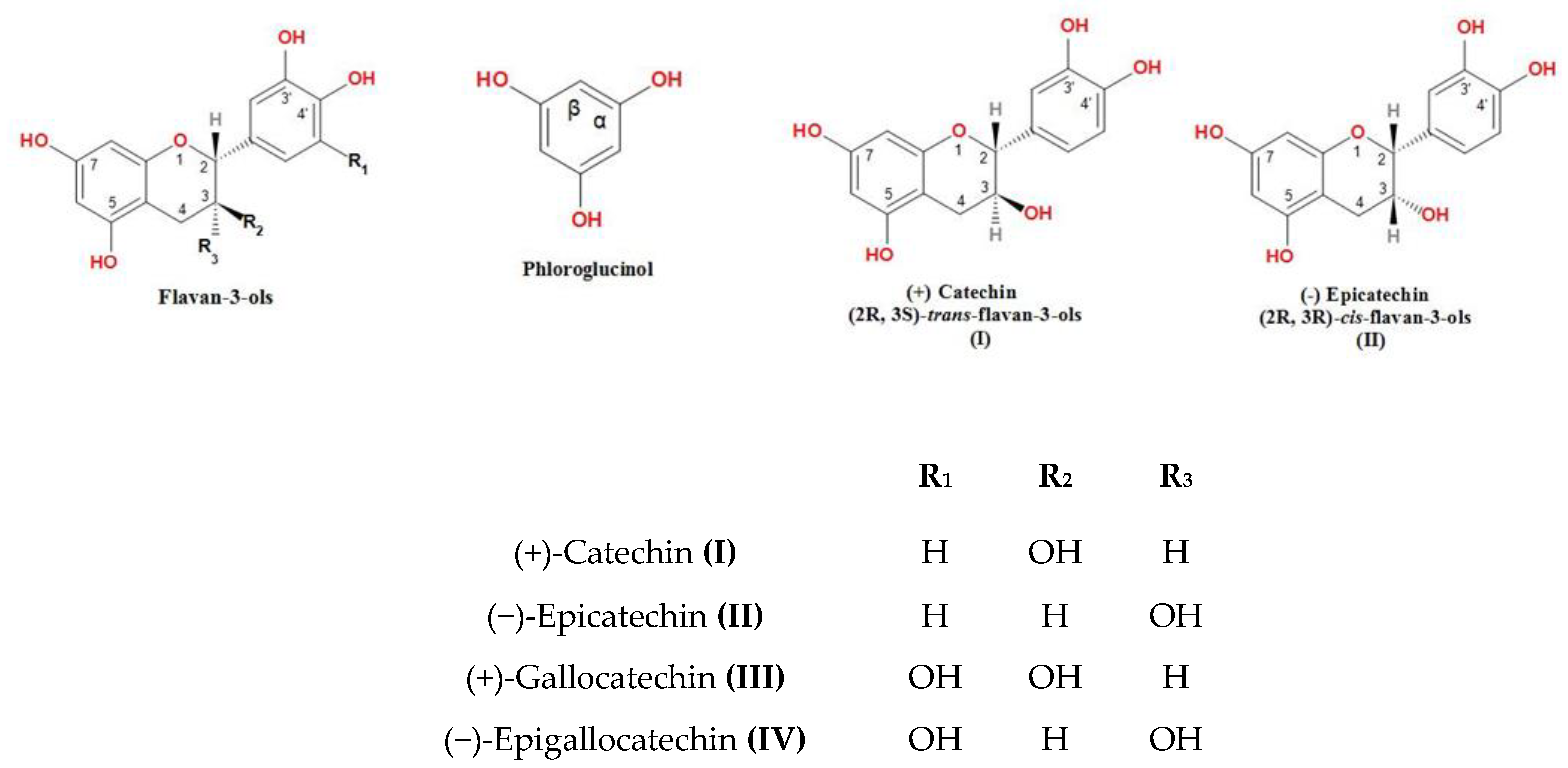
2.1. Extraction and Molecular Characterization of Proanthocyanidins
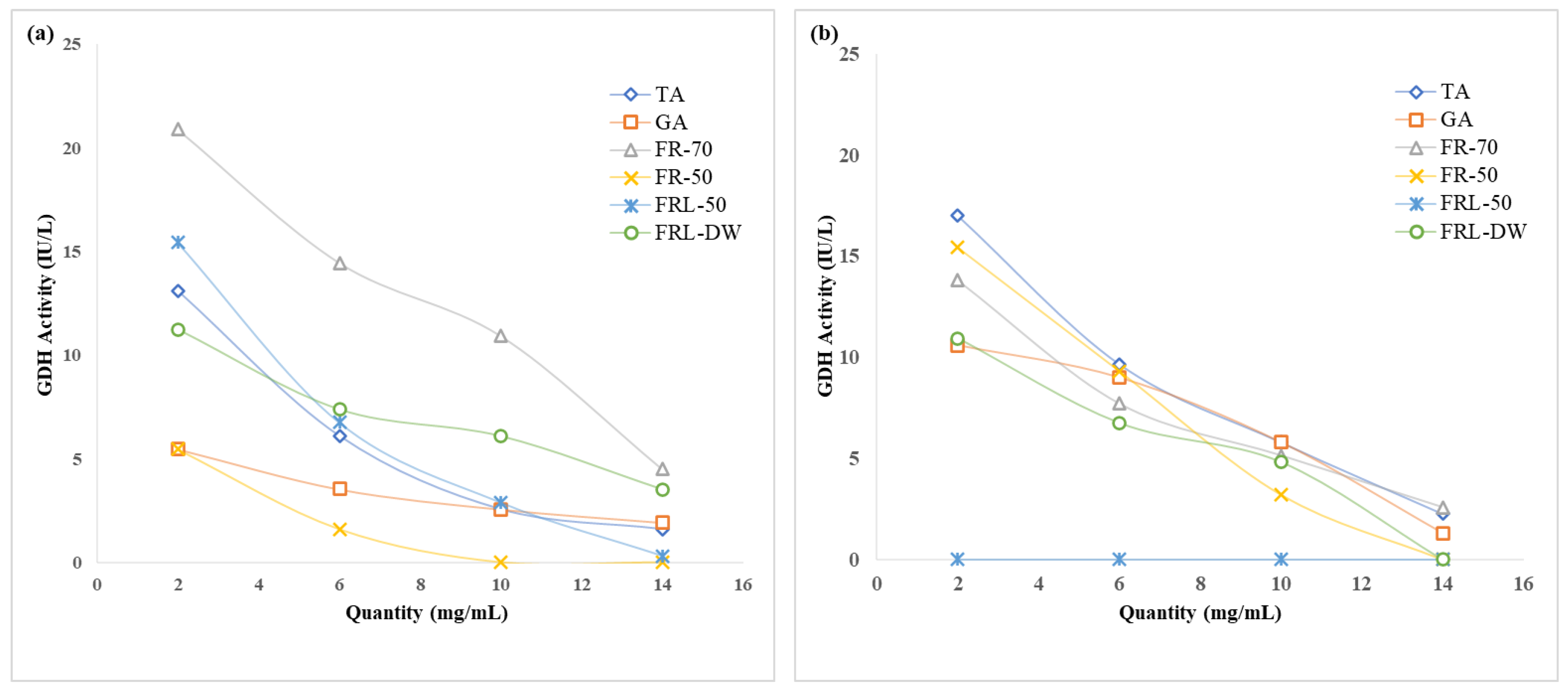
2.2. Effect of Proanthocyanidins on Ruminal Enzymatic Activities
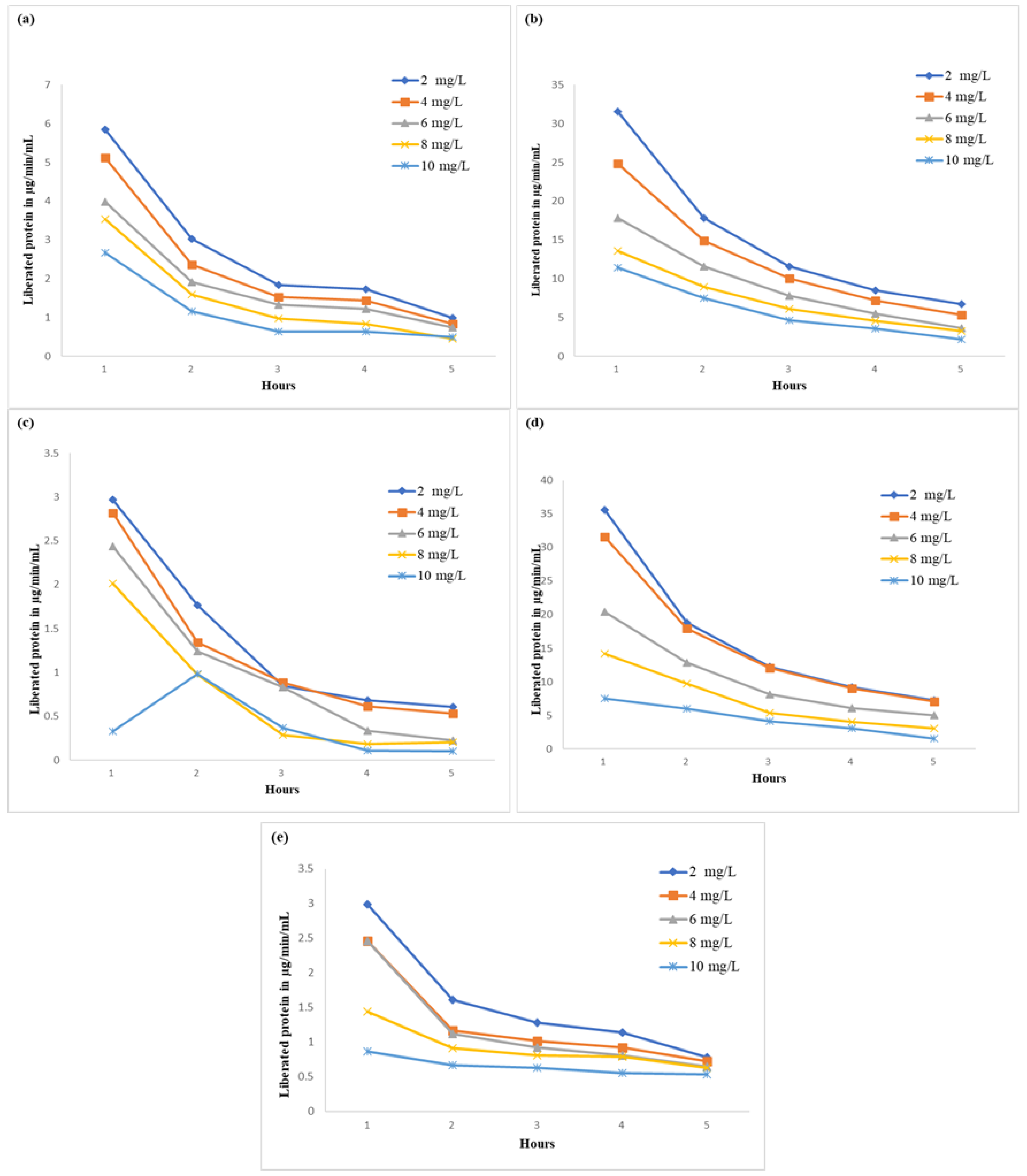
2.3. Effect on Principal Protein (RUBISCO) Solubilization and Microbial Protein Efficiency
3. Materials and Methods
3.1. Reagents
3.2. Plant Materials
3.3. Extraction and Molecular Characterization of Proanthocyanidins
3.4. Ruminal Enzymatic Activities
3.5. Principal Protein (RUBISCO) Solubilization and Microbial Protein Efficiency
3.6. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Leterme, P.; Londoño, A.M.; Estrada, F.; Souffrant, W.B.; Buldgen, A. Chemical composition, nutritive value and voluntary intake of tropical tree foliage and cocoyam in pigs. J. Sci. Food Agric. 2005, 85, 1725–1732. [Google Scholar] [CrossRef]
- Gemede, H.F.; Ratta, N. Antinutritional factors in plant foods: Potential health benefits and adverse effects. Int. J. Food Sci. Nutr. 2014, 3, 284–289. [Google Scholar] [CrossRef]
- Singh, S.; Bhadoria, B.K.; Koli, P.; Singh, A. Nutritional evaluation of top foliages for livestock feeding in semi arid region of India. Indian J. Anim. Sci. 2019, 89, 1389–1398. [Google Scholar]
- Singh, S.; Bhadoria, B.K.; Koli, P.; Lata, S. Seasonal variation in chemical and biochemical constituents of tropical top feed species: Components in silvipasture system. Range Manag. Agrofor. 2021, 42, 312–319. [Google Scholar]
- Koupai-Abyazani, M.R.; McCallum, J.; Bohm, B.A. Identification of the constituent flavanoid units in sainfoin proanthocyanidins by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1992, 594, 117–123. [Google Scholar] [CrossRef]
- Porter, L.J. Flavans and Proanthocyanidins; Routledge: London, UK, 2017; pp. 23–55. [Google Scholar]
