
Antidiarrheal and Antibacterial Activities of Monterey Cypress Phytochemicals: In Vivo and In Vitro Approach

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Metabolite Profiling of the C. macrocarpa Roots Methanol Extract
2.1.1. Flavones, Biflavones and Their Glycosides
2.1.2. Flavonols and Their Glycosides
2.1.3. Flavanones and Their Glycosides
2.1.4. Procyanidins
2.1.5. Stilbenes and Other Compounds
2.2. Spectroscopic Data of Isolated Compounds
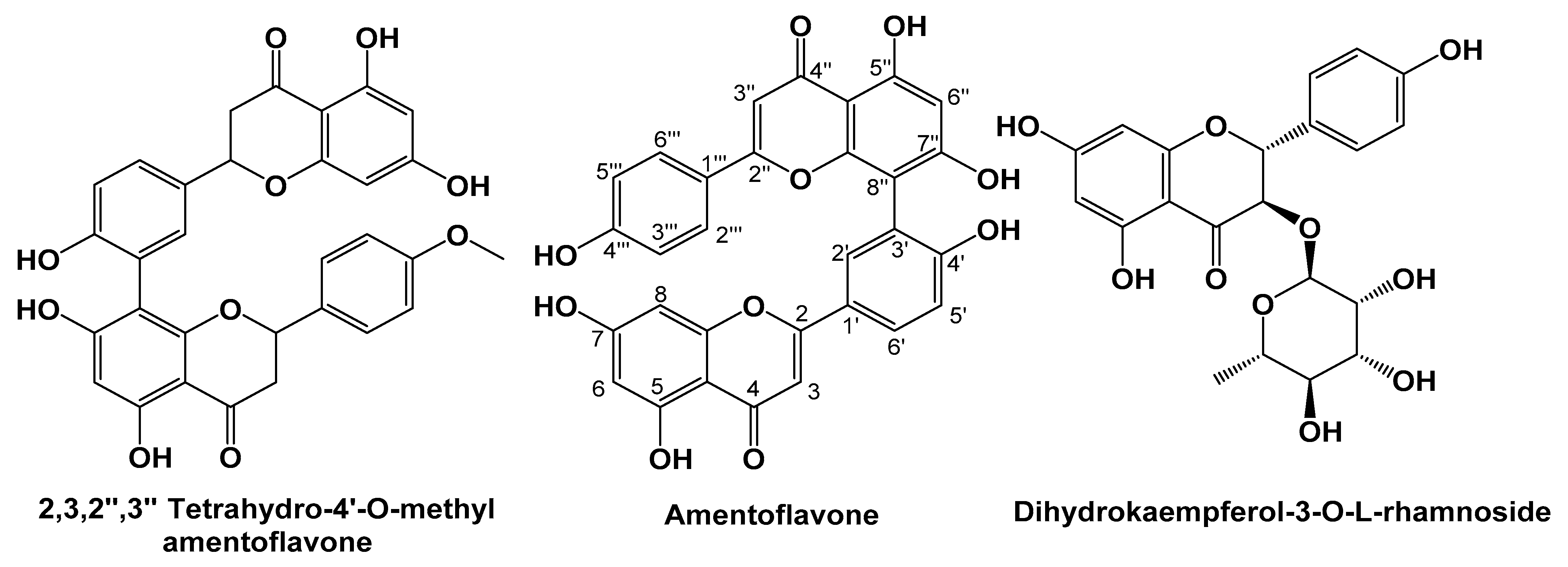
2.2.1. Characterization of 2,3,2″,3″-tetrahydro-4′-O-methyl Amentoflavone
2.2.2. Characterization of Amentoflavone
2.2.3. Characterization of Dihydrokaempferol-3-O-α-l-rhamnoside
2.3. In Vitro Antibacterial Effect of CRME against S. enterica Isolates
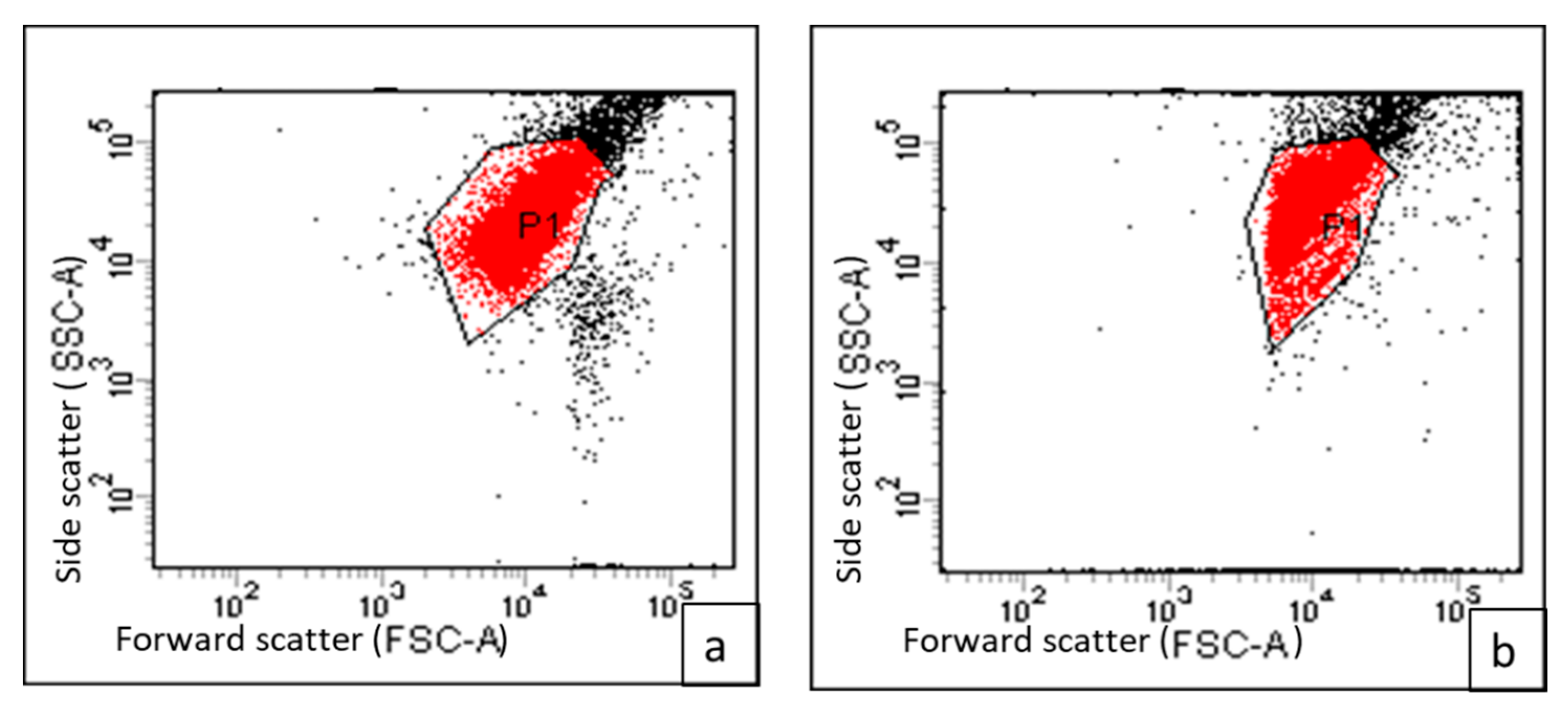
2.3.1. Genotypic Patterns of S. enterica Isolates Using the ERIC-PCR Technique
2.3.2. Antimicrobial Activity of C. macrocarpa Roots
2.3.3. Antimicrobial Activity of Isolated Pure Compounds
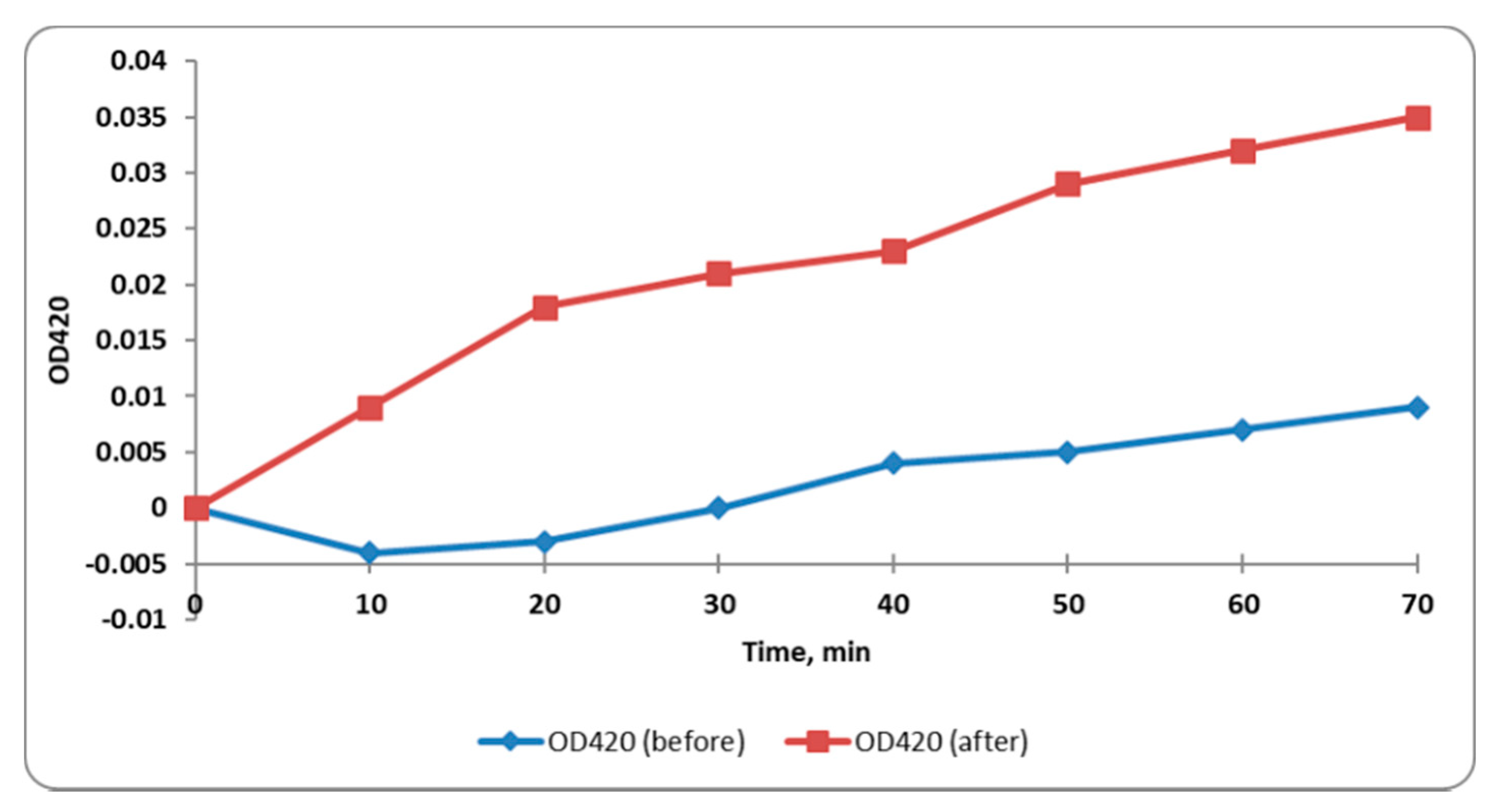
2.3.4. Integrity of Cell Membrane
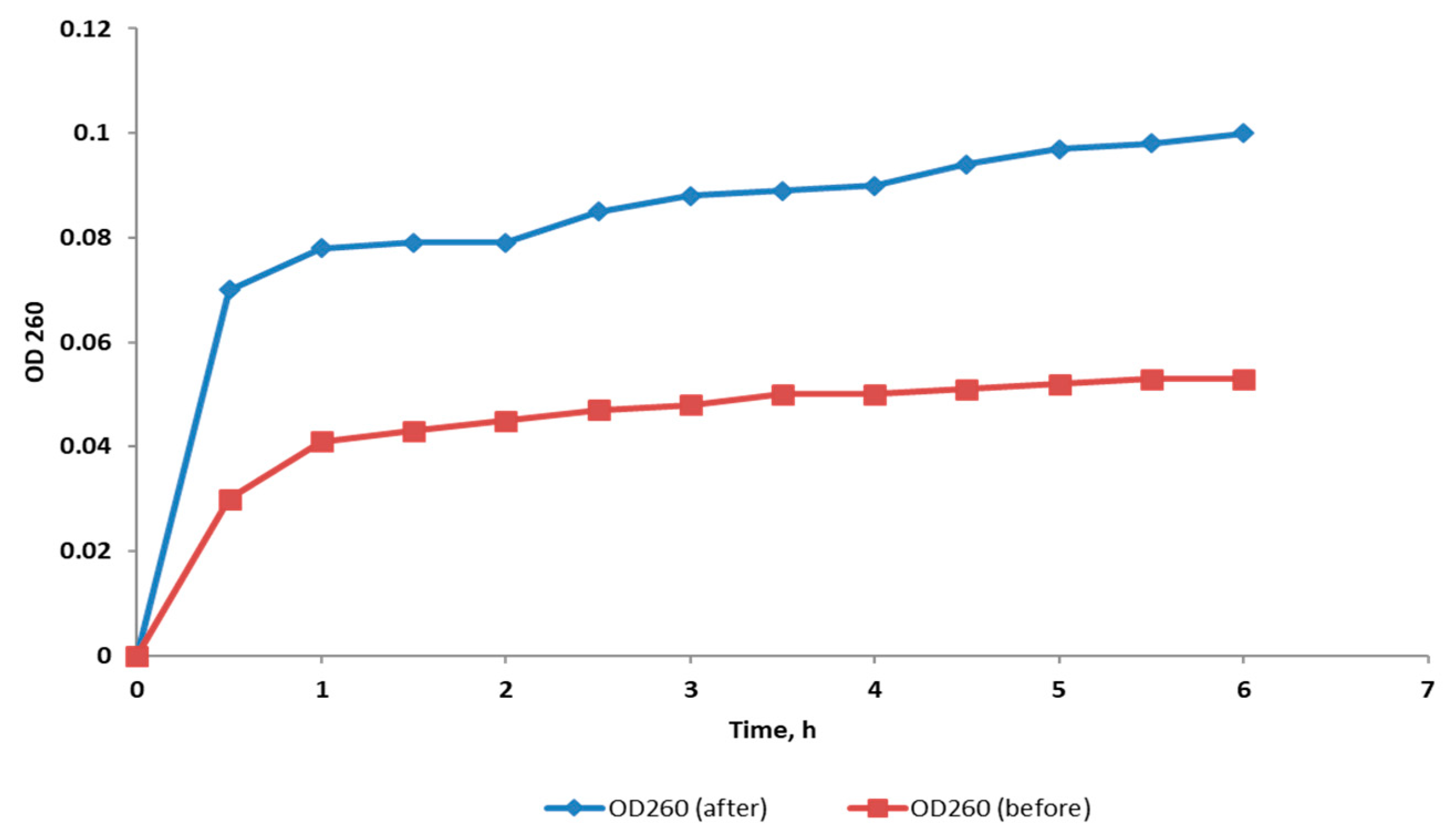
2.3.5. Inner Membrane Permeability
2.3.6. Outer Membrane Permeability
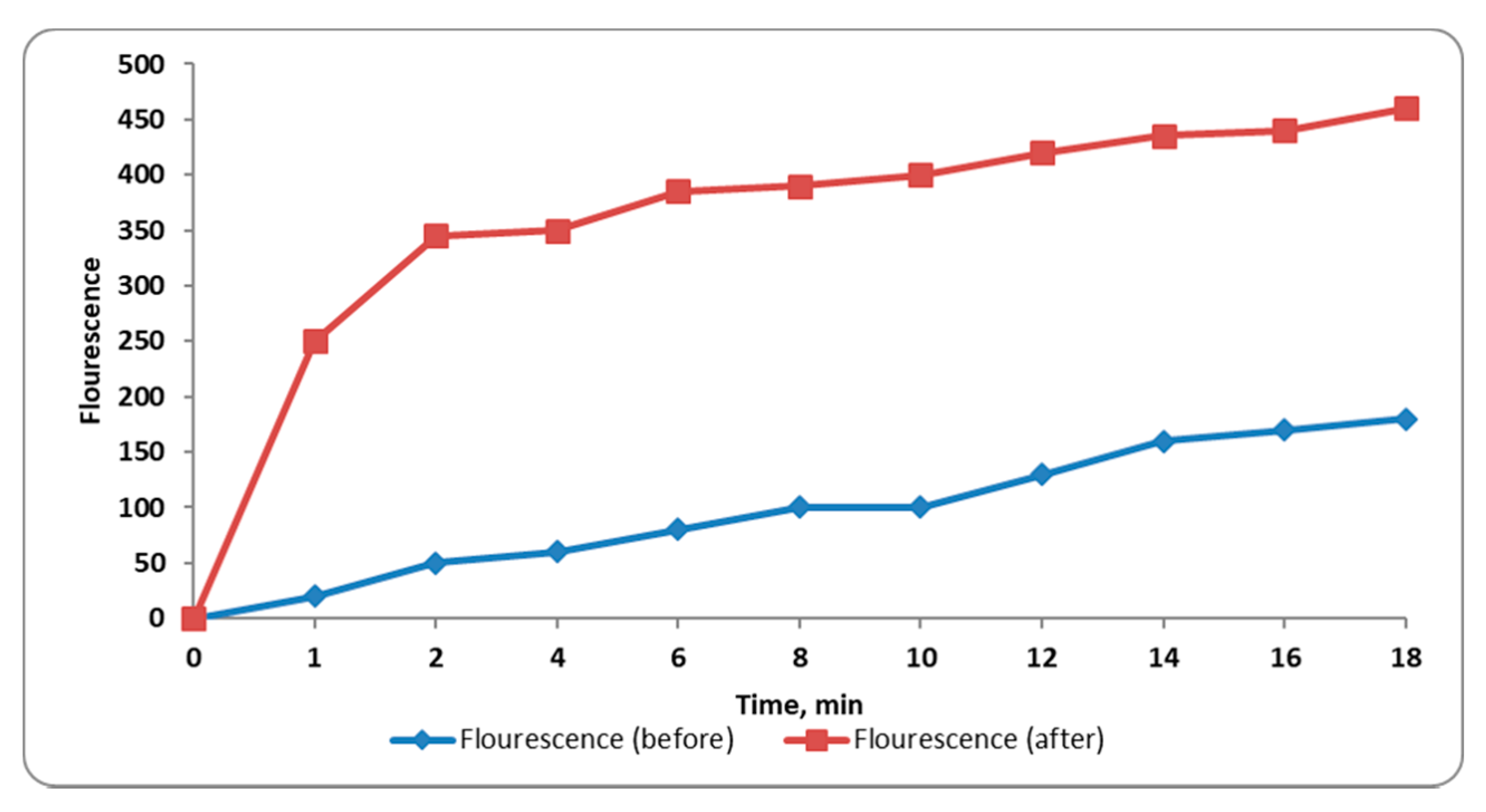
2.3.7. Membrane Depolarization
2.3.8. Effect on Efflux Activity
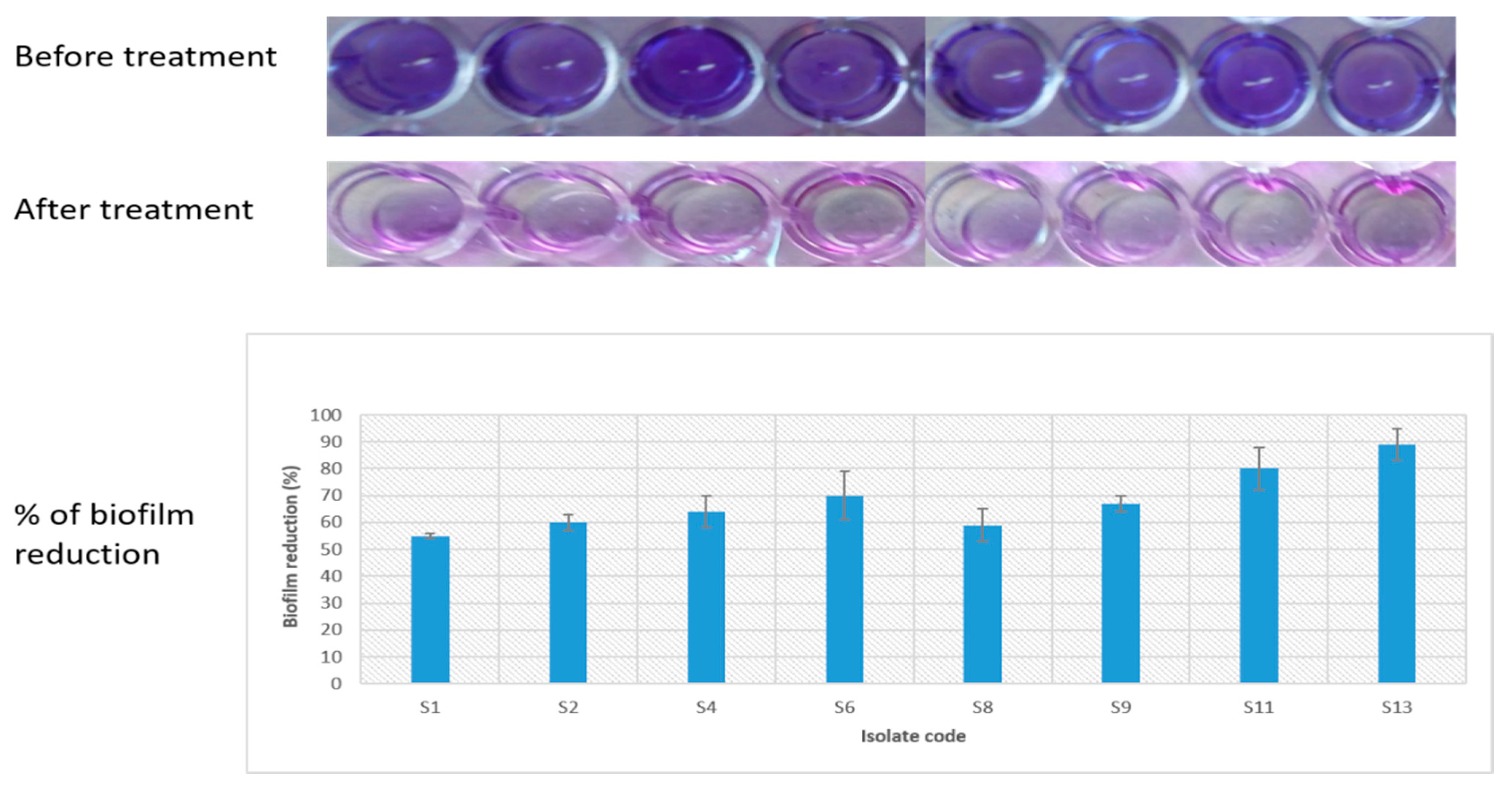
2.3.9. Antibiofilm Activity of C. macrocarpa Roots Methanol Extract
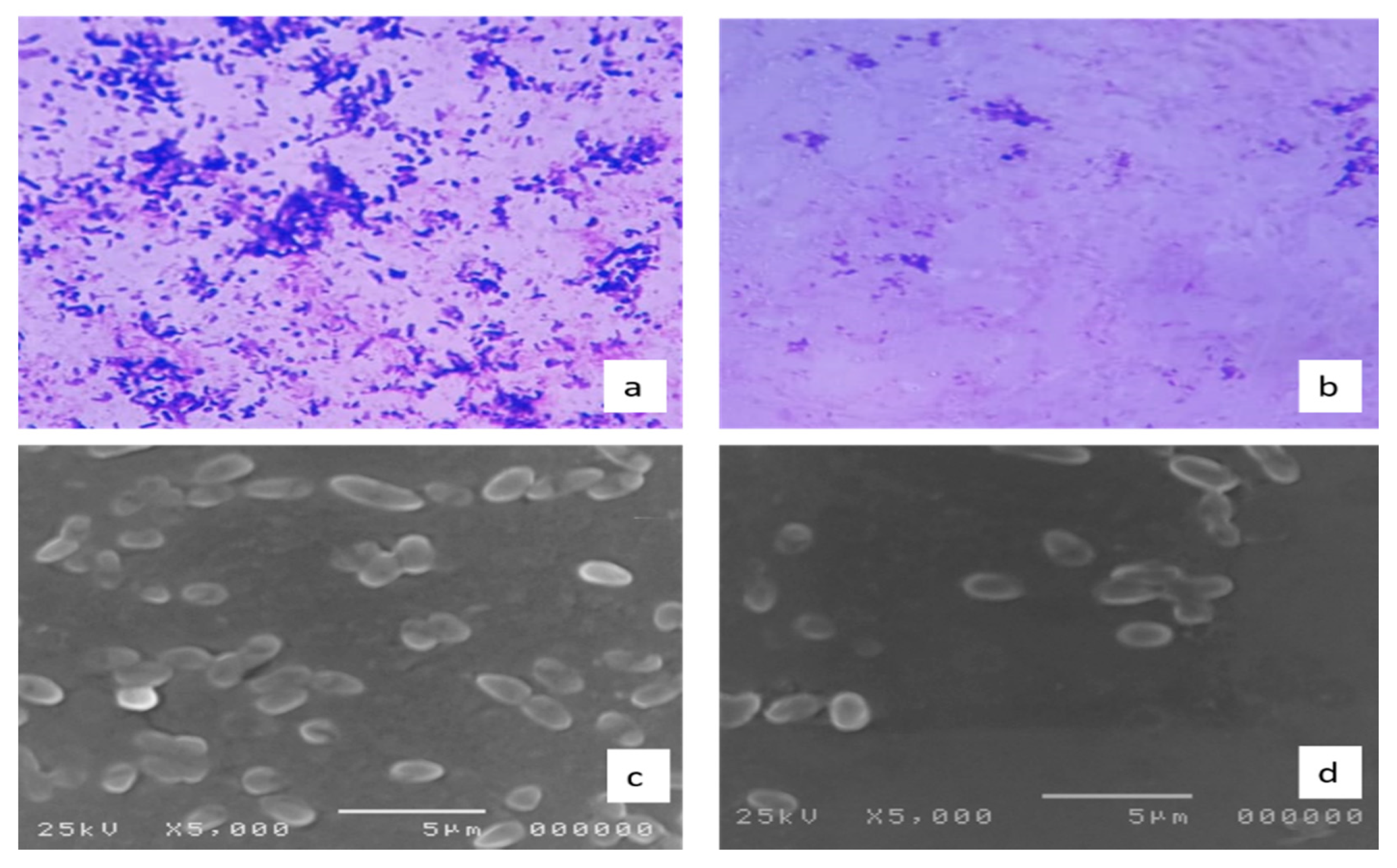
2.3.10. Effect on the Biofilm Morphology
2.3.11. Quantitative Real-Time PCR (qRT-PCR)
2.4. Antidiarrheal Effect of Cupressus Macrocarpa Roots Methanol Extract
2.4.1. Test of Acute Oral Toxicity
2.4.2. Castor Oil-Induced Diarrhea
2.4.3. Impact on Castor Oil-Induced Enteropooling
2.4.4. Gastrointestinal Motility Test
2.4.5. Antidiarrheal Index
3. Discussion
4. Materials and Methods
4.1. General
4.2. Plant Material
4.3. Extraction and Isolation of Different Compounds
4.4. LC-MS/MS Conditions
4.5. Antibacterial Screening
4.5.1. Determination of MICs
4.5.2. Integrity of Cell Membranes
4.5.3. Efflux Assay
4.5.4. Antibiofilm Activity Assay
4.5.5. Impact on Biofilm Morphology by Light Microscope and SEM
4.5.6. qRT-PCR
4.6. In Vivo Antidiarrheal Effect of C. macrocarpa Roots Methanol Extract
4.6.1. Experimental Animals
4.6.2. Experimental Design of the Antidiarrheal Study
4.6.3. Diarrhea Induced with Castor Oil Oral Administration in Mice
4.6.4. Enteropooling Induction by Castor Oil Oral Administration in Mice
4.6.5. Gastrointestinal Motility Test
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sadiq, M.B.; Tarning, J.; Aye Cho, T.Z.; Anal, A.K. Antibacterial activities and possible modes of action of Acacia nilotica (L.) Del. against multidrug-resistant Escherichia coli and Salmonella. Molecules 2017, 22, 47. [Google Scholar] [CrossRef] [PubMed]
- Heredia, N.; García, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef]
- Ćwiek, K.; Korzekwa, K.; Tabiś, A.; Bania, J.; Bugla-Płoskońska, G.; Wieliczko, A. Antimicrobial Resistance and Biofilm Formation Capacity of Salmonella enterica Serovar Enteritidis Strains Isolated from Poultry and Humans in Poland. Pathogens 2020, 9, 643. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Regal, P.; Vázquez, B.; Miranda, J.M.; Cepeda, A.; Franco, C.M. Salmonella and Campylobacter biofilm formation: A comparative assessment from farm to fork. J. Sci. Food Agric. 2018, 98, 4014–4032. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Bazzaz, B.S.F. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control. 2019, 8, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petri, W.A.; Miller, M.; Binder, H.J.; Levine, M.M.; Dillingham, R.; Guerrant, R.L. Enteric infections, diarrhea, and their impact on function and development. J. Clin. Investig. 2008, 118, 1277–1290. [Google Scholar] [CrossRef]
