



Synthesis and Biological Evaluation of Cassane Diterpene (5α)-Vuacapane-8(14), 9(11)-Diene and of Some Related Compounds

 , ,
, ,  ,
,  , ,
, ,
Abstract
:1. Introduction
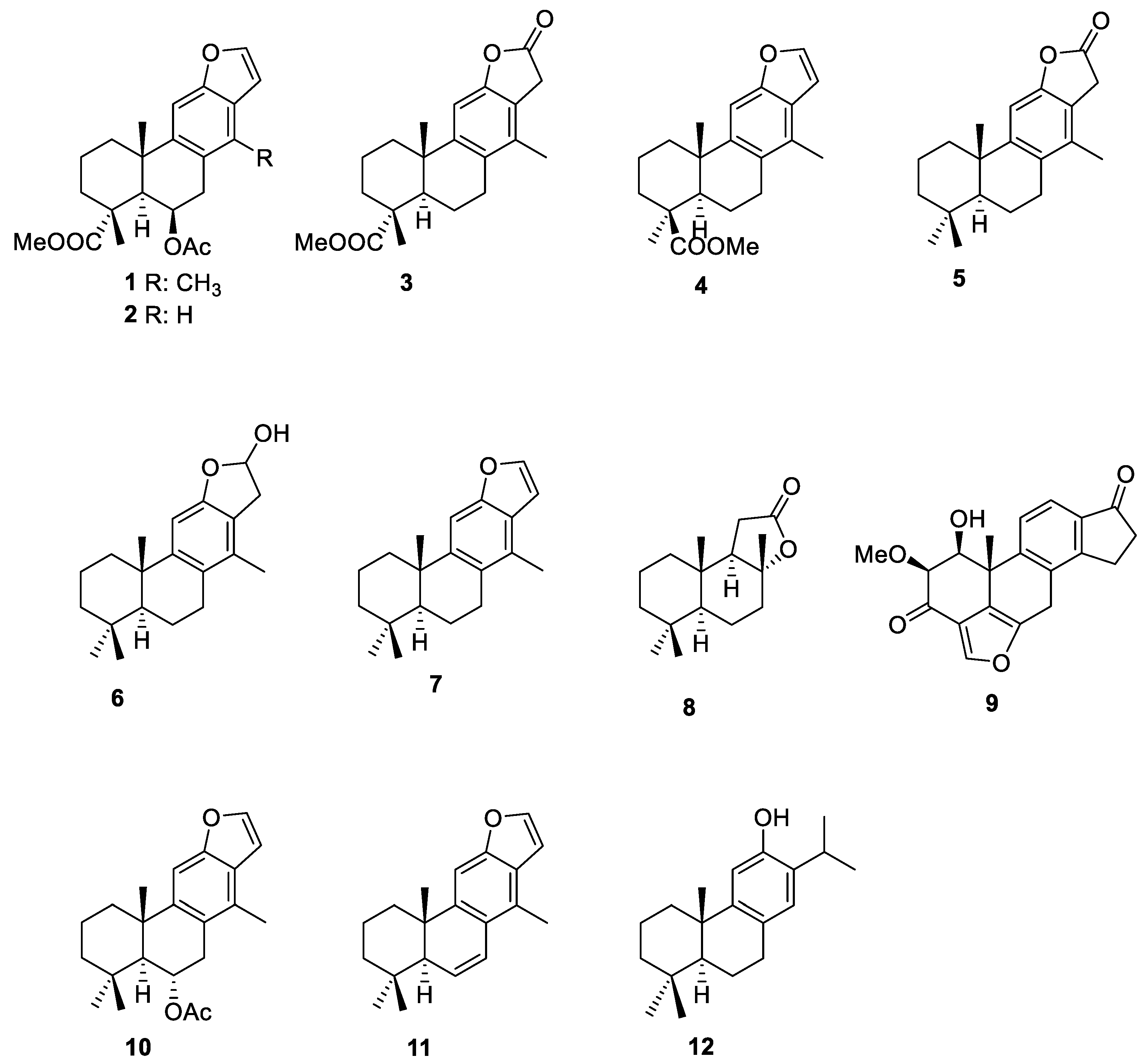
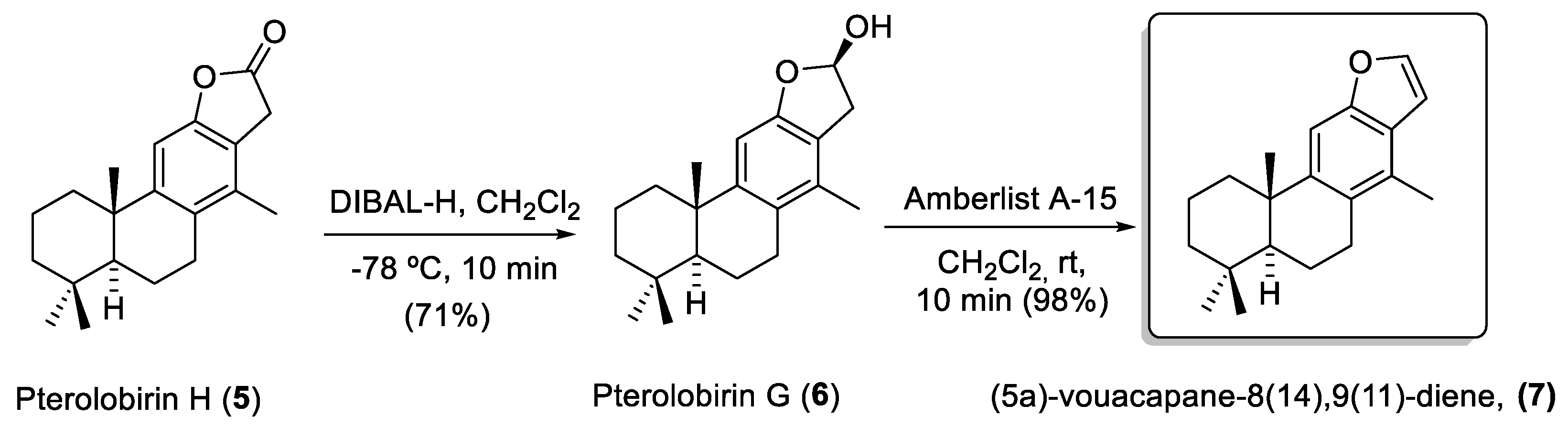
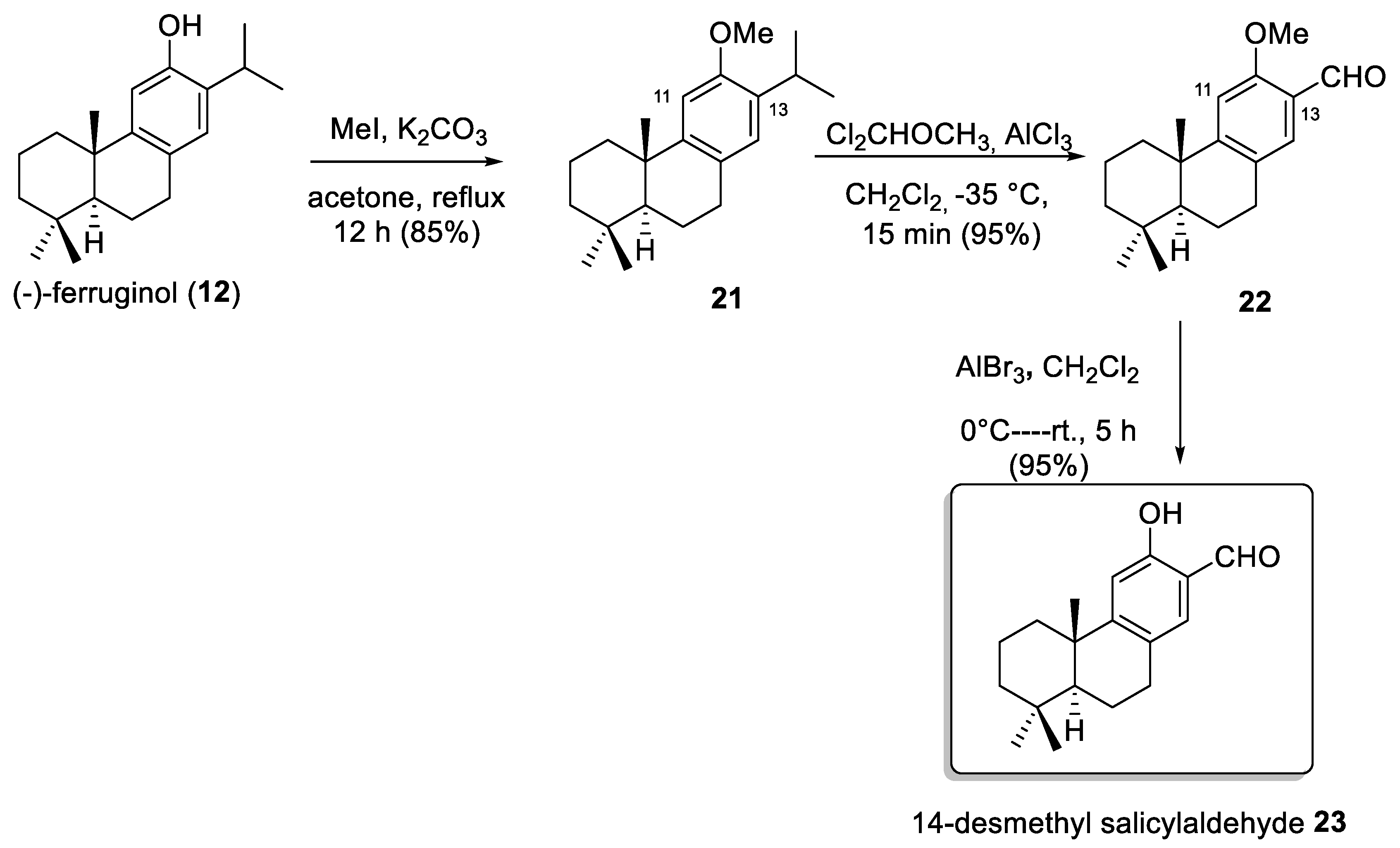
2. Results and Discussion
2.1. Chemistry
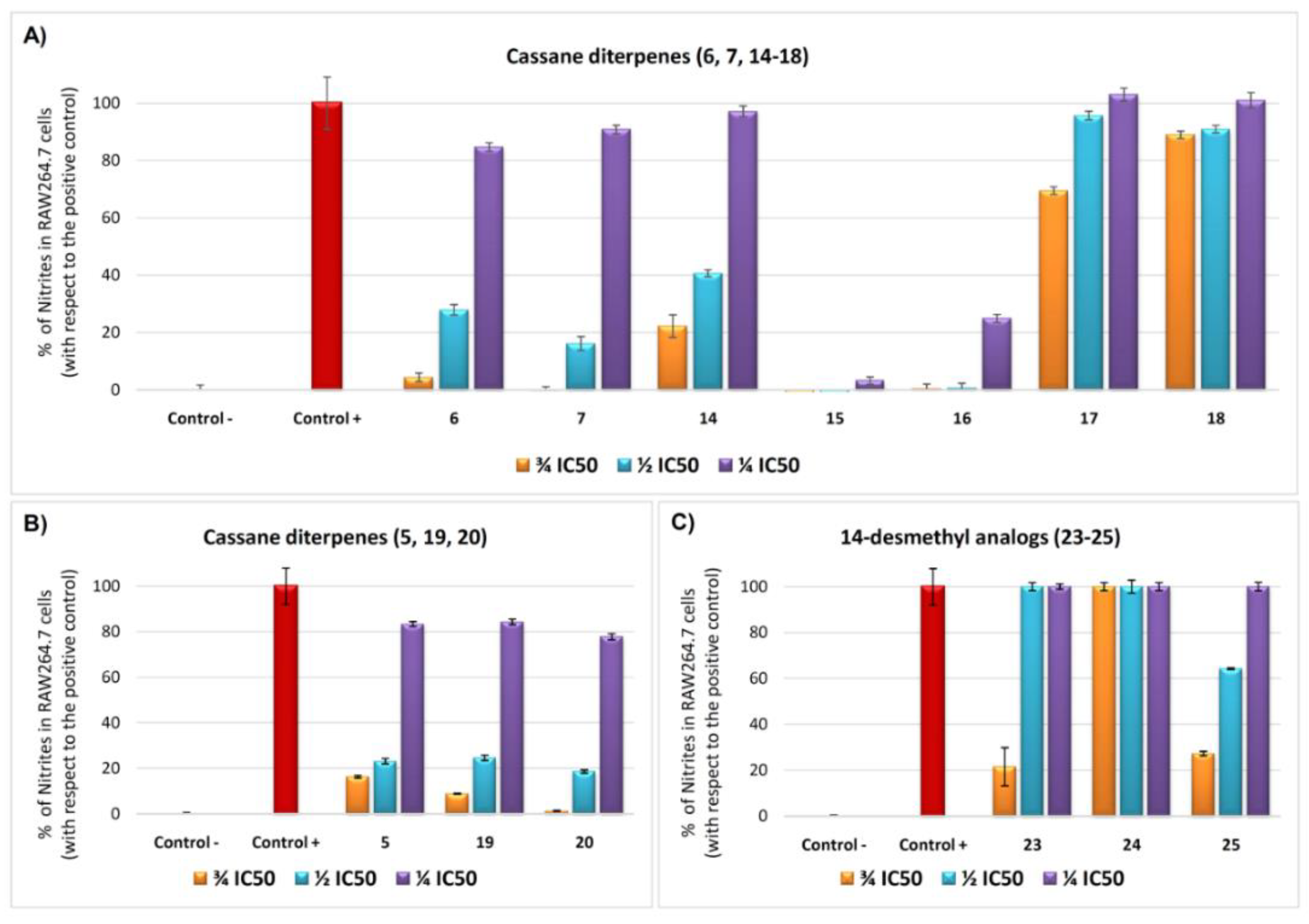
2.2. Biological Assays
2.2.1. Cytotoxicity on RAW 264.7 Cell Line
2.2.2. Effects on Inhibition of NO Production
2.2.3. Effects on RAW 264.7 Cell Cycle Arrest and Distribution
2.2.4. Cell Cytotoxicity Assay and SAR
2.2.5. Effects on HT29 Cell Cycle Arrest and Distribution
2.2.6. Characterization of Apoptotic Effects by Flow Cytometry with Annexin-V
2.2.7. Effects on Changes in Mitochondrial-Membrane Potential
3. Experimental Section
3.1. Synthesis
3.2. Biological Experimental Procedures
3.2.1. Test Compounds
3.2.2. Cell Cytotoxicity Assay
3.2.3. Measurement of Nitric Oxide Concentration
3.2.4. RAW264.7 Cell Cycle
3.2.5. HT29 Cell Cycle
3.2.6. Annexin V-FICT/Propidium Iodide Flow-Cytometry Analysis
3.2.7. Flow-Cytometry Analysis of the Mitochondrial Membrane Potential
3.2.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Jing, W.; Zhang, X.X.; Zhou, H.; Wang, Y.; Yang, M.; Long, L.; Gao, H. Naturally Occurring Cassane Diterpenoids (CAs) of Caesalpinia: A Systematic Review of Its Biosynthesis, Chemistry, and Pharmacology. Fitoterapia 2019, 134, 226–249. