
Alzheimer’s Disease as a Major Public Health Concern: Role of Dietary Saponins in Mitigating Neurodegenerative Disorders and Their Underlying Mechanisms

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Global Prevalence and Risk Factors of AD
3. Pathological Targets of Alzheimer’s Disease
3.1. Brain Structure Alterations
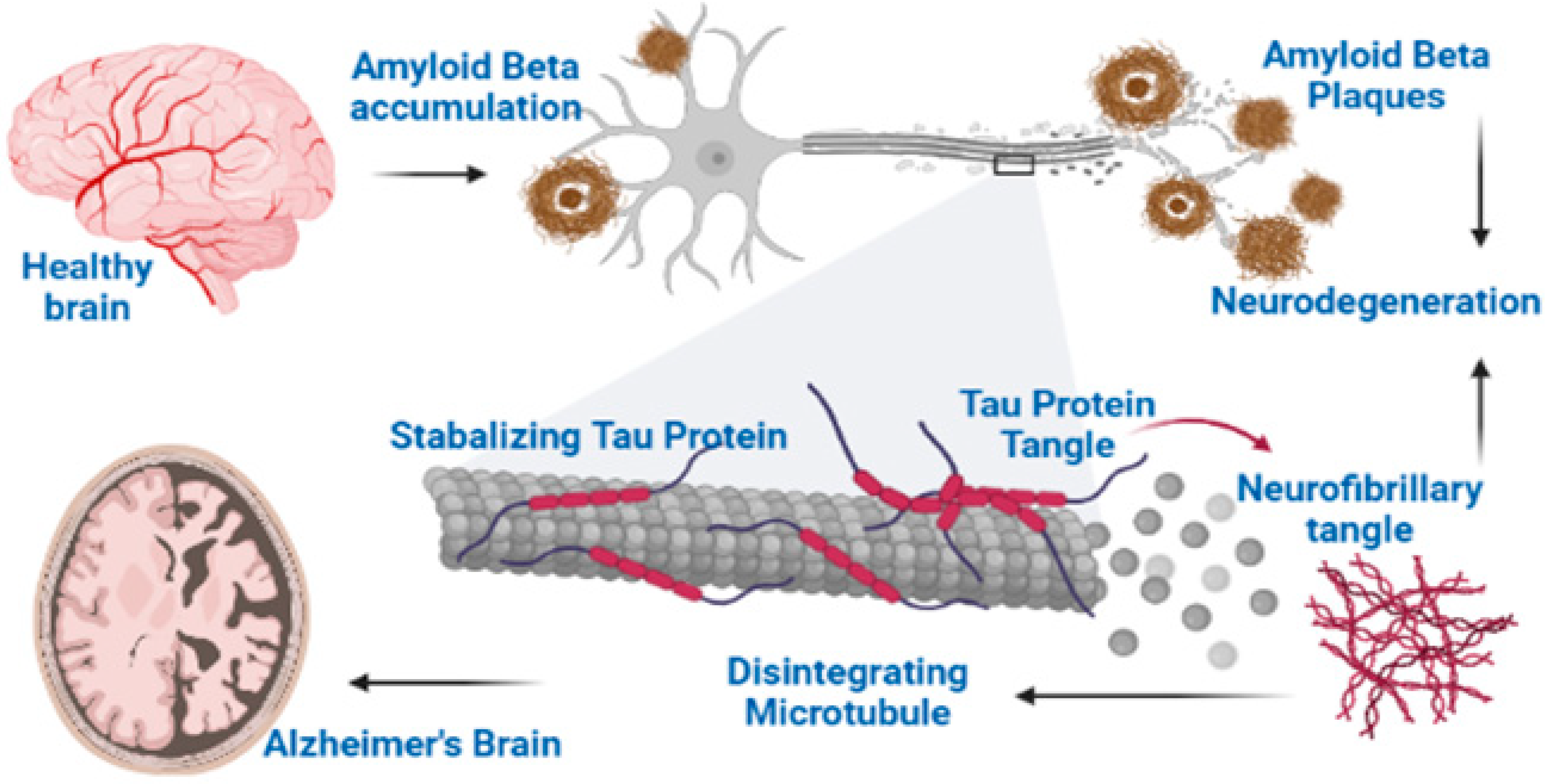
3.2. Progression of Degeneration in Alzheimer’s Disease
3.2.1. The β-Amyloid Hypothesis
3.2.2. APP Mutations Promote Longer Aβ Formation
3.2.3. Neurofibrillary Tangles (NFTs)
3.2.4. The Cholinergic Hypothesis
3.2.5. Oxidative Stress and Alzheimer’s Disease
3.2.6. Chronic Inflammation Hypothesis
3.2.7. Other Neurotransmitters Deficiency
4. Current Therapeutics against Alzheimer’s Disease
Plant-Based Alzheimer’s Disease Therapeutics
5. Saponins and Their Neuro-Pharmacological Properties
5.1. Brief Chemistry of Saponins
5.2. Occurrence and Distribution in Plants
6. Neuropharmacological Potentials of Saponins
6.1. Dementia and Age-Related Cognitive Decline
6.2. Anti-Amyloid and Anti-Neurofibrillay Tangles Potential of Saponins
6.3. Efficacy in Parkinsonism

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. no | Botanical Name | Compounds | Disease/Model | Neuroprotective Mechanism | Ref. |
|---|---|---|---|---|---|
| 1 | Gynostemma pentaphyllum | Gypenoside TN-2 | Learning deficit | Brain-derived neurotrophic factor (BDNF) and, cAMP-response element binding protein (CREB) | [104] |
| 2 | Liriope platyphylla | Spicatoside A | AD | Nerve growth factor (NGF), extracellular signal-regulated kinase (ERK), PI3-kinase/serine/threonine kinase (AKT), Cell surface transmembrane receptor tyrosine kinase (TrkA) receptor activation, neural networks reconstruction | [105] |
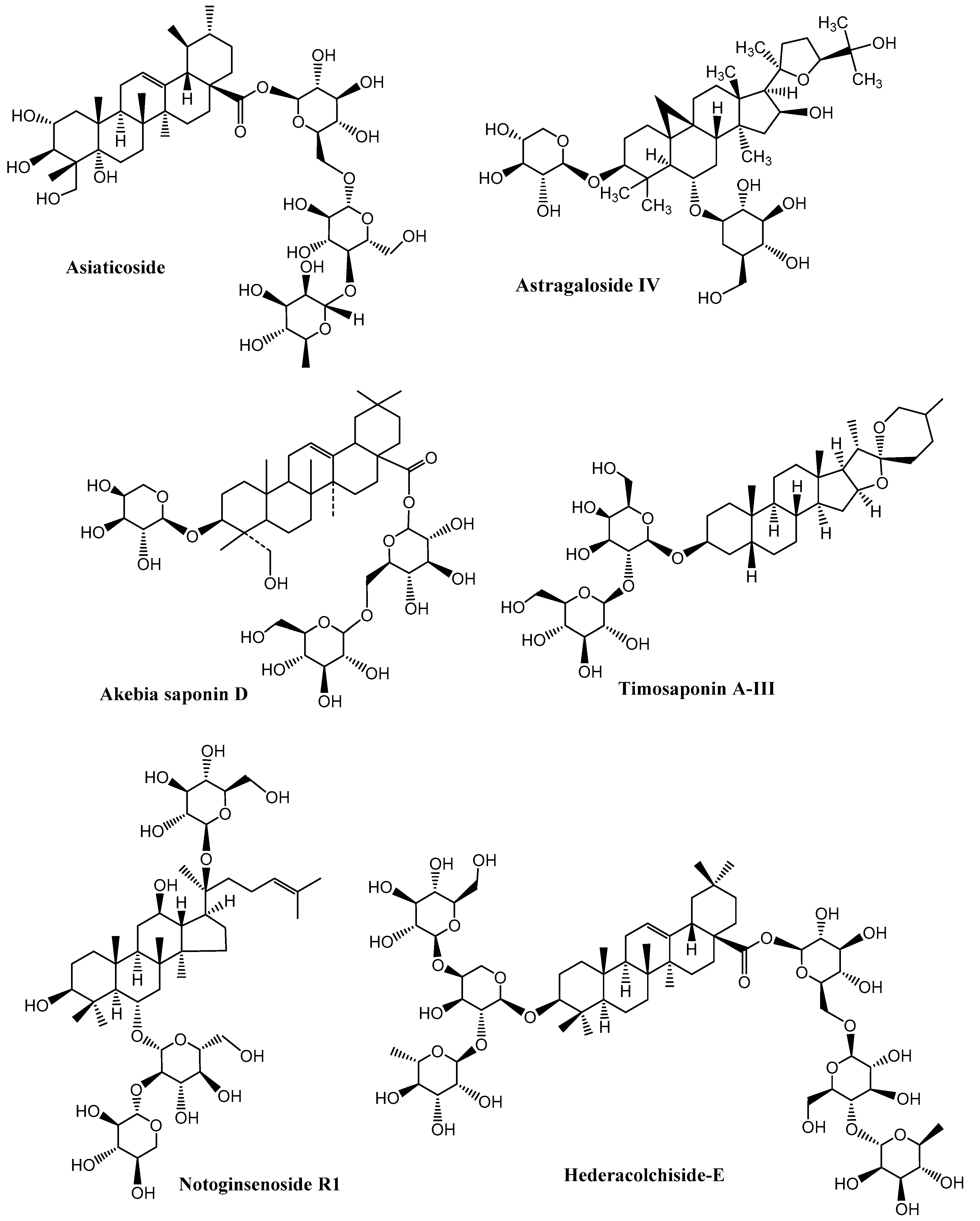
| 3 | Anemarrhena asphodeloides | Timosaponin AIII | AD | Acetylcholinesterase (AChE) Inhibition | [106] |
| 4 | Xanthoceras sorbifolia | Xanthoceraside | AD | Acetylcholinesterase (AChE) inhibition, antioxidant | [107] |
| 5 | Polygala tenuifolia | Onjisaponin F | AD | Nerve growth factor (NGF), Choline Acetyltransferase (ChAT) | [108] |
| 6 | Kalopanax pictus | Kalopanaxsaponins A | AD | Acetylcholinesterase (AChE) inhibition, cAMP-response element binding protein (p-CREB) | [109] |
| 7 | K. pictus | Kalopanaxsaponins B | AD | Acetylcholinesterase (AChE) inhibition, cAMP-response element, binding protein (p-CREB) | [109] |
| 8 | Astragalus membranaceus | Astragaloside IV | PD, Stroke | Antioxidant, tumor interleukin-1β (IL-1β), necrosis factor α (TNFα), Nuclear factor kappa B (NF-κB), Ca2+ influx, regeneration of the neural network | [74] |
| 9 | Dipsacus asper | Akebia saponin D | AD | mitogen-activated protein kinase (MAPK) anti- apoptosis | [99] |
| 10 | Panax ginseng | Ginsenoside Rb1 | AD, Stroke, HD | Antioxidant, Interleukin 6 (IL-6), necrosis factor α (TNF-α), anti-apoptosis, Ca2+ influx, Nerve growth factor (NGF), Glial Cell-Line Derived Neurotrophic Factor (GDNF), Brain-derived neurotrophic factor (BDNF), tau phosphorylation, NF-κB, PKA, Gβ1/PI3K/Akt, Ho-1, neurite outgrowth enhancing, TNF-α, NF-Κb | [74] |
| 11 | P. ginseng | Ginsenoside Rg1 | PD, AD, Stroke | Antioxidant, Tumor necrosis factor α (TNF-α), NO, BDNF, GDNF, IGF-IR, NGF, Nuclear factor kappa B (NF-κB), PKA, JNK, ER, AChE, signaling pathway, neural networks reconstruction | [74] |
| 12 | P. ginseng | Ginsenoside Rg3 | AD | Antioxidant, Tumor necrosis factor α (TNF-α), promote Aβ intake, iNOS, NMDA, interleukin-1β (IL-1β), AP-1, MSRA, PKA, | [74] |
| 13 | P. ginseng | Ginsenoside Rh2 | AD | Tumor necrosis factor α (TNF-α), NMDA, AP-1, JNK-AP-1, PKA | [110] |
| 14 | P. ginseng | Ginsenoside compound K | AD | Nuclear factor kappa B (NF-κB), Tumor necrosis factor α (TNF-α), interleukin-1β (IL-1β), GABA, iNOS, Intercellular adhesion molecule-1 (ICAM-1), JNK/activator protein-1 (AP-1)-signaling pathway (JNK-AP-1), | [111] |