- Diwani, N.; Fakhfakh, J.; Athmouni, K.; Belhaj, D.; El Feki, A.; Allouche, N.; Ayadi, H.; Bouaziz-Ketata, H. Optimization, extraction, structure analysis and antioxidant properties of flavan-3-ol polymers: Proanthocyanidins isolated from Periploca angustifolia using surface response methodology. Ind. Crop. Prod. 2020, 144, 112040. [Google Scholar] [CrossRef]
- Alexovič, M.; Dotsikas, Y.; Bober, P.; Sabo, J. Achievements in robotic automation of solvent extraction and related approaches for bioanalysis of pharmaceuticals. J. Chromatogr. B 2018, 1092, 402–421. [Google Scholar] [CrossRef]
- Žitek, T.; Postružnik, V.; Knez, Ž.; Golle, A.; Dariš, B.; Marevci, M.K. Arnica montana L. Supercritical Extraction Optimization for Antibiotic and Anticancer Activity. Front. Bioeng. Biotechnol. 2022, 10, 897185. [Google Scholar] [CrossRef]
- Hammerstone, J.F.; Lazarus, S.A.; Mitchell, A.E.; Rucker, A.R.; Schmitz, H.H. Identification of Procyanidins in Cocoa (Theobroma cacao) and Chocolate Using High-Performance Liquid Chromatography/Mass Spectrometry. J. Agric. Food Chem. 1999, 47, 490–496. [Google Scholar] [CrossRef]
- McMurrough, I.; Madigan, D.; Smyth, M.R. Semipreparative Chromatographic Procedure for the Isolation of Dimeric and Trimeric Proanthocyanidins from Barley. J. Agric. Food Chem. 1996, 44, 1731–1735. [Google Scholar] [CrossRef]
- Markham, K. Isolation techniques for flavonoids. In The flavonoids; Springer: Boston, MA, USA, 1975; pp. 1–44. [Google Scholar]
- Zlatkis, A.; Kaiser, R.E. HPTLC-High Performance Thin-Layer Chromatography; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Kholif, A.; Elghandour, M.; Rodríguez, G.; Olafadehan, O.; Salem, A. Anaerobic ensiling of raw agricultural waste with a fibrolytic enzyme cocktail as a cleaner and sustainable biological product. J. Clean. Prod. 2017, 142, 2649–2655. [Google Scholar] [CrossRef]
- Serraino, I.; Dugo, L.; Dugo, P.; Mondello, L.; Mazzon, E.; Dugo, G.; Caputi, A.P.; Cuzzocrea, S. Protective effects of cyanidin-3-O-glucoside from blackberry extract against peroxynitrite-induced endothelial dysfunction and vascular failure. Life Sci. 2003, 73, 1097–1114. [Google Scholar] [CrossRef]