- Rudra, S.; Tahamina, A.; Emon, N.U.; Adnan, M.; Shakil, M.; Chowdhury, M.; Uddin, H.; Barlow, J.W.; Alwahibi, M.S.; Soliman Elshikh, M. Evaluation of various solvent extracts of Tetrastigma leucostaphylum (Dennst.) Alston leaves, a Bangladeshi traditional medicine used for the treatment of Diarrhea. Molecules 2020, 25, 4994. [Google Scholar] [CrossRef]
- Mekonnen, B.; Asrie, A.B.; Wubneh, Z.B. Antidiarrheal activity of 80% methanolic leaf extract of Justicia schimperiana. Evid.-Based Complementary Altern. Med. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.P.; Bartel, J.A.; Price, R.A. A new genus, Hesperocyparis, for the cypresses of the Western Hemisphere (Cupressaceae). Phytologia 2009, 91, 160–185. [Google Scholar]
- Al-Sayed, E.; Abdel-Daim, M. Protective role of Cupressuflavone from Cupressus macrocarpa against carbon tetrachloride-induced hepato-and nephrotoxicity in mice. Planta Med. 2014, 80, 1665–1671. [Google Scholar] [CrossRef] [Green Version]
- Al-Sayed, E.; Gad, H.A.; El-Shazly, M.; Abdel-Daim, M.M.; Nasser Singab, A. Anti-inflammatory and analgesic activities of cupressuflavone from Cupressus macrocarpa: Impact on pro-inflammatory mediators. Drug Dev. Res. 2018, 79, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Saab, A.M.; Tundis, R.; Statti, G.A.; Menichini, F.; Lampronti, I.; Gambari, R.; Cinatl, J.; Doerr, H.W. Phytochemical analysis and in vitro antiviral activities of the essential oils of seven Lebanon species. Chem. Biodivers. 2008, 5, 461–470. [Google Scholar] [CrossRef]
- Negm, W.; Abo El-Seoud, K.; Kabbash, A.; El-Aasr, M. Investigation of the Biological Activity Some Gymnosperm Plants Belong to Cycadales Order. J. Adv. Med. Pharm. Res. 2020, 1, 9–13. [Google Scholar] [CrossRef]
- Al-Sayed, E.; Ke, T.-Y.; Hwang, T.-L.; Chen, S.-R.; Korinek, M.; Chen, S.-L.; Cheng, Y.-B. Cytotoxic and anti-inflammatory effects of lignans and diterpenes from Cupressus macrocarpa. Bioorg. Med. Chem. Lett. 2020, 30, 127127. [Google Scholar] [CrossRef]
- Briggs, L.H.; Sutherland, M.D. The essential oil of Cupressus macrocarpa. J. Org. Chem. 1942, 7, 397–407. [Google Scholar] [CrossRef]
- Cool, L.G. Sesquiterpenes from Cupressus macrocarpa foliage. Phytochemistry 2005, 66, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Attallah, N.G.M.; Negm, W.A.; Elekhnawy, E.; Elmongy, E.I.; Altwaijry, N.; El-Haroun, H.; El-Masry, T.A.; El-Sherbeni, S.A. Elucidation of Phytochemical Content of Cupressus macrocarpa Leaves: In Vitro and In Vivo Antibacterial Effect against Methicillin-Resistant Staphylococcus aureus Clinical Isolates. Antibiotics 2021, 10, 890. [Google Scholar] [CrossRef]
- Moawad, A.; Hetta, M.; Zjawiony, J.K.; Jacob, M.R.; Hifnawy, M.; Ferreira, D. Phytochemical investigation of Cycas circinalis and Cycas revoluta leaflets: Moderately active antibacterial biflavonoids. Planta Med. 2010, 76, 796. [Google Scholar] [CrossRef] [Green Version]
- Negm, W.A.; Ibrahim, A.E.-R.S.; El-Seoud, K.A.; Attia, G.I.; Ragab, A.E. A new cytotoxic and antioxidant Amentoflavone Monoglucoside from Cycas revoluta Thunb growing in Egypt. J. Pharm. Sci. Res. 2016, 8, 343. [Google Scholar]