| 15 | P. ginseng | Ginsenoside Re | AD | Inhibit BACE1 via activation of PPARγ, and reduce the generation of Aβ1–40 and Aβ1–42 | [112] |
| 16 | P. ginseng | Ginsenoside Rd | Stroke | Antioxidant, iNOS, cyclooxigenase-2 (COX-2), prostaglandins E2 (PGE2), Ca2+ influx, tau phosphorylation | [113,114] |
| 17 | P. ginseng | Ginsenoside Rg2 | Stroke | Anti-apoptosis | [115,116] |
| 18 | P. ginseng | Ginsenoside Rh3 | Microglia cells | iNOS, TNF-α, IL-1β | [117] |
| 19 | L. macranthoides | Akebiasaponin D | AD | antagonizes Aβ25-35-induced cytotoxicity in PC 12 cells | [118] |
| 20 | Panax notoginseng | Notoginsenoside R1 | Neuroprotection | NMDA, Bcl-2/Bax, Ca2+ influx | [119] |
| 21 | P. notoginseng | Notoginsenoside R4 | Neurite growth | Neural networks reconstruction | [120] |
| 22 | P. notoginseng | Notoginsenoside Fa | Neurite growth | Neural networks reconstruction | [120] |
| 23 | Platycodon grandiflorum | Platycodin D | Stroke | NF-κB, COX-2 | [121] |
| 24 | P. grandiflorum | 2″-o-Acetyl-polygalacin D2 | Stroke | NF-κB, COX-2 | [121] |
| 25 | White ginseng | Extract | AD | AChE/BChE | [122] |
| 26 | Red ginseng | Extract | AD | AChE/BChE | [122] |
| 27 | Black ginseng | Extract | AD | AChE/BChE, antioxidant | [122] |
| 28 | P. ginseng | Ginsenoside Rb3 | Neurite growth | Antioxidant, GABA receptor, neurite outgrowth enhancing | [123] |
| 29 | P. ginseng | Ginsenoside Rc | HD | Ca2+ signaling pathway | [124] |
| 30 | P. ginseng | Ginsenoside Rd | Stroke | Antioxidant, iNOS, COX-2, PGE2, Ca2+ influx, tau phosphorylation | [113,114] |
| 31 | Asparagus racemosus | Sarsasapogenin | AD | Inhibits AChE/BChE, MAO-B, Inhibits Aβ42) fibrillization | [76] |
| 32 | Green Tea | theasaponin E1 | AD | Inhibits tau hyperphosphorylation, declined Aβ levels, reduce inflammation | [77] |
| 33 | Panax notoginseng | Notoginsenoside R1 | Neuronal cells | B-cell lymphoma protein 2 (Bcl-2)-associated X (Bax), N-methyl-D-aspartate (NMDA) receptors, Ca2+ influx | [119] |
| 34 | C. asiatica | Asiaticoside | PD | Modulation of B-cell lymphoma protein 2 (Bcl-2)-associated X (Bax), free radicals, dopamine balance | [125] |
| 35 | Astragalus membranaceus | Astragaloside IV | PD, Stroke, | Suppression of free radicals, TNF-α, NF-κB, IL-1β, Ca2+ influx, Improvement in regeneration of the neural network | [74] |
7. Underlying Neuroprotective Mechanisms of Saponins
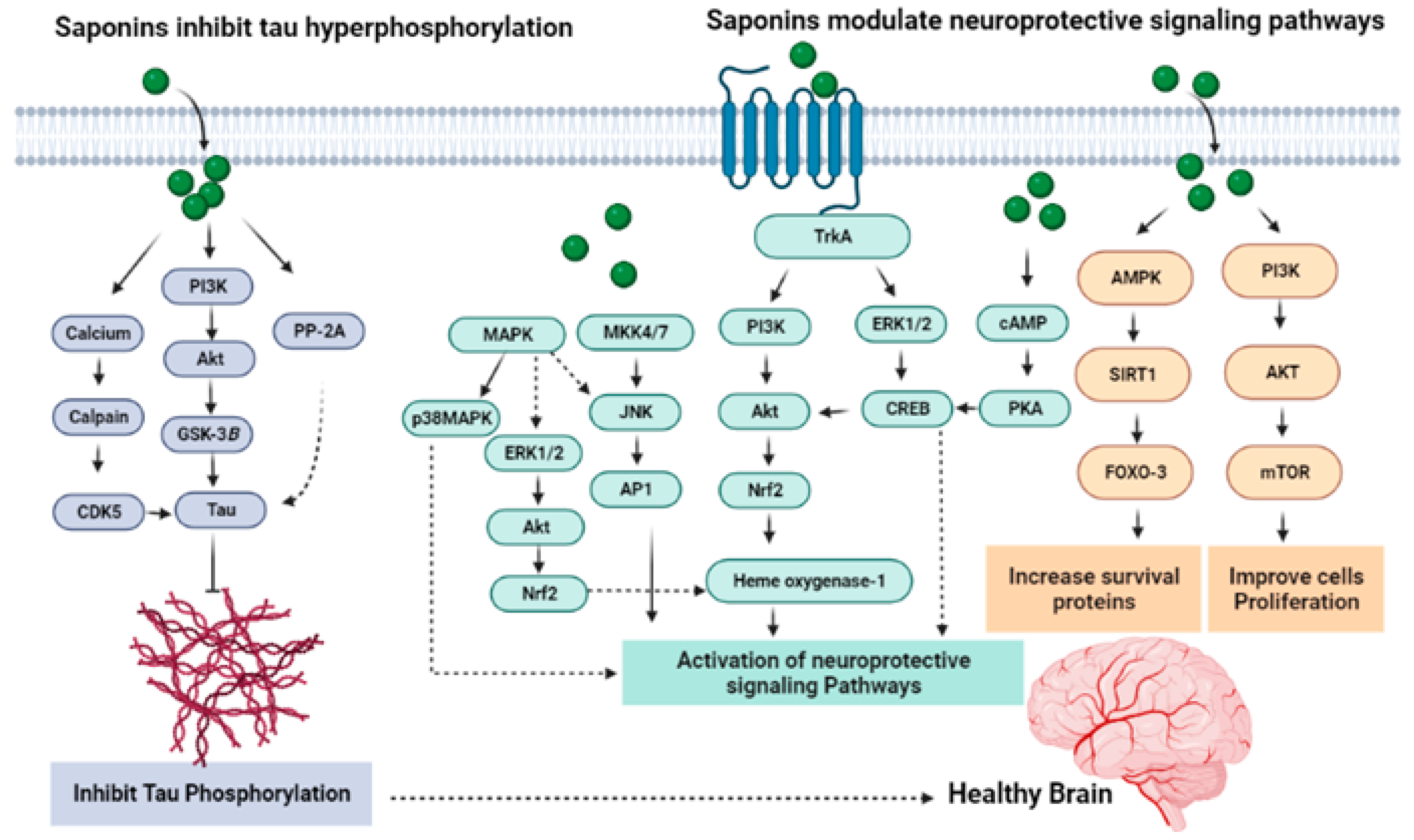
7.1. Mechanism Mediated through Antioxidant Activity
7.2. Mechanism Linked to Modulation of Neurotransmitters
7.3. Modulation of NMDA Receptors
7.4. Modulating Adenosine Receptors
7.5. Saponins Reduce Tau Phosphorylation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levey, A.I. Progress with treatments for Alzheimer’s disease. N. Engl. J. Med. 2021, 384, 1762–1763. [Google Scholar] [CrossRef] [PubMed]
- Jahn, H. Memory loss in Alzheimer’s disease. Dialogues Clin. Neurosci. 2013, 15, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, A.; Sun, H.; Han, Y.; Kong, L.; Wang, X. Two decades of new drug discovery and development for Alzheimer’s disease. RSC Adv. 2017, 7, 6046–6058. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.; Tam, K.Y. Pathological mechanisms and therapeutic strategies for Alzheimer’s disease. Neural Regen. Res. 2022, 17, 543. [Google Scholar]
- Ayaz, M.; Nawaz, A.; Naz, F.; Ullah, F.; Sadiq, A.; Islam, Z.U. Phytochemicals-based therapeutics against Alzheimer’s disease: An update. Curr. Top. Med. Chem. 2022. [Google Scholar] [CrossRef]
- Pinyopornpanish, K.; Soontornpun, A.; Wongpakaran, T.; Wongpakaran, N.; Tanprawate, S.; Pinyopornpanish, K.; Nadsasarn, A.; Pinyopornpanish, M. Impact of behavioral and psychological symptoms of Alzheimer’s disease on caregiver outcomes. Sci. Rep. 2022, 12, 1–9. [Google Scholar] [CrossRef]
- Ernst, R.L.; Hay, J.W. Economic research on Alzheimer disease: A review of the literature. Alzheimer Dis. Assoc. Disord. 1997, 11, 135–145. [Google Scholar]