- Jafari, S.; Goh, Y.M.; Rajion, M.A.; Jahromi, M.F.; Ahmad, Y.H.; Ebrahimi, M. Papaya (Carica papaya) leaf methanolic extract modulates in vitro rumen methanogenesis and rumen biohydrogenation. Anim. Sci. J. 2017, 88, 267–276. [Google Scholar] [CrossRef]
- Aiman-Zakaria, A.; Yong-Meng, G.; Ali-Rajion, M.; Jafari, S.; Faseleh-Jahromi, M.; Shokriyazdan, P.; Ebrahimi, M. The influence of plant polyphenols from oil palm (Elaeis guineensis Jacq.) leaf extract on fermentation characteristics, biohydrogenation of C18 PUFA, and microbial populations in rumen of goats: In vitro study. Acta Agric. Scand. Sect. A Anim. Sci. 2017, 67, 76–84. [Google Scholar] [CrossRef]
- Jayanegara, A.; Togtokhbayar, N.; Makkar, H.P.; Becker, K. Tannins determined by various methods as predictors of methane production reduction potential of plants by an in vitro rumen fermentation system. Anim. Feed Sci. Technol. 2009, 150, 230–237. [Google Scholar] [CrossRef]
- Hatew, B.; Stringano, E.; Mueller-Harvey, I.; Hendriks, W.H.; Carbonero, C.H.; Smith, L.M.J.; Pellikaan, W.F. Impact of variation in structure of condensed tannins from sainfoin (Onobrychis viciifolia) on in vitro ruminal methane production and fermentation characteristics. J. Anim. Physiol. Anim. Nutr. 2016, 100, 348–360. [Google Scholar] [CrossRef]
- Kökten, K.; Kaplan, M.; Hatipoğlu, R.; Saruhan, V.; Çınar, S. Nutritive value of Mediterranean shrubs. J. Anim. Plant Sci. 2012, 22, 188–194. [Google Scholar]
- Barakat, N.A.M.; Laudadio, V.; Cazzato, E.; Tufarelli, V. Potential contribution of Retama raetam (Forssk.) Webb & Berthel as a forage shrub in Sinai, Egypt. Arid Land Res. Manag. 2013, 27, 257–271. [Google Scholar]
- Das, A.; Singh, G.P. Effect of partial replacement of groundnut cake with berseem (Trifolium alexandrinum) on nutrient digestibility, rumen fermentation, pool size and passage rate in cattle fed wheat straw based diet. Anim. Nutr. Feed Technol. 2006, 6, 19–28. [Google Scholar]