- Negm, W.A.; Abo El-Seoud, K.A.; Kabbash, A.; Kassab, A.A.; El-Aasr, M. Hepatoprotective, cytotoxic, antimicrobial and antioxidant activities of Dioon spinulosum leaves Dyer Ex Eichler and its isolated secondary metabolites. Nat. Prod. Res. 2020, 35, 5166–5176. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A review on flavonoid apigenin: Dietary intake, ADME, antimicrobial effects, and interactions with human gut microbiota. BioMed Res. Int. 2019, 2019. [Google Scholar] [CrossRef]
- Nitiema, L.W.; Savadogo, A.; Simpore, J.; Dianou, D.; Traore, A.S. In vitro antimicrobial activity of some phenolic compounds (coumarin and quercetin) against gastroenteritis bacterial strains. Int. J. Microbiol. Res. 2012, 3, 183–187. [Google Scholar]
- Sholkamy, E.N.; Ahmed, M.S.; Yasser, M.M.; Mostafa, A.A. Antimicrobial quercetin 3-O-glucoside derivative isolated from Streptomyces antibioticus strain ess_amA8. J. King Saud Univ.-Sci. 2020, 32, 1838–1844. [Google Scholar] [CrossRef]
- Chiruvella, K.K.; Mohammed, A.; Dampuri, G.; Ghanta, R.G.; Raghavan, S.C. Phytochemical and antimicrobial studies of methyl angolensate and luteolin-7-O-glucoside isolated from callus cultures of Soymida febrifuga. Int. J. Biomed. Sci. 2007, 3, 269. [Google Scholar]
- Qian, W.; Liu, M.; Fu, Y.; Zhang, J.; Liu, W.; Li, J.; Li, X.; Li, Y.; Wang, T. Antimicrobial mechanism of luteolin against Staphylococcus aureus and Listeria monocytogenes and its antibiofilm properties. Microb. Pathog. 2020, 142, 104056. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Bao, Y.; Zhu, Y.; Liu, S.; Peng, Z.; Zhang, Y.; Zhou, G. Isorhamnetin, hispidulin, and cirsimaritin identified in Tamarix ramosissima barks from southern Xinjiang and their antioxidant and antimicrobial activities. Molecules 2019, 24, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, S.-H.; Lee, K.-A.; Park, K.-K.; Kim, K.-T.; Park, Y.-S.; Nah, S.-Y.; Mendonca, A.F.; Paik, H.-D. Antimicrobial effects of natural flavonoids and a novel flavonoid, 7-O-Butyl Naringenin, on growth of meat-borne Staphylococcus aureus strains. Food Sci. Anim. Resour. 2011, 31, 413–419. [Google Scholar] [CrossRef]
- Sati, P.; Dhyani, P.; Bhatt, I.D.; Pandey, A. Ginkgo biloba flavonoid glycosides in antimicrobial perspective with reference to extraction method. J. Tradit. Complement. Med. 2019, 9, 15–23. [Google Scholar] [CrossRef]
- Mandalari, G.; Bennett, R.; Bisignano, G.; Trombetta, D.; Saija, A.; Faulds, C.; Gasson, M.; Narbad, A. Antimicrobial activity of flavonoids extracted from bergamot (Citrus bergamia Risso) peel, a byproduct of the essential oil industry. J. Appl. Microbiol. 2007, 103, 2056–2064. [Google Scholar] [CrossRef] [PubMed]
- Alejo-Armijo, A.; Glibota, N.; Frías, M.P.; Altarejos, J.; Gálvez, A.; Ortega-Morente, E.; Salido, S. Antimicrobial and antibiofilm activities of procyanidins extracted from laurel wood against a selection of foodborne microorganisms. Int. J. Food Sci. Technol. 2017, 52, 679–686. [Google Scholar] [CrossRef]
- Chibueze, I.P.; Earnest, E.O.; Sonne, M.I.; Emmanuel, I.; Emeka, A.L.D.; Basden, O.F. Antidiarrheal property of Napoleona imperialis may be due to Procyanidins and Ellagic acid derivatives. J. Appl. Pharm. Sci. 2016, 6, 101–106. [Google Scholar]
- Cruz, L.F.; de Figueiredo, G.F.; Pedro, L.P.; Amorin, Y.M.; Andrade, J.T.; Passos, T.F.; Rodrigues, F.F.; Souza, I.L.A.; Gonçalves, T.P.R.; dos Santos Lima, L.A.R. Umbelliferone (7-hydroxycoumarin): A non-toxic antidiarrheal and antiulcerogenic coumarin. Biomed. Pharmacother. 2020, 129, 110432. [Google Scholar] [CrossRef] [PubMed]