- Karthika, C.; Appu, A.P.; Akter, R.; Rahman, M.H.; Tagde, P.; Ashraf, G.M.; Abdel-Daim, M.M.; Hassan, S.S.u.; Abid, A.; Bungau, S. Potential innovation against Alzheimer’s disorder: A tricomponent combination of natural antioxidants (vitamin E, quercetin, and basil oil) and the development of its intranasal delivery. Environ. Sci. Pollut. Res. 2022, 29, 10950–10965. [Google Scholar] [CrossRef]
- Qiu, C.; Kivipelto, M.; Von Strauss, E. Epidemiology of Alzheimer’s disease: Occurrence, determinants, and strategies toward intervention. Dialogues Clin. Neurosci. 2009, 11, 111–128. [Google Scholar] [CrossRef]
- Alzheimer’s Association. Alzheimer’s disease facts and figures. Alzheimers Dement. 2020, 16, 391–460. [Google Scholar]
- Conrado, D.J.; Duvvuri, S.; Geerts, H.; Burton, J.; Biesdorf, C.; Ahamadi, M.; Macha, S.; Hather, G.; Francisco Morales, J.; Podichetty, J. Challenges in Alzheimer’s Disease Drug Discovery and Development: The role of modeling, simulation, and open data. Clin. Pharmacol. Ther. 2020, 107, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Hebert, L.E.; Weuve, J.; Scherr, P.A.; Evans, D.A. Alzheimer disease in the United States (2010–2050) estimated using the 2010 census. Neurology 2013, 80, 1778–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, D.A. Estimated prevalence of Alzheimer’s disease in the United States. Milbank Q. 1990, 68, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Pandey, P.; Sharma, P. Understanding the genetic, molecular, and cellular basis of ageing as the biggest risk factor of Alzheimer’s disease. Eur. J. Biol. Res. 2022, 12, 62–76. [Google Scholar]
- Brookmeyer, R.; Gray, S.; Kawas, C. Projections of Alzheimer’s disease in the United States and the public health impact of delaying disease onset. Am. J. Public Health 1998, 88, 1337–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzin, J.; Lang, K.; Friedman, M.; Neumann, P.; Cummings, J.L. The economic cost of Alzheimer’s disease and related dementias to the California Medicaid program (“Medi-Cal”) in 1995. Am. J. Geriatr. Psychiatry 1999, 7, 300–308. [Google Scholar] [CrossRef]
- Omura, J.D.; McGuire, L.C.; Patel, R.; Baumgart, M.; Lamb, R.; Jeffers, E.M.; Olivari, B.S.; Croft, J.B.; Thomas, C.W.; Hacker, K. Modifiable risk factors for Alzheimer disease and related dementias among adults aged ≥45 years—United States, 2019. Morb. Mortal. Wkly. Rep. 2022, 71, 680–685. [Google Scholar]
- Morrison, A.S.; Lyketsos, C. The pathophysiology of Alzheimer’s disease and directions in treatment. Adv. Stud. Nurs. 2005, 3, 256–270. [Google Scholar]
- Förstl, H.; Zerfaß, R.; Geiger-Kabisch, C.; Sattel, H.; Besthorn, C.; Hentschel, F. Brain atrophy in normal ageing and Alzheimer’s disease: Volumetric discrimination and clinical correlations. Br. J. Psychiatry 1995, 167, 739–746. [Google Scholar]
- Hippius, H.; Neundörfer, G. The discovery of Alzheimer’s disease. Dialogues Clin. Neurosci. 2003, 5, 101–108. [Google Scholar] [CrossRef]
- Bianchetti, A.; Ranieri, P.; Margiotta, A.; Trabucchi, M. Pharmacological treatment of Alzheimer’s Disease. Aging Clin. Exp. Res. 2006, 18, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Ovais, M.; Zia, N.; Ahmad, I.; Khalil, A.T.; Raza, A.; Ayaz, M.; Sadiq, A.; Ullah, F.; Shinwari, Z.K. Phyto-Therapeutic and Nanomedicinal Approach to Cure Alzheimer Disease: Present Status and Future Opportunities. Front. Aging Neurosci. 2018, 10, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.X.; Yan, S.D. Amyloid-β-induced mitochondrial dysfunction. J. Alzheimer’s Dis. 2007, 12, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Su, B.; Perry, G.; Smith, M.A.; Zhu, X. Insights into amyloid-β-induced mitochondrial dysfunction in Alzheimer disease. Free Radic. Biol. Med. 2007, 43, 1569–1573. [Google Scholar] [CrossRef]
- Tong, X.; Li, X.; Ayaz, M.; Ullah, F.; Sadiq, A.; Ovais, M.; Shahid, M.; Khayrullin, M.; Hazrat, A. Neuroprotective studies on Polygonum hydropiper L. essential oils using transgenic animal models. Front. Pharmacol. 2020, 11, 580069. [Google Scholar] [CrossRef]