- Terrill, T.H.; Windham, W.R.; Hoveland, C.S.; Amos, H.E. Forage Preservation Method Influences on Tannin Concentration, Intake, and Digestibility of Sericea Lespedeza by Sheep. Agron. J. 1989, 81, 435–439. [Google Scholar] [CrossRef]
- Chigayo, K.; Mojapelo, P.E.L.; Mnyakeni-Moleele, S.; Misihairabgwi, J.M. Phytochemical and antioxidant properties of different solvent extracts of Kirkia wilmsii tubers. Asian Pac. J. Trop. Biomed. 2016, 6, 1037–1043. [Google Scholar] [CrossRef]
- Merghem, R.; Jay, M.; Brun, N.; Voirin, B. Qualitative analysis and HPLC isolation and identification of procyanidins from Vicia faba. Phytochem. Anal. 2004, 15, 95–99. [Google Scholar] [CrossRef]
- Wen, L.; Wu, D.; Jiang, Y.; Prasad, K.N.; Lin, S.; Jiang, G.; He, J.; Zhao, M.; Luo, W.; Yang, B. Identification of flavonoids in litchi (Litchi chinensis Sonn.) leaf and evaluation of anticancer activities. J. Funct. Foods 2014, 6, 555–563. [Google Scholar] [CrossRef]
- Dhanani, T.; Singh, R.; Kumar, S. Extraction optimization of gallic acid,(+)-catechin, procyanidin-B2, (−)-epicatechin, (−)-epigallocatechin gallate, and (−)-epicatechin gallate: Their simultaneous identification and quantification in Saraca asoca. J. Food Drug Anal. 2017, 25, 691–698. [Google Scholar] [CrossRef]
- Orejola, J.; Matsuo, Y.; Saito, Y.; Tanaka, T. Characterization of Proanthocyanidin Oligomers of Ephedra sinica. Molecules 2017, 22, 1308. [Google Scholar] [CrossRef]
- Moridani, M.Y.; Pourahmad, J.; Bui, H.; Siraki, A.; O’Brien, P.J. Dietary flavonoid iron complexes as cytoprotective superoxide radical scavengers. Free Radic. Biol. Med. 2003, 34, 243–253. [Google Scholar] [CrossRef]
- Oliveira, J.; Mateus, N.; de Freitas, V. Flavanols: Catechins and Proanthocyanidins. In Natural Products; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1753–1801. [Google Scholar] [CrossRef]
- Cuevas-Rodríguez, E.O.; Yousef, G.G.; García-Saucedo, P.A.; López-Medina, J.; Paredes-López, O.; Lila, M.A. Characterization of Anthocyanins and Proanthocyanidins in Wild and Domesticated Mexican Blackberries (Rubus spp.). J. Agric. Food Chem. 2010, 58, 7458–7464. [Google Scholar] [CrossRef]