- Jalilzadeh-Amin, G.; Maham, M. The application of 1, 8-cineole, a terpenoid oxide present in medicinal plants, inhibits castor oil-induced diarrhea in rats. Pharm. Biol. 2015, 53, 594–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, S.A.; Martins, C.; Pereira, C.; Silvestre, A.J.; Rocha, S.M. Current challenges and perspectives for the use of aqueous plant extracts in the management of bacterial infections: The case-study of Salmonella enterica serovars. Int. J. Mol. Sci. 2019, 20, 940. [Google Scholar] [CrossRef] [Green Version]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 980–987. [Google Scholar] [CrossRef]
- Chen, C.Z.; Cooper, S.L. Interactions between dendrimer biocides and bacterial membranes. Biomaterials 2002, 23, 3359–3368. [Google Scholar] [CrossRef]
- Heesterbeek, D.A.; Muts, R.M.; van Hensbergen, V.P.; de Saint Aulaire, P.; Wennekes, T.; Bardoel, B.W.; van Sorge, N.M.; Rooijakkers, S.H. Outer membrane permeabilization by the membrane attack complex sensitizes Gram-negative bacteria to antimicrobial proteins in serum and phagocytes. PLoS Pathog. 2021, 17, e1009227. [Google Scholar] [CrossRef]
- Bouyahya, A.; Abrini, J.; Dakka, N.; Bakri, Y. Essential oils of Origanum compactum increase membrane permeability, disturb cell membrane integrity, and suppress quorum-sensing phenotype in bacteria. J. Pharm. Anal. 2019, 9, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Te Winkel, J.D.; Gray, D.A.; Seistrup, K.H.; Hamoen, L.W.; Strahl, H. Analysis of antimicrobial-triggered membrane depolarization using voltage sensitive dyes. Front. Cell Dev. Biol. 2016, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Anbazhagan, P.V.; Thavitiki, P.R.; Varra, M.; Annamalai, L.; Putturu, R.; Lakkineni, V.R.; Pesingi, P.K. Evaluation of efflux pump activity of multidrug-resistant Salmonella Typhimurium isolated from poultry wet markets in India. Infect. Drug Resist. 2019, 12, 1081. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med. Res. 2019, 149, 129. [Google Scholar]
- Wi, Y.M.; Patel, R. Understanding biofilms and novel approaches to the diagnosis, prevention, and treatment of medical device-associated infections. Infect. Dis. Clin. 2018, 32, 915–929. [Google Scholar] [CrossRef]
- Gomes, F.; Martins, N.; Ferreira, I.C.; Henriques, M. Anti-biofilm activity of hydromethanolic plant extracts against Staphylococcus aureus isolates from bovine mastitis. Heliyon 2019, 5, e01728. [Google Scholar] [CrossRef] [Green Version]
- Famuyide, I.M.; Aro, A.O.; Fasina, F.O.; Eloff, J.N.; McGaw, L.J. Antibacterial and antibiofilm activity of acetone leaf extracts of nine under-investigated south African Eugenia and Syzygium (Myrtaceae) species and their selectivity indices. BMC Complement. Altern. Med. 2019, 19, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Alam, K.; Al Farraj, D.A.; Mah-E-Fatima, S.; Yameen, M.A.; Elshikh, M.S.; Alkufeidy, R.M.; Mustafa, A.E.-Z.M.; Bhasme, P.; Alshammari, M.K.; Alkubaisi, N.A. Anti-biofilm activity of plant derived extracts against infectious pathogen-Pseudomonas aeruginosa PAO1. J. Infect. Public Health 2020, 13, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.M.; Vivacqua, C.G.; Abdulhamid, M.B.; Baigori, M.D.; Slanis, A.C.; Allori, M.C.G.D.; Tereschuk, M.L. Biofilm inhibition activity of traditional medicinal plants from Northwestern Argentina against native pathogen and environmental microorganisms. Rev. Soc. Bras. Med. Trop. 2016, 49, 703–712. [Google Scholar] [CrossRef]