- Ayaz, M.; Sadiq, A.; Junaid, M.; Ullah, F.; Subhan, F.; Ahmed, J. Neuroprotective and anti-aging potentials of essential oils from aromatic and medicinal plants. Front. Aging Neurosci. 2017, 9, 168. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, M.; Sadiq, A.; Junaid, M.; Ullah, F.; Ovais, M.; Ullah, I.; Ahmed, J.; Shahid, M. Flavonoids as prospective neuroprotectants and their therapeutic propensity in aging associated neurological disorders. Front. Aging Neurosci. 2019, 11, 155. [Google Scholar] [CrossRef]
- Suzuki, N.; Cheung, T.T.; Cai, X.-D.; Odaka, A.; Otvos, L., Jr.; Eckman, C.; Golde, T.E.; Younkin, S.G. An increased percentage of long amyloid β protein secreted by familial amyloid β protein precursor (βAPP717) mutants. Science 1994, 264, 1336–1340. [Google Scholar] [CrossRef]
- Rong, X.; Jiang, L.; Qu, M.; Hassan, S.S.u.; Liu, Z. Enhancing Therapeutic Efficacy of Donepezil by Combined Therapy: A Comprehensive Review. Curr. Pharm. Des. 2020, 27, 332–344. [Google Scholar] [CrossRef]
- Kaur, D.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Badavath, V.N.; Hassan, S.S.u.; Hasan, M.M.; Bhatia, S.; Al-Harassi, A.; et al. Unravelling the potential neuroprotective facets of erythropoietin for the treatment of Alzheimer’s disease. Metab. Brain Dis. 2022, 37, 1–16. [Google Scholar] [CrossRef]
- Brion, J.-P. Neurofibrillary tangles and Alzheimer’s disease. Eur. Neurol. 1998, 40, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Amber, S.; Zahid, S.; Malik, N. Rosmarinus officinalis and Methylphenidate Exposure Improves Cognition and Depression and Regulates Anxiety like Behavior in AlCl3-induced Mouse Model of Alzheimer’s Disease. Front. Pharmacol. 2022, 13, 943163. [Google Scholar] [CrossRef]
- Perry, E.K.; Tomlinson, B.E.; Blessed, G.; Bergmann, K.; Gibson, P.H.; Perry, R.H. Correlation of cholinergic abnormalities with senile plaques and mental test scores in senile dementia. Br. Med. J. 1978, 2, 1457–1459. [Google Scholar] [CrossRef] [Green Version]
- Mir, N.T.; Saleem, U.; Anwar, F.; Ahmad, B.; Ullah, I.; Hira, S.; Ismail, T.; Ali, T.; Ayaz, M. Lawsonia Inermis Markedly Improves Cognitive Functions in Animal Models and Modulate Oxidative Stress Markers in the Brain. Medicina 2019, 55, 192. [Google Scholar] [CrossRef] [Green Version]
- Saleem, U.; Akhtar, R.; Anwar, F.; Shah, M.A.; Chaudary, Z.; Ayaz, M.; Ahmad, B. Neuroprotective potential of Malva neglecta is mediated via down-regulation of cholinesterase and modulation of oxidative stress markers. Metab. Brain Dis. 2021, 36, 889–900. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Griffin, S.; Munch, G.; Pasinetti, G.M. Amyloid β-peptide and amyloid pathology are central to the oxidative stress and inflammatory cascades under which Alzheimer’s disease brain exists. J. Alzheimer’s Dis. 2002, 4, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Raina, A.K.; Smith, M.A. Cell cycle events in neurons: Proliferation or death? Am. J. Pathol. 1999, 155, 327–329. [Google Scholar] [CrossRef]
- Zhu, X.; Lee, H.-G.; Casadesus, G.; Avila, J.; Drew, K.; Perry, G.; Smith, M.A. Oxidative imbalance in Alzheimer’s disease. Mol. Neurobiol. 2005, 31, 205–217. [Google Scholar] [CrossRef]
- Rozemuller, J.; Eikelenboom, P.; Stam, F. Role of microglia in plaque formation in senile dementia of the Alzheimer type. Virchows Arch. B 1986, 51, 247–254. [Google Scholar] [CrossRef]
- Hassan, S.S.u.; Abdel-Daim, M.M.; Behl, T.; Bungau, S. Natural Products for Chronic Diseases: A Ray of Hope. Molecules 2022, 27, 5573. [Google Scholar] [CrossRef]
- Rogers, J.; Shen, Y. A perspective on inflammation in Alzheimer’s disease. Ann. N.Y. Acad. Sci. 2000, 924, 132–135. [Google Scholar] [CrossRef]