- Oskoueian, E.; Abdullah, N.; Oskoueian, A. Effects of Flavonoids on Rumen Fermentation Activity, Methane Production, and Microbial Population. BioMed Res. Int. 2013, 2013, 349129. [Google Scholar] [CrossRef]
- McAllister, T.A.; Martinez, T.; Bae, H.D.; Muir, A.D.; Yanke, L.J.; Jones, G.A. Characterization of Condensed Tannins Purified From Legume Forages: Chromophore Production, Protein Precipitation, and Inhibitory Effects on Cellulose Digestion. J. Chem. Ecol. 2005, 31, 2049–2068. [Google Scholar] [CrossRef]
- Raza, M.; Shahwar, D.; Khan, T. Radical Scavenging, Proteases Activities, and Phenolics Composition of Bark Extracts from 21 Medicinal Plants. J. Chem. 2015, 2015, 951840. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 2010, 91, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.A.; McAllister, T.A.; Muir, A.D.; Cheng, K.J. Effects of sainfoin (Onobrychis viciifolia Scop.) condensed tannins on growth and proteolysis by four strains of ruminal bacteria. Appl. Environ. Microbiol. 1994, 60, 1374–1378. [Google Scholar] [CrossRef] [PubMed]
- McManus, J.P.; Davis, K.G.; Lilley, T.H.; Haslam, E. The association of proteins with polyphenols. J. Chem. Soc. Chem. Commun. 1981, 7, 309b–311b. [Google Scholar] [CrossRef]
- Aerts, R.J.; Barry, T.N.; McNabb, W. Polyphenols and agriculture: Beneficial effects of proanthocyanidins in forages. Agric. Ecosyst. Environ. 1999, 75, 1–12. [Google Scholar] [CrossRef]
- Jonker, A.; Yu, P. The Occurrence, Biosynthesis, and Molecular Structure of Proanthocyanidins and Their Effects on Legume Forage Protein Precipitation, Digestion and Absorption in the Ruminant Digestive Tract. Int. J. Mol. Sci. 2017, 18, 1105. [Google Scholar] [CrossRef] [PubMed]