- Omwenga, E.O.; Hensel, A.; Pereira, S.; Shitandi, A.A.; Goycoolea, F.M. Antiquorum sensing, antibiofilm formation and cytotoxicity activity of commonly used medicinal plants by inhabitants of Borabu sub-county, Nyamira County, Kenya. PLoS ONE 2017, 12, e0185722. [Google Scholar] [CrossRef] [Green Version]
- Gidudu, J.; Sack, D.; Pina, M.; Hudson, M.; Kohl, K.; Bishop, P.; Chatterjee, A.; Chiappini, E.; Compingbutra, A.; Da Costa, C. Diarrhea: Case definition and guidelines for collection, analysis, and presentation of immunization safety data. Vaccine 2011, 29, 1053. [Google Scholar] [CrossRef] [PubMed]
- Famuyide, I.M.; Aro, A.O.; Fasina, F.O.; Eloff, J.N.; McGaw, L.J. Antibacterial activity and mode of action of acetone crude leaf extracts of under-investigated Syzygium and Eugenia (Myrtaceae) species on multidrug resistant porcine diarrhoeagenic Escherichia coli. BMC Vet. Res. 2019, 15, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayede, D.; Mulaw, T.; Kahaliw, W. Antidiarrheal activity of hydromethanolic root extract and solvent fractions of clutia abyssinica jaub. & spach.(euphorbiaceae) in mice. Evid.-Based Complementary Altern. Med. 2020, 2020, 5416749. [Google Scholar]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Das, D.C.; De, S.; Bhattacharya, S.; Das, M. Antibacterial activity and Phytochemical analysis of Cardanthera difformis Druce leaf extracts from West Bengal, India. Int. J. Phytomedicine 2013, 5, 446. [Google Scholar]
- Wayne, A. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing. CLSI Doc. 2017, 31, 100–121. [Google Scholar]
- Elekhnawy, E.; Sonbol, F.; Abdelaziz, A.; Elbanna, T. An investigation of the impact of triclosan adaptation on Proteus mirabilis clinical isolates from an Egyptian university hospital. Braz. J. Microbiol. 2021, 52, 927–937. [Google Scholar] [CrossRef]
- Sonbol, F.I.; El-Banna, T.; Abd El-Aziz, A.A.; El-Ekhnawy, E. Impact of triclosan adaptation on membrane properties, efflux and antimicrobial resistance of Escherichia coli clinical isolates. J. Appl. Microbiol. 2019, 126, 730–739. [Google Scholar] [CrossRef]
- Abdelaziz, A.; Sonbol, F.; Elbanna, T.; El-Ekhnawy, E. Exposure to sublethal concentrations of benzalkonium chloride induces antimicrobial resistance and cellular changes in Klebsiellae pneumoniae clinical isolates. Microb. Drug Resist. 2019, 25, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Farrag, H.A.; Hosny, A.E.-D.M.; Hawas, A.M.; Hagras, S.A.; Helmy, O.M. Potential efficacy of garlic lock therapy in combating biofilm and catheter-associated infections; experimental studies on an animal model with focus on toxicological aspects. Saudi Pharm. J. 2019, 27, 830–840. [Google Scholar] [CrossRef]
- Bakkiyaraj, D.; Pandian, S.T.K. In vitro and in vivo antibiofilm activity of a coral associated actinomycete against drug resistant Staphylococcus aureus biofilms. Biofouling 2010, 26, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; She, P.; Wang, Y.; Liu, F.; Zhang, D.; Chen, L.; Luo, Z.; Xu, H.; Qi, Y.; Wu, Y. Effects of norspermidine on Pseudomonas aeruginosa biofilm formation and eradication. Microbiologyopen 2016, 5, 402–412. [Google Scholar] [CrossRef] [Green Version]