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, S.W.; Xuan, L.J. Trinorcassane and Cassane Diterpenoids from the Seeds of Caesalpinia Minax. Fitoterapia 2015, 102, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Maurya, R.; Ravi, M.; Singh, S.; Yadav, P.P. A Review on Cassane and Norcassane Diterpenes and Their Pharmacological Studies. Fitoterapia 2012, 83, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Ochieng’, C.O.; Owuor, P.O.; Mang’Uro, L.A.O.; Akala, H.; Ishola, I.O. Antinociceptive and Antiplasmodial Activities of Cassane Furanoditerpenes from Caesalpinia Volkensii H. Root Bark. Fitoterapia 2012, 83, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, A.; Chatzopoulou, M.; Bantzi, M.; Athanassopoulos, C.M.; Giannis, A.; Pitsinos, E.N. Identification of Gli-Mediated Transcription Inhibitors through Synthesis and Evaluation of Taepeenin D Analogues. Medchemcomm 2016, 7, 2328–2331. [Google Scholar] [CrossRef]
- Chatzopoulou, M.; Antoniou, A.; Pitsinos, E.N.; Bantzi, M.; Koulocheri, S.D.; Haroutounian, S.A.; Giannis, A. A Fast Entry to Furanoditerpenoid-Based Hedgehog Signaling Inhibitors: Identifying Essential Structural Features. Org. Lett. 2014, 16, 3344–3347. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, P.; Altarejos, J.; Linares-Palomino, P.J.; Chahboun, R.; Alvarez-Manzaneda, E. Synthesis of Cassane-Type Diterpenes from Abietane Compounds: The First Synthesis of Taepeenin F. Org. Chem. Front. 2018, 5, 2542–2546. [Google Scholar] [CrossRef]
- Mahdjour, S.; Harche-Kaid, M.; Haidour, A.; Chahboun, R.; Alvarez-Manzaneda, E. Short Route to Cassane-Type Diterpenoids: Synthesis of the Supposed Structure of Benthaminin 1. Org. Lett. 2016, 18, 5964–5967. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.A.; Houghton, P.J.; Hylands, P.J. Antibacterial and Antioxidant Cassane Diterpenoids from Caesalpinia Benthamiana. Phytochemistry 2007, 68, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Zentar, H.; Arias, F.; Haidour, A.; Alvarez-Manzaneda, R.; Chahboun, R.; Alvarez-Manzaneda, E. Protecting-Group-Free Synthesis of Cassane-Type Furan Diterpenes via a Decarboxylative Dienone-Phenol Rearrangement. Org. Lett. 2018, 20, 7007–7010. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yang, Y.R.; Yin, Y.; Song, K.R.; Long, L.P.; Li, X.Z.; Zhou, B.; Gao, H.Y. New Cassane Diterpenoids from the Seed Kernels of Caesalpinia Cucullata, Exhibit Anti-Inflammatory Effect in Vitro by Inhibiting INOS Enzymatic Activity. Chin. J. Chem. 2021, 39, 1625–1634. [Google Scholar] [CrossRef]
- Anderson, E.A.; Alexanian, E.J.; Sorensen, E.J. Synthesis of the furanosteroidal antibiotic viridin. Angew. Chem. Int. Ed. 2004, 43, 1998–2001. [Google Scholar] [CrossRef] [PubMed]
- Del Bel, M.; Abela, A.R.; Ng, J.D.; Guerrero, C.A. Enantioselective Chemical Syntheses of the Furanosteroids (-)-Viridin and (-)-Viridiol. J. Am. Chem. Soc. 2017, 139, 6819–6822. [Google Scholar] [CrossRef] [PubMed]
- Pitsinos, E.N.; Mavridis, I.; Tzouma, E.; Vidali, V.P. Enantioselective Synthesis of Cassane-Type Furanoditerpenoids: Total Synthesis of Sucutiniranes C and D. Eur. J. Org. Chem. 2020, 2020, 4730–4742. [Google Scholar] [CrossRef]