- Min, B.R.; McNabb, W.C.; Barry, T.N.; Peters, J.S. Solubilization and degradation of ribulose-1,5-bisphosphate carboxylase/oxygenase (EC 4.1.1.39; Rubisco) protein from white clover (Trifolium repens) and Lotus corniculatus by rumen microorganisms and the effect of condensed tannins on these processes. J. Agri. Sci. 2000, 134, 305–317. [Google Scholar] [CrossRef]
- Mangan, J.L. 2.1 The Nitrogenous Constituents of Fresh Forages. BSAP Occas. Publ. 1982, 6, 25–40. [Google Scholar] [CrossRef]
- Waghorn, G.C.; McNabb, W. Consequences of plant phenolic compounds for productivity and health of ruminants. Proc. Nutr. Soc. 2003, 62, 383–392. [Google Scholar] [CrossRef]
- Bento, M.; Makkar, H.; Acamovic, T. Effect of mimosa tannin and pectin on microbial protein synthesis and gas production during in vitro fermentation of 15N-labelled maize shoots. Anim. Feed Sci. Technol. 2005, 123–124, 365–377. [Google Scholar] [CrossRef]
- Getachew, G.; Pittroff, W.; Putnam, D.; Dandekar, A.; Goyal, S.; DePeters, E. The influence of addition of gallic acid, tannic acid, or quebracho tannins to alfalfa hay on in vitro rumen fermentation and microbial protein synthesis. Anim. Feed Sci. Technol. 2008, 140, 444–461. [Google Scholar] [CrossRef]
- Mandels, M.; Weber, J. The Production of Cellulases; ACS Publications: Washington, DC, USA, 1969; pp. 391–414. [Google Scholar]
- Tsubota, H.; Hoshino, S. Transaminase Activity in Sheep Rumen Content. J. Dairy Sci. 1969, 52, 2024–2028. [Google Scholar] [CrossRef]
- Yatzidis, H. Measurement of Transaminases in Serum. Nature 1960, 186, 79–80. [Google Scholar] [CrossRef]
- Strecker, H.J. L-Glutamic dehydrogenase from liver. Meth. Enzymol. 1955, 2, 220–221. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Blackburn, T.H.; Hobson, P.N. Isolation of proteolytic bacteria from the sheep rumen. Microbiology 1960, 22, 282–289. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, M. Tannins: Their adverse role in ruminant nutrition. J. Agric. Food Chem. 1984, 32, 447–453. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bradstreet, R.B. Kjeldahl method for organic nitrogen. Anal. Chem. 1954, 26, 185–187. [Google Scholar] [CrossRef]