- Attallah, N.G.; Negm, W.A.; Elekhnawy, E.; Altwaijry, N.; Elmongy, E.I.; El-Masry, T.A.; Alturki, E.A.; Yousef, D.A.; Shoukheba, M.Y. Antibacterial Activity of Boswellia sacra Flueck. Oleoresin Extract against Porphyromonas gingivalis Periodontal Pathogen. Antibiotics 2021, 10, 859. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, D.; Zhang, Q.; Liao, X.-P.; Liu, Y.-H.; Sun, J. Efflux pump overexpression contributes to tigecycline heteroresistance in Salmonella enterica serovar Typhimurium. Front. Cell. Infect. Microbiol. 2017, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Borges, K.A.; Furian, T.Q.; de Souza, S.N.; Menezes, R.; de Lima, D.A.; Fortes, F.B.B.; Salle, C.T.P.; Moraes, H.L.S.; Nascimento, V.P. Biofilm formation by Salmonella Enteritidis and Salmonella Typhimurium isolated from avian sources is partially related with their in vivo pathogenicity. Microb. Pathog. 2018, 118, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Khosravani, M.; Dallal, M.M.S.; Norouzi, M. Phytochemical Composition and Anti-Efflux Pump Activity of Hydroalcoholic, Aqueous, and Hexane Extracts of Artemisia tournefortiana in Ciprofloxacin-Resistant Strains of Salmonella enterica Serotype Enteritidis. Iran. J. Public Health 2020, 49, 134. [Google Scholar] [CrossRef]
- Zheng, J.; Tian, F.; Cui, S.; Song, J.; Zhao, S.; Brown, E.W.; Meng, J. Differential gene expression by RamA in ciprofloxacin-resistant Salmonella Typhimurium. PLoS ONE 2011, 6, e22161. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory; National Academy Press: Washington, DC, USA, 2011; ISBN 9780309154000. [Google Scholar]
- Carlo, G.D.; Mascolo, N.; Izzo, A.A.; Capasso, F.; Autore, G. Effects of quercetin on the gastrointestinal tract in rats and mice. Phytother. Res. 1994, 8, 42–45. [Google Scholar] [CrossRef]
- MacFaddin, J. Biochemical Tests for Identification of Medical Bacteria; Williams and Wilkins: Philadelphia, PA, USA, 2000. [Google Scholar]
- Kim, J.E.; Lee, Y.J. Molecular characterization of antimicrobial resistant non-typhoidal Salmonella from poultry industries in Korea. Ir. Vet. J. 2017, 70, 1–9. [Google Scholar] [CrossRef]
- Organization for Economic Cooperation and Development. Test No. 425: Acute Oral Toxicity: Up-and-Down Procedure. In OECD Guidelines for the Testing of Chemicals, Section 4; OECD Publishing: Paris, France, 2008. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Assignment | RT (min.) | (M–H)− m/z | Formulas | Fragments or MS2 m/z |
|---|---|---|---|---|---|
| 1 | D-(-)-Quinic acid | 1.234 | 191.056 | C7H12O6 | 127.039, 171.030, 191.053 |
| 2 | (-)-Shikimic acid | 1.235 | 173.0456 | C7H10O5 | 129.053, 137.021, 155.039, 173.043 |
| 3 | Maleic acid | 1.237 | 115.0018 | C4H4O4 | 71.018, 114.999 |
| 4 | Citraconic acid | 1.299 | 129.0532 | C5H6O4 | 29.050, 85.001 |
| 5 | Procyanidin B2 | 4.592 | 577.1344 | C30H26O12 | 289.021, 425.126, 577.134 |
| 6 | Urocanic acid | 4.745 | 137.0244 | C6H6N2O2 | 137.024 |
| 7 | (-)-Epicatechin | 4.865 | 289.0729 | C15H14O6 | 245.080, 289.072 |
| 8 | Neohesperidin dihydrochalcone | 5.273 | 611.1586 | C28H36O15 | 543.161, 548.867, 611.165 |
| 9 | Dihydrokaempferol-3-O-α-l-rhamnoside | 5.355 | 433.1909 | C21H22O10 | 179.059, 288.312, 342.957, 434.189 |
| 10 | Apigenin-7-O-neohesperidoside (Rhoifolin) | 5.677 | 577.1384 | C27H30O14 | 269.043, 425.081, 532.910, 577.118 |
| 11 | Procyanidin C1 | 5.677 | 865.1961 | C45H38O18 | 289.201, 465.301, 865.194 |
| 12 | Procyanidin B1 | 6.034 | 577.1349 | C30H26O12 | 289.231, 425.102, 577.125 |
| 13 | Naringenin-7-O-glucoside (Prunin) | 6.209 | 433.1129 | C21H22O10 | 271.065, 433.102, 433.196 |