- Zotova, E.; Nicoll, J.A.; Kalaria, R.; Holmes, C.; Boche, D. Inflammation in Alzheimer’s disease: Relevance to pathogenesis and therapy. Alzheimers Res. Ther. 2010, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Barger, S.; Barnum, S.; Bradt, B.; Bauer, J.; Cole, G.; Cooper, N.; Eikelenboom, P.; Emmerling, M.; Fiebich, B.L.; et al. Inflammation and Alzheimer’s disease. Neurobiol. Aging 2000, 21, 383–421. [Google Scholar] [CrossRef]
- Kalaria, R.N. Microglia and Alzheimer’s disease. Curr. Opin. Hematol. 1999, 6, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, L.; Robakis, N.K.; Figueiredo-Pereira, M.E. It may take inflammation, phosphorylation and ubiquitination to ‘tangle’in Alzheimer’s disease. Neurodegener. Dis. 2006, 3, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Gorlovoy, P.; Larionov, S.; Pham, T.T.H.; Neumann, H. Accumulation of tau induced in neurites by microglial proinflammatory mediators. FASEB J. 2009, 23, 2502–2513. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, J.A.; Mrak, R.E.; Graham, D.I.; Stewart, J.; Wilcock, G.; MacGowan, S.; Esiri, M.M.; Murray, L.S.; Dewar, D.; Love, S. Association of interleukin-1 gene polymorphisms with Alzheimer’s disease. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 2000, 47, 365–368. [Google Scholar] [CrossRef]
- Hayes, A.; Green, E.; Pritchard, A.; Harris, J.; Zhang, Y.; Lambert, J.; Chartier-Harlin, M.; Pickering-Brown, S.; Lendon, C.L.; Mann, D.M. A polymorphic variation in the interleukin 1A gene increases brain microglial cell activity in Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 2004, 75, 1475–1477. [Google Scholar] [CrossRef] [Green Version]
- Culpan, D.; MacGowan, S.H.; Ford, J.M.; Nicoll, J.A.; Griffin, W.S.; Dewar, D.; Cairns, N.J.; Hughes, A.; Kehoe, P.G.; Wilcock, G.K. Tumour necrosis factor-α gene polymorphisms and Alzheimer’s disease. Neurosci. Lett. 2003, 350, 61–65. [Google Scholar] [CrossRef]
- Horsburgh, K.; McCarron, M.O.; White, F.; Nicoll, J.A. The role of apolipoprotein E in Alzheimer’s disease, acute brain injury and cerebrovascular disease: Evidence of common mechanisms and utility of animal models. Neurobiol. Aging 2000, 21, 245–255. [Google Scholar] [CrossRef]
- Egensperger, R.; Kösel, S.; von Eitzen, U.; Graeber, M.B. Microglial activation in Alzheimer disease: Association with APOE genotype. Brain Pathol. 1998, 8, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Galea, E.; Heneka, M.T.; Russo, C.D.; Feinstein, D.L. Intrinsic regulation of brain inflammatory responses. Cell. Mol. Neurobiol. 2003, 23, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, N.; Lanctôt, K.L.; Khan, L.R. The role of norepinephrine in the behavioral and psychological symptoms of dementia. J. Neuropsychiatry Clin. Neurosci. 2004, 16, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Junaid, M.; Ullah, F.; Sadiq, A.; Khan, M.A.; Ahmad, W.; Shah, M.R.; Imran, M.; Ahmad, S. Comparative chemical profiling, cholinesterase inhibitions and anti-radicals properties of essential oils from Polygonum hydropiper L: A Preliminary anti-Alzheimer’s study. Lipids Health Dis. 2015, 14, 141. [Google Scholar] [CrossRef] [Green Version]
- Khalil, A.T.; Ayaz, M.; Ovais, M.; Wadood, A.; Ali, M.; Shinwari, Z.K.; Maaza, M. In vitro cholinesterase enzymes inhibitory potential and in silico molecular docking studies of biogenic metal oxides nanoparticles. Inorg. Nano-Met. Chem. 2018, 48, 441–448. [Google Scholar] [CrossRef]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Kim, M.O.; Ali, T. Natural products-based drugs: Potential therapeutics against Alzheimer’s disease and other neurological disorders. Front. Pharmacol. 2019, 10, 1417. [Google Scholar] [CrossRef] [Green Version]
- Faheem, M.; Shah, F.A.; Khan, A.U.; Li, S.P. Investigation of Natural Isolated Compounds for Therapeutic Potential in Streptozotocin-induced Diabetic Neuroinflammation, Neurodegeneration and Neuropathic Pain. Front. Pharmacol. 2022. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, T.; Sadiq, A.; Ullah, F.; Naseer, M.I. Current Trends in Medicinal Plant Research and Neurodegenerative Disorders. Front. Media SA 2022, 13, 922373. [Google Scholar]