- Zinn, R.A.; Owens, F.N. A rapid procedure for purine measurement and its use for estimating net ruminal protein synthesis. Can. J. Anim. Sci. 1986, 66, 157–166. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components (mg/g) | FRL-50 * | FRL-DW | FR-70 | FR-50 | FR-DW |
|---|---|---|---|---|---|
| (+) Catechin | 0.151 ± 0.01 b | N.D. | 0.098 ± 0.01 c | N.D. | 0.524 ± 0.03 a |
| (−) Epicatechin | 0.317 ± 0.02 a | 0.181 ± 0.03 bc | 0.135 ± 0.04 c | 0.235 ± 0.04 ab | 0.221 ± 0.02 bc |
| (+) Gallocatechin | 0.092 ± 0.01 cd | 0.249 ± 0.06 bc | 0.458 ± 0.13 a | 0.347 ± 0.05 ab | 0.035 ± 0.02 d |
| (−) Epigallocatechin | 0.117 ± 0.09 ab | 0.056 ± 0.05 bd | 0.065 ± 0.01 c | 0.016 ± 0.07 cd | 0.315 ± 0.11 a |
| (+) Catechin-4-phloroglucinol | 0.963 ± 0.07 b | N.D. | 1.461 ± 0.12 a | 0.990 ± 0.22 b | 1.361 ± 0.07 b |
| (+) Gallocatechin-4-phloroglucinol | 0.649 ± 0.09 b | 0.873 ± 0.11 b | 1.321 ± 0.17 a | 0.550 ± 0.09 c | 0.555 ± 0.10 c |
| (−) Epigallocatechin-4-phloroglucinol | 0.458 ± 0.04 b | 0.168 ± 0.05 c | 0.974 ± 0.12 a | 1.130 ± 0.10 a | 0.947 ± 0.08 a |
| Total monomer | 0.676 | 0.51 | 0.745 | 0.773 | 1.093 |
| Total adduct | 2.066 | 2.03 | 3.789 | 2.678 | 2.875 |
| Degree of polymerization | 3.06 | 3.99 | 5.09 | 3.46 | 2.63 |
| Subunits ratio (Dp:Cy) | 50:50 | 50:50 | 50:50 | 50:50 | 50:50 |
| Enzymes (EC50 mg/mL) | TA | GA | FRL-50 | FRL-DW | FR-70 | FR-50 | |
|---|---|---|---|---|---|---|---|
| R-GOT | P | 20.54 ± 1.13 a | 21.77 ± 0.80 a | 4.72 ± 0.42 d | 10.67 ± 1.12 c | 17.17 ± 1.01 b | 12.33 ± 1.26 c |
| B | 17.54 ± 0.13 bc | 22.78 ± 0.80 a | 7.12 ± 0.02 d | 7.69 ± 1.25 d | 19.17 ± 1.00 b | 16.33 ± 0.50 c | |
| R-GPT | P | 17.52 ± 0.70 a | 6.89 ± 0.12 cd | 5.44 ± 0.49 d | 13.59 ± 1.42 b | 16.42 ± 1.07 a | 8.61 ± 0.96 c |
| B | 17.12 ± 0.15 a | 15.52 ± 0.53 a | 5.28 ± 0.63 b | 16.69 ± 1.44 a | 15.35 ± 0.82 a | 5.31 ± 0.84 b | |
| R-Cellulase | P + B | 124.32 ± 1.63 a | 6.62 ± 0.56 d | N.O. | N.O. | 24.72 ± 1.25 b | 9.70 ± 0.54 c |