- Hanson, J.R. The Viridin Family of Steroidal Antibiotics. Nat. Prod. Rep. 1995, 12, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Raksat, A.; Choodej, S.; Aree, T.; Nejad Ebrahimi, S.; Pudhom, K. Cassane-Type Diterpenes from Roots of Pterolobium Macropterum and Their Anti-Inflammatory Activity. Phytochemistry 2022, 196, 113074. [Google Scholar] [CrossRef]
- Mori, N.; Kuzuya, K.; Watanabe, H. Synthesis of (-)-Chamobtusin A from (+)-Dehydroabietylamine. J. Org. Chem. 2016, 81, 11866–11870. [Google Scholar] [CrossRef] [PubMed]
- Roa-Linares, V.C.; Brand, Y.M.; Agudelo-Gomez, L.S.; Tangarife-Castaño, V.; Betancur-Galvis, L.A.; Gallego-Gomez, J.C.; González, M.A. Anti-Herpetic and Anti-Dengue Activity of Abietane Ferruginol Analogues Synthesized from (+)-Dehydroabietylamine. Eur. J. Med. Chem. 2016, 108, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Manzaneda, E.; Chahboun, R.; Cabrera, E.; Alvarez, E.; Haidour, A.; Ramos, J.M.; Alvarez-Manzaneda, R.; Hmamouchi, M.; Es-Samti, H. A Thermal 6π Electrocyclization Strategy towards Taiwaniaquinoids. First Enantiospecific Synthesis of (-)-Taiwaniaquinone G. Chem. Commun. 2009, 592–594. [Google Scholar] [CrossRef]
- Alvarez-Manzaneda, E.; Chahboun, R.; Alvarez, E.; Cano, M.J.; Haidour, A.; Alvarez-Manzaneda, R. Enantioselective Total Synthesis of the Selective PI3 Kinase Inhibitor Liphagal. Org. Lett. 2010, 12, 4450–4453. [Google Scholar] [CrossRef]
- Dong, R.; Yuan, J.; Wu, S.; Huang, J.; Xu, X.; Wu, Z.; Gao, H. Anti-Inflammation Furanoditerpenoids from Caesalpinia Minax Hance. Phytochemistry 2015, 117, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Fadok, V.A.; Bratton, D.L.; Frasch, S.C.; Warner, M.L.; Henson, P.M. The Role of Phosphatidylserine in Recognition of Apoptotic Cells by Phagocytes. Cell Death Differ. 