- Ratheesh, G.; Tian, L.; Venugopal, J. Role of medicinal plants in neurodegenerative diseases. Biomanuf. Rev. 2017, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Lim, G.P.; Begum, A.N.; Ubeda, O.J.; Simmons, M.R.; Ambegaokar, S.S.; Chen, P.P.; Kayed, R.; Glabe, C.G.; Frautschy, S.A. Curcumin inhibits formation of amyloid β oligomers and fibrils, binds plaques, and reduces amyloid in vivo. J. Biol. Chem. 2005, 280, 5892–5901. [Google Scholar] [CrossRef] [Green Version]
- McClure, R.; Ong, H.; Janve, V.; Barton, S.; Zhu, M.; Li, B.; Dawes, M.; Jerome, W.G.; Anderson, A.; Massion, P. Aerosol delivery of curcumin reduced amyloid-β deposition and improved cognitive performance in a transgenic model of Alzheimer’s disease. J. Alzheimer’s Dis. 2017, 55, 797–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.H.; Manczak, M.; Yin, X.; Grady, M.C.; Mitchell, A.; Tonk, S.; Kuruva, C.S.; Bhatti, J.S.; Kandimalla, R.; Vijayan, M. Protective effects of Indian spice curcumin against amyloid-β in Alzheimer’s disease. J. Alzheimer’s Dis. 2018, 61, 843–866. [Google Scholar] [CrossRef]
- Ono, K.; Yoshiike, Y.; Takashima, A.; Hasegawa, K.; Naiki, H.; Yamada, M. Potent anti-amyloidogenic and fibril-destabilizing effects of polyphenols in vitro: Implications for the prevention and therapeutics of Alzheimer’s disease. J. Neurochem. 2003, 87, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D. Effect of flavonoids on learning, memory and neurocognitive performance: Relevance and potential implications for Alzheimer’s disease pathophysiology. J. Sci. Food Agric. 2014, 94, 1042–1056. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-M.; Kim, B.C.; Cho, Y.-H.; Choi, K.-H.; Chang, J.; Park, M.-S.; Kim, M.-K.; Cho, K.-H.; Kim, J.-K. Effects of flavonoid compounds on β-amyloid-peptide-induced neuronal death in cultured mouse cortical neurons. Chonnam Med. J. 2014, 50, 45–51. [Google Scholar] [CrossRef] [Green Version]
- DeKosky, S.T.; Scheff, S.W. Synapse loss in frontal cortex biopsies in Alzheimer’s disease: Correlation with cognitive severity. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 1990, 27, 457–464. [Google Scholar] [CrossRef]
- Jackson, M.; Gentleman, S.; Lennox, G.; Ward, L.; Gray, T.; Randall, K.; Morrell, K.; Lowe, J. The cortical neuritic pathology of Huntington’s disease. Neuropathol. Appl. Neurobiol. 1995, 21, 18–26. [Google Scholar] [CrossRef]
- Hassan, S.S.u.; Muhammad, I.; Abbas, S.Q.; Hassan, M.; Majid, M.; Jin, H.Z.; Bungau, S. Stress driven discovery of natural products from actinobacteria with anti-oxidant and cytotoxic activities including docking and admet properties. Int. J. Mol. Sci. 2021, 22, 11432. [Google Scholar] [CrossRef]
- Mattila, P.; Rinne, J.; Helenius, H.; Röyttä, M. Neuritic degeneration in the hippocampus and amygdala in Parkinson’s disease in relation to Alzheimer pathology. Acta Neuropathol. 1999, 98, 157–164. [Google Scholar] [CrossRef]
- Ayaz, M.; Junaid, M.; Ullah, F.; Sadiq, A.; Subhan, F.; Khan, M.A.; Ahmad, W.; Ali, G.; Imran, M.; Ahmad, S. Molecularly characterized solvent extracts and saponins from Polygonum hydropiper L. show high anti-angiogenic, anti-tumor, brine shrimp, and fibroblast NIH/3T3 cell line cytotoxicity. Front. Pharmacol. 2016, 7, 74. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Henkel, T. Traditional Chinese medicine (TCM): Are polyphenols and saponins the key ingredients triggering biological activities? Curr. Med. Chem. 2002, 9, 1483–1485. [Google Scholar] [CrossRef] [PubMed]
- Güçlü-Üstündağ, Ö.; Mazza, G. Saponins: Properties, applications and processing. Crit. Rev. Food Sci. Nutr. 2007, 47, 231–258. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.; Xu, X.; Lin, J.; Cui, X.; Xu, R. Neuroprotection by saponins. Phytother. Res. 2015, 29, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Oyeleke, M.B.; Owoyele, B.V. Saponins and flavonoids from Bacopa floribunda plant extract exhibit antioxidant and anti-inflammatory effects on amyloid beta 1-42-induced Alzheimer’s disease in BALB/c mice. J. Ethnopharmacol. 2022, 288, 114997. [Google Scholar] [CrossRef]