| 14 | Amentoflavone | 6.221 | 537.194 | C30H18O10 | 375.152, 399.210, 443.012, 537.189 |
| 15 | E-3,4,5′-Trihydroxy-3′ glucopyranosylstilbene (Astringin) | 6.482 | 405.1201 | C20H22O9 | 243.067, 405.124 |
| 16 | Isorhamnetin-3-O-rutinoside (Narcissin) | 6.894 | 623.1995 | C28H32O16 | 315. 211, 577.199, 623.171, 623.192 |
| 17 | Luteolin-7-O-glucoside | 6.982 | 447.0892 | C21H20O11 | 285.035, 402.878, 447.084 |
| 18 | Quercetin-3-D-xyloside | 7.060 | 433.1666 | C20H18O11 | 301.028, 326.927, 364.901, 433.164 |
| 19 | 5-Methoxysalicylic acid | 7.061 | 167.0348 | C8H8O4 | 152.010, 167.034 |
| 20 | Isorhamnetin-3-O-glucoside | 7.234 | 477.1055 | C22H22O12 | 315.325, 454.144, 477.109 |
| 21 | Ferulic acid | 7.402 | 193.0874 | C10H10O4 | 178.066, 193.085 |
| 22 | Tetrahydro-4′-O-methyl amentoflavone | 7.668 | 555.440 | C31H24O10 | 541.022, 555.401 |
| 23 | Apigenin-7-O-glucoside | 7.805 | 431.0969 | C21H20O10 | 269.042, 430.886, 431.103 |
| 24 | Acacetin-7-O-rutinoside | 7.855 | 591.1512 | C28H32O14 | 283.0124, 392.898, 528.866, 591.144 |
| 25 | Baicalein-7-O-glucuronide | 7.994 | 445.1177 | C21H18O11 | 112.987, 163.079, 269.353, 445.109 |
| 26 | Kaempferol-3-glucuronide | 8.006 | 461.1081 | C21H18O12 | 285.331, 324.911, 392.897, 461.103 |
| 27 | Quercetin | 9.627 | 301.0349 | C15H10O7 | 255.101, 301.031 |
| 28 | Naringenin | 10.183 | 271.0601 | C15H12O5 | 93.040, 151.007, 271.057 |
| 29 | Sinapyl aldehyde | 11.077 | 207.0636 | C11H12O4 | 192.0412, 207.065 |
| 30 | 3,3′,4′,5,7-pentahydroxyflavan | 12.067 | 289.1812 | C15H14O6 | 271.159, 289.180 |
| 31 | 3′-methoxy-4′,5,7-trihydroxyflavonol (isorhamnetin) | 13.426 | 315.1947 | C16H12O7 | 227.109, 283.168, 315.195 |
| 32 | Hesperetin | 16.591 | 301.1769 | C16H14O6 | 301.178 |
| 33 | Apigenin | 16.725 | 269.1541 | C15H10O5 | 269.157 |
| 34 | Acacetin | 18.035 | 283.1747 | C16H12O5 | 268.153, 283.162 |
| 35 | Esculin | 18.257 | 339.1972 | C15H16O9 | 295.205, 303.902, 339.191 |
| 36 | Luteolin | 18.330 | 285.1844 | C15H10O6 | 269.158, 285.183 |
| 37 | 3,5,7-trihydroxy-4′-methoxyflavone (diosmetin) | 20.352 | 299.2003 | C16H12O6 | 231.101, 283.155, 299.199 |
| 38 | Kaempferol-7-neohesperidoside | 21.203 | 593.1549 | C27H30O15 | 285.092, 389.172, 547.337, 593.131 |
| 39 | Glycyrrhizate (glycyrrhizin) | 26.874 | 821.3727 | C42H62O16 | 685.426, 775.427, 821.3 |
| Before Treatment | After Treatment | ||
|---|---|---|---|
| Minimum Conc. of EtBr (mg/L) (Number of Isolates) | Efflux Activity * | Minimum Conc. of EtBr (mg/L) (Number of Isolates) | Efflux Activity * |
| ≤0.5 (3) | -(N) | ≤0.5 (6) | -(N) |
| 1 (1) | +(I) | 1 (9) | +(I) |
| 1.5 (1) | +(I) | 1.5 (2) | +(I) |
| 2 (2) | +(I) | 2 (1) | +(I) |
| 2.5 (13) | ++(P) | 2 (2) | ++(P) |
| Isolate Code | Relative Gene Expression * | |||
|---|---|---|---|---|
| acrA | acrB | tolC | oqxB | |
| S1 | 0.1 ± 0.3 | 0.2 ± 0.0 | 1.1 ± 0.3 | 1.3 ± 0.2 |
| S2 | 1.3 ± 0.1 | 0.3 ± 0.1 | 1.5 ± 0.3 | 1.0 ± 0.7 |
| S3 | 1.4 ± 0.0 | 0.2 ± 0.1 | 1.7 ± 0.1 | 1.8 ± 0.2 |
| S5 | 0.1 ± 0.0 | 1.1 ± 0.2 | 1.0 ± 0.4 | 1.1 ± 0.2 |
| S6 | 1.3 ± 0.2 | 0.2 ± 0.1 | 1.2 ± 0.8 | 1.6 ± 0.2 |
| S7 | 0.5 ± 0.1 | 1.0 ± 2.0 | 1.7 ± 0.2 | 1.5 ± 0.4 |
| S8 | 0.4 ± 0.0 | 0.1 ± 0.1 | 1.4 ± 0.9 | 1.4 ± 0.3 |
| S9 | 0.2 ± 0.1 | 0.5 ± 0.1 | 1.3 ± 0.0 | 0.9 ± 0.2 |
| S15 | 0.9 ± 0.1 | 0.2 ± 0.0 | 1.2 ± 0.0 | 1.6 ± 0.2 |
| S19 | 0.4 ± 0.2 | 0.3 ± 0.2 | 1.6 ± 0.7 | 1.7 ± 0.2 |
| S20 | 0.3 ± 0.0 | 1.6 ± 0.2 | 1.3 ± 0.1 | 1.2 ± 0.0 |
| Isolate Code | Relative Gene Expression | Isolate Code |