1998, 5, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Zhang, L.; Liu, Q.; Feng, L.; Ye, Y.; Lu, J.; Lin, L. Cytotoxic and Pro-Apoptotic Effects of Cassane Diterpenoids from the Seeds of Caesalpinia Sappan in Cancer Cells. Molecules 2016, 21, 791. [Google Scholar] [CrossRef] [PubMed]
- Tran, M.H.; Nguyen, M.T.T.; Nguyen, H.D.; Nguyen, T.D.; Phuong, T.T. Cytotoxic Constituents from the Seeds of Vietnamese Caesalpinia Sappan. Pharm. Biol. 2015, 53, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, Q.; Ma, G.; Chen, L.; Zhang, C.; Chen, B.; Yang, J.; Li, W. Cassane-Type Diterpenes from Caesalpinia Minax Induce Apoptosis in Pituitary Adenoma: Structure–Activity Relationship, ER Stress, and Wnt/β-Catenin Pathways. Asian Nat. Prod. Res. 2017, 19, 423–435. [Google Scholar] [CrossRef]
- Kandasamy, V.; Balasundaram, U. Caesalpinia Bonduc (L.) Roxb. as a Promising Source of Pharmacological Compounds to Treat Poly Cystic Ovary Syndrome (PCOS): A Review. J. Ethnopharmacol. 2021, 279, 114375. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Bryan, N.S.; Grisham, M.B. Methods to Detect Nitric Oxide and Its Metabolites in Biological Samples. Free Radic. Biol. Med. 2007, 43, 645–657. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC20 (μg/mL) | IC50 (μg/mL) | IC80 (μg/mL) |
|---|---|---|---|
| 5 | 34.92 ± 2.11 | 45.34 ± 1.33 | 64.21 ± 3.29 |
| 6 | 12.69 ± 0.30 | 14.99 ± 0.32 | 18.56± 0.45 |
| 7 | 50.92 ±1.61 | 57.55 ± 1.06 | 65.51 ± 0.95 |
| 14 | 22.92 ± 0.13 | 24.67 ± 0.04 | 26.89 ± 0.02 |
| 15 | 43.66 ± 2.37 | 64.62 ± 1.39 | 83.67 ± 0.64 |