| Diet | Solubilization (%) | NRNAQ | |
|---|---|---|---|
| FRL | 22.36 ± 1.24 bc | 0.21 ± 0.02 bcd | |
| FR | 17.26 ± 0.61 e | 0.22 ± 0.01 ab | |
| WS | 8.95 ± 0.95 gh | 0.16 ± 0.01 ef | |
| BH | 3.04 ± 0.08 i | 0.17 ± 0.01 cdef | |
| FR + WS | 1:1 | 16.61 ± 1.04 e | 0.15 ± 0.01 f |
| 2:1 | 28.58 ± 1.48 a | 0.22 ± 0.01 ab | |
| 3:1 | 15.29 ± 0.61 ef | 0.19 ± 0.01 bcde | |
| FR + BH | 1:1 | 18.35 ± 1.48 de | 0.26 ± 0.03 a |
| 2:1 | 21.70 ± 3.00 cd | 0.21 ± 0.01 bc | |
| 3:1 | 11.81 ± 1.01 fg | 0.21 ± 0.02 bc | |
| FRL + WS | 1:1 | 27.95 ± 1.95 a | 0.19 ± 0.01 bcdef |
| 2:1 | 22.47 ± 1.37 bc | 0.16 ± 0.01 ef | |
| 3:1 | 5.59 ± 0.34 hi | 0.18 ± 0.01 cdef | |
| FRL + BH | 1:1 | 14.47 ± 0.74 ef | 0.22 ± 0.01 ab |
| 2:1 | 17.80 ± 1.26 de | 0.17 ± 0.01 def | |
| 3:1 | 25.90 ± 1.18 ab | 0.17 ± 0.0 ef | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koli, P.; Singh, S.; Bhadoria, B.K.; Agarwal, M.; Lata, S.; Ren, Y. Sequential Extraction of Proanthocyanidin Fractions from Ficus Species and Their Effects on Rumen Enzyme Activities In Vitro. Molecules 2022, 27, 5153. https://doi.org/10.3390/molecules27165153
Koli P, Singh S, Bhadoria BK, Agarwal M, Lata S, Ren Y. Sequential Extraction of Proanthocyanidin Fractions from Ficus Species and Their Effects on Rumen Enzyme Activities In Vitro. Molecules. 2022; 27(16):5153. https://doi.org/10.3390/molecules27165153
Chicago/Turabian StyleKoli, Pushpendra, Sultan Singh, Brijesh K. Bhadoria, Manjree Agarwal, Suman Lata, and Yonglin Ren. 2022. "Sequential Extraction of Proanthocyanidin Fractions from Ficus Species and Their Effects on Rumen Enzyme Activities In Vitro" Molecules 27, no. 16: 5153. https://doi.org/10.3390/molecules27165153
APA StyleKoli, P., Singh, S., Bhadoria, B. K., Agarwal, M., Lata, S., & Ren, Y. (2022). Sequential Extraction of Proanthocyanidin Fractions from Ficus Species and Their Effects on Rumen Enzyme Activities In Vitro. Molecules, 27(16), 5153. https://doi.org/10.3390/molecules27165153