- Kashyap, P.; Muthusamy, K.; Niranjan, M.; Trikha, S.; Kumar, S. Sarsasapogenin: A steroidal saponin from Asparagus racemosus as multi target directed ligand in Alzheimer’s disease. Steroids 2020, 153, 108529. [Google Scholar] [CrossRef]
- Khan, M.I.; Khan, M.Z.; Shin, J.H.; Shin, T.S.; Lee, Y.B.; Kim, M.Y.; Kim, J.D. Neuroprotective Effects of Green Tea Seed Isolated Saponin Due to the Amelioration of Tauopathy and Alleviation of Neuroinflammation: A Therapeutic Approach to Alzheimer’s Disease. Molecules 2022, 27, 2079. [Google Scholar] [CrossRef]
- Vincken, J.-P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef]
- El Aziz, M.; Ashour, A.; Melad, A. A review on saponins from medicinal plants: Chemistry, isolation, and determination. J. Nanomed. Res. 2019, 8, 282–288. [Google Scholar]
- Dinda, B.; Debnath, S.; Mohanta, B.C.; Harigaya, Y. Naturally occurring triterpenoid saponins. Chem. Biodivers. 2010, 7, 2327–2580. [Google Scholar] [CrossRef]
- Sparg, S.; Light, M.; Van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef]
- Parente, J.P.; da Silva, B.P. Bioactive complex triterpenoid saponins from the Leguminosae family. Nat. Prod. Commun. 2009, 4, 1934578X0900400131. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Arunasalam, K.; Yeung, D.; Kakuda, Y.; Mittal, G.; Jiang, Y. Saponins from edible legumes: Chemistry, processing, and health benefits. J. Med. Food 2004, 7, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Osbourn, A.E. Saponins in cereals. Phytochemistry 2003, 62, 1–4. [Google Scholar] [CrossRef]
- Hoffmann, D. Medical Herbalism: The Science and Practice of Herbal Medicine; Simon and Schuster: New York, NY, USA, 2003. [Google Scholar]
- Hostettmann, K.; Marston, A. Saponins; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- He, L.; Mu, L.; Jean, J.A.; Zhang, L.; Wu, H.; Zhou, T.; Bu, H. Contributions and Challenges of Public Health Social Work Practice during the Initial 2020 COVID-19 Outbreak in China. Br. J. Soc. Work. 2022. [Google Scholar] [CrossRef]
- He, X.; Zhu, Y.; Yang, L.; Wang, Z.; Wang, Z.; Feng, J.; Wen, X.; Cheng, L.; Zhu, R. MgFe-LDH Nanoparticles: A Promising Leukemia Inhibitory Factor Replacement for Self-Renewal and Pluripotency Maintenance in Cultured Mouse Embryonic Stem Cells. Adv. Sci. 2021, 8, 2003535. [Google Scholar] [CrossRef] [PubMed]
- Roddick, J.; Melchers, G. Steroidal glycoalkaloid content of potato, tomato and their somatic hybrids. Theor. Appl. Genet. 1985, 70, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.P.; Spink, J.; Cannon, P.F.; Daniels, M.J.; Osbourn, A.E. Isolation, characterization, and avenacin sensitivity of a diverse collection of cereal-root-colonizing fungi. Appl. Environ. Microbiol. 1999, 65, 3364–3372. [Google Scholar] [CrossRef] [Green Version]
- Field, B.; Jordán, F.; Osbourn, A. First encounters–deployment of defence-related natural products by plants. New Phytol. 2006, 172, 193–207. [Google Scholar] [CrossRef]
- Hostettmann, K.; Marston, A. Chemistry and Pharmacology of Natural Products; Cambridge University Press: Cambridge, UK, 1995; Volume 548. [Google Scholar]
- Tohda, C.; Matsumoto, N.; Zou, K.; Meselhy, M.R.; Komatsu, K. A β (25–35)-induced memory impairment, axonal atrophy, and synaptic loss are ameliorated by M1, a metabolite of protopanaxadiol-type saponins. Neuropsychopharmacology 2004, 29, 860–868. [Google Scholar] [CrossRef]
- Cheng, Y.; Shen, L.H.; Zhang, J.T. Anti-amnestic and anti-aging effects of ginsenoside Rg1 and Rb1 and its mechanism of action. ACTA Pharmacol. Sin. 2005, 26, 143–149. [Google Scholar] [CrossRef]
- Chen, F.; Eckman, E.A.; Eckman, C.B. Reductions in levels of the Alzheimer’s amyloid beta peptide after oral administration of ginsenosides. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2006, 20, 1269–1271. [Google Scholar]