| 16 | 14.51 ± 0.68 | 26.72 ± 0.88 | N/A |
| 17 | 4.95 ± 0.13 | 7.49 ± 0.86 | 11.78 ± 2.97 |
| 18 | 7.32 ± 2.53 | 13.11 ± 1.51 | 20.38 ± 0.26 |
| 19 | 70.83 ± 0.28 | 74.65 ± 0.17 | 78.94 ± 0.28 |
| 20 | 7.86 ±3.97 | 8.34 ± 2.51 | 9.35 ± 0.95 |
| 23 | 5.13 ± 0.18 | 5.87 ± 0.18 | 6.80 ± 0.24 |
| 24 | 9.09 ± 0.03 | 13.94 ± 0.07 | 20.75 ± 0.48 |
| 25 | 42.62 ± 0.74 | 53.92 ± 0.36 | 66.33 ± 0.35 |
| Cassane Diterpenes | 14-Desmethyl Analogs | ||||
|---|---|---|---|---|---|
| Comp. | IC50 NO (μg/mL) | Comp. | IC50 NO (μg/mL) | Comp. | IC50 NO (μg/mL) |
| 6 | 4.01 ± 0.07 | 5 | 17.57 ± 0.31 | 23 | 3.32 ± 0.05 |
| 7 | 22.02 ± 0.57 | 19 | 27.57 ± 0.20 | 24 | 15.21 ± 2.13 |
| 14 | 10.71 ± 1.14 | 20 | 2.98 ± 0.04 | 25 | 22.58 ± 0.05 |
| 15 | 7.98 ± 0.10 | ||||
| 16 | 5.71 ± 0.14 | ||||
| 17 | 4.74 ± 1.27 | ||||
| 18 | 31.37 ± 4.90 | ||||
| IC50 (μg/mL) | |||||
|---|---|---|---|---|---|
| Entry | Product | HT29 | HepG2 | B16-F10 | |
| Cassane diterpenes | 1 | 5 | 70.07 ± 1.57 | 45.31 ± 1.81 | 39.71 ± 0.28 |
| 2 | 6 | 3.87 ± 0.98 | 11.75 ± 0.15 | 10.34 ± 0.74 | |
| 3 | 7 | 20.66 ± 1.09 | 13.96 ± 0.50 | 31.73 ± 1.10 | |
| 4 | 14 | 15.43 ± 2.72 | 33.77 ± 0.80 | 9.25 ± 0.74 | |
| 5 | 15 | 25.29 ± 0.89 | 58.32 ± 0.32 | 13.01 ± 1.80 | |
| 6 | 16 | 11.89 ± 0.80 | 26.72 ± 0.88 | 6.01 ± 0.41 | |
| 7 | 17 | 5.96 ± 0.55 | 8.15 ± 0.10 | 5.97 ± 0.55 | |
| 8 | 18 | 30.67 ± 0.49 | 13.57 ± 0.12 | 5.96 ± 0.35 | |
| 9 | 19 | 48.99 ± 0.72 | 12.22 ± 1.78 | 48.82 ± 2.79 | |
| 10 | 20 | 3.54 ± 0.19 | 8.41 ± 0.30 | 2.38 ± 0.39 | |
| Desmethyl analogs | 11 | 23 | 5.19 ± 0.17 | 8.36 ± 1.77 | 4.33 ± 0.27 |