|---|---|---|
| agfA | spiA | |
| S1 | 0.4 ± 0.3 | 0.4 ± 0.2 |
| S2 | 0.3 ± 0.1 | 0.1 ± 0.2 |
| S4 | 1.5 ± 0.3 | 0.3 ± 0.2 |
| S6 | 0.3 ± 0.2 | 1.2 ± 0.1 |
| S8 | 0.4 ± 0.0 | 0.1 ± 0.1 |
| S9 | 0.9 ± 0.2 | 0.2 ± 0.1 |
| S11 | 0.2 ± 0.1 | 1.8 ± 0.1 |
| S13 | 0.2 ± 0.0 | 0.1 ± 0.0 |
| Treatment | The Onset of Diarrhea (min) | No. of Dry Feces | No. of Wet Feces | Weight of Dry Feces (g) | Weight of Wet Feces (g) | % Defecation Inhibition |
|---|---|---|---|---|---|---|
| Control | 18.17 ± 1.5 | 11 ± 1.6 | 7.3 ± 1.2 | 1.8 ± 0.2 | 1.6 ± 0.0 | - |
| Loperamide | 100.33 ± 1.2 | 4 ± 0.8 | 1.7 ± 0.5 | 0.19 ± 0.0 | 0.1 ± 0.0 | 91.5 |
| CRME (100 mg/kg) | 68.5 ± 2.4 | 7 ± 0.8 | 3.7 ± 0.9 | 0.29 ± 0.0 | 0.19 ± 0.0 | 85.9 |
| CRME (200 mg/kg) | 87.6 ± 3.0 | 3.4 ± 0.5 | 2.4 ± 0.5 | 0.27 ± 0.0 | 0.14 ± 0.0 | 88 |
| CRME (400 mg/kg) | 108.5 ± 2.2 | 2.7 ± 0.5 | 1.7 ± 0.5 | 0.13 ± 0.0 | 0.09 ± 0.0 | 93.5 |
| Treatment | MVSIC * (mL) | % Inhibition in MVSIC | MWSIC ** (g) | % Inhibition in MWSIC |
|---|---|---|---|---|
| Control | 0.71 ± 0.00 | - | 0.78 ± 0.00 | - |
| Loperamide | 0.23 ± 0.02 | 67.6 | 0.18 ± 0.01 | 77 |
| CRME (100 mg/kg) | 0.68 ± 0.00 | 4.23 | 0.71 ± 0.00 | 8.97 |
| CRME (200 mg/kg) | 0.61 ± 0.00 | 14.08 | 0.7± | 10.26 |
| CRME (400 mg/kg) | 0.18±0.00 | 74.6 | 0.17± | 78.2 |
| Treatment | Distance Traveled by Charcoal (cm) | Length of the Small Intestine (cm) | Peristalsis Index | % of Inhibition |
|---|---|---|---|---|
| Control | 41 ± 0.07 | 50.2 ± 0.17 | 81.67 | - |
| Loperamide | 10.4 ± 0.22 | 51.3 ± 0.25 | 20.27 | 74.63 |
| CRME (100 mg/kg) | 37.37 ± 0.12 | 53.3 ± 0.21 | 70.11 | 8.85 |
| CRME (200 mg/kg) | 14.37 ± 0.26 | 52.4 ± 0.33 | 27.42 | 64.95 |
| CRME (400 mg/kg) | 8.1 ± 0.08 | 53.4 ± 0.22 | 15.16 | 80.24 |
| Treatment | Delay in Defecation Time (Dfreq) | Gut Meal Travel Reduction (Gmeq) | Purging Frequency (Pfreq) | Antidiarrheal Index ADI |
|---|---|---|---|---|
| Control | - | - | - | - |
| Loperamide | 452.17 | 74.63 | 91.5 | 145.62 |
| CRME (100 mg/kg) | 276.99 | 8.85 | 85.9 | 59.5 |
| CRME (200 mg/kg) | 382.11 | 64.95 | 88 | 129.74 |
| CRME (400 mg/kg) | 497.14 | 80.24 | 93.5 | 155.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmongy, E.I.; Negm, W.A.; Elekhnawy, E.; El-Masry, T.A.; Attallah, N.G.M.; Altwaijry, N.; Batiha, G.E.-S.; El-Sherbeni, S.A. Antidiarrheal and Antibacterial Activities of Monterey Cypress Phytochemicals: In Vivo and In Vitro Approach. Molecules 2022, 27, 346. https://doi.org/10.3390/molecules27020346
Elmongy EI, Negm WA, Elekhnawy E, El-Masry TA, Attallah NGM, Altwaijry N, Batiha GE-S, El-Sherbeni SA. Antidiarrheal and Antibacterial Activities of Monterey Cypress Phytochemicals: In Vivo and In Vitro Approach. Molecules. 2022; 27(2):346. https://doi.org/10.3390/molecules27020346
Chicago/Turabian StyleElmongy, Elshaymaa I., Walaa A. Negm, Engy Elekhnawy, Thanaa A. El-Masry, Nashwah G. M. Attallah, Najla Altwaijry, Gaber El-Saber Batiha, and Suzy A. El-Sherbeni. 2022. "Antidiarrheal and Antibacterial Activities of Monterey Cypress Phytochemicals: In Vivo and In Vitro Approach" Molecules 27, no. 2: 346. https://doi.org/10.3390/molecules27020346
APA StyleElmongy, E. I., Negm, W. A., Elekhnawy, E., El-Masry, T. A., Attallah, N. G. M., Altwaijry, N., Batiha, G. E. -S., & El-Sherbeni, S. A. (2022). Antidiarrheal and Antibacterial Activities of Monterey Cypress Phytochemicals: In Vivo and In Vitro Approach. Molecules, 27(2), 346. https://doi.org/10.3390/molecules27020346