| 12 | 24 | 17.43 ± 1.97 | 24.22 ± 2.71 | 10.51 ± 0.01 | |
| 13 | 25 | 74.26 ± 2.19 | 77.97 ± 0.72 | 74.26 ± 2.19 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zentar, H.; Jannus, F.; Medina-O’Donnell, M.; Lupiáñez, J.A.; Justicia, J.; Alvarez-Manzaneda, R.; Reyes-Zurita, F.J.; Alvarez-Manzaneda, E.; Chahboun, R. Synthesis and Biological Evaluation of Cassane Diterpene (5α)-Vuacapane-8(14), 9(11)-Diene and of Some Related Compounds. Molecules 2022, 27, 5705. https://doi.org/10.3390/molecules27175705
Zentar H, Jannus F, Medina-O’Donnell M, Lupiáñez JA, Justicia J, Alvarez-Manzaneda R, Reyes-Zurita FJ, Alvarez-Manzaneda E, Chahboun R. Synthesis and Biological Evaluation of Cassane Diterpene (5α)-Vuacapane-8(14), 9(11)-Diene and of Some Related Compounds. Molecules. 2022; 27(17):5705. https://doi.org/10.3390/molecules27175705
Chicago/Turabian StyleZentar, Houda, Fatin Jannus, Marta Medina-O’Donnell, José A. Lupiáñez, José Justicia, Ramón Alvarez-Manzaneda, Fernando J. Reyes-Zurita, Enrique Alvarez-Manzaneda, and Rachid Chahboun. 2022. "Synthesis and Biological Evaluation of Cassane Diterpene (5α)-Vuacapane-8(14), 9(11)-Diene and of Some Related Compounds" Molecules 27, no. 17: 5705. https://doi.org/10.3390/molecules27175705
APA StyleZentar, H., Jannus, F., Medina-O’Donnell, M., Lupiáñez, J. A., Justicia, J., Alvarez-Manzaneda, R., Reyes-Zurita, F. J., Alvarez-Manzaneda, E., & Chahboun, R. (2022). Synthesis and Biological Evaluation of Cassane Diterpene (5α)-Vuacapane-8(14), 9(11)-Diene and of Some Related Compounds. Molecules, 27(17), 5705. https://doi.org/10.3390/molecules27175705