- Han, C.-K.; Choi, W.R.; Oh, K.-B. Cognition-enhancing and neuroprotective effects of hederacolchiside-E from Pulsatilla koreana. Planta Med. 2007, 73, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.-H.; Liu, Y.; Hu, H.-T.; Ren, H.-M.; Chen, X.-L.; Xu, J.-H. The effects of the total saponin of Dipsacus asperoides on the damage of cultured neurons induced by β-amyloid protein 25–35. Anat. Sci. Int. 2002, 77, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Long, X.; Chen, H.; Ji, Z.; Shu, B.; Yue, R.; Liao, Y.; Ma, S.; Qiao, K.; Liu, Y.; et al. Photoclick Reaction Constructs Glutathione-Responsive Theranostic System for Anti-Tuberculosis. Front. Mol. Biosci. 2022, 9, 845179. [Google Scholar] [CrossRef]
- Yu, X.; Wang, L.-N.; Ma, L.; You, R.; Cui, R.; Ji, D.; Wu, Y.; Zhang, C.-F.; Yang, Z.-L.; Ji, H. Akebia saponin D attenuates ibotenic acid-induced cognitive deficits and pro-apoptotic response in rats: Involvement of MAPK signal pathway. Pharmacol. Biochem. Behav. 2012, 101, 479–486. [Google Scholar] [CrossRef]
- Luo, F.-C.; Wang, S.-D.; Qi, L.; Song, J.-Y.; Lv, T.; Bai, J. Protective effect of panaxatriol saponins extracted from Panax notoginseng against MPTP-induced neurotoxicity in vivo. J. Ethnopharmacol. 2011, 133, 448–453. [Google Scholar] [CrossRef]
- Xu, B.-B.; Liu, C.-Q.; Gao, X.; Zhang, W.-Q.; Wang, S.-W.; Cao, Y.-L. Possible mechanisms of the protection of ginsenoside Re against MPTP-induced apoptosis in substantia nigra neurons of Parkinson’s disease mouse model. J. Asian Nat. Prod. Res. 2005, 7, 215–224. [Google Scholar] [CrossRef]
- Wang, J.; Xu, H.-M.; Yang, H.-D.; Du, X.-X.; Jiang, H.; Xie, J.-X. Rg1 reduces nigral iron levels of MPTP-treated C57BL6 mice by regulating certain iron transport proteins. Neurochem. Int. 2009, 54, 43–48. [Google Scholar] [CrossRef]
- Xu, C.-L.; Qu, R.; Zhang, J.; Li, L.-F.; Ma, S.-P. Neuroprotective effects of madecassoside in early stage of Parkinson’s disease induced by MPTP in rats. Fitoterapia 2013, 90, 112–118. [Google Scholar] [CrossRef]
- Hong, S.-W.; Yang, J.-H.; Joh, E.-H.; Kim, H.J.; Kim, D.-H. Gypenoside TN-2 ameliorates scopolamine-induced learning deficit in mice. J. Ethnopharmacol. 2011, 134, 1010–1013. [Google Scholar] [CrossRef]
- Hur, J.; Lee, P.; Moon, E.; Kang, I.; Kim, S.-H.; Oh, M.S.; Kim, S.Y. Neurite outgrowth induced by spicatoside A, a steroidal saponin, via the tyrosine kinase A receptor pathway. Eur. J. Pharmacol. 2009, 620, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Jung, K.; Kim, D.-H. Timosaponin AIII, a saponin isolated from Anemarrhena asphodeloides, ameliorates learning and memory deficits in mice. Pharmacol. Biochem. Behav. 2009, 93, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Chi, T.-Y.; Wang, L.-H.; Qu, C.; Yang, B.-Z.; Ji, X.-F.; Wang, Y.; Okuyama, T.; Yoshihito, O.; Zou, L.-B. Protective effects of xanthoceraside on learning and memory impairment induced by Aβ25–35 in mice. J. Asian Nat. Prod. Res. 2009, 11, 1019–1027. [Google Scholar] [CrossRef]
- Yabe, T.; Tuchida, H.; Kiyohara, H.; Takeda, T.; Yamada, H. Induction of NGF synthesis in astrocytes by onjisaponins of Polygala tenuifolia, constituents of kampo (Japanese herbal) medicine, Ninjin-yoei-to. Phytomedicine 2003, 10, 106–114. [Google Scholar] [CrossRef]
- Joh, E.H.; Lee, I.A.; Kim, D.H. Kalopanaxsaponins A and B isolated from Kalopanax pictus ameliorate memory deficits in mice. Phytother. Res. 2012, 26, 546–551. [Google Scholar] [CrossRef]
- Lee, E.; Kim, S.; Chung, K.C.; Choo, M.-K.; Kim, D.-H.; Nam, G.; Rhim, H. 20 (S)-ginsenoside Rh2, a newly identified active ingredient of ginseng, inhibits NMDA receptors in cultured rat hippocampal neurons. Eur. J. Pharmacol. 2006, 536, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.C.; Chen, Y.F. Effect of convalescent toy image design on memory recovery in patients with Alzheimer’s disease. Indian J. Pharm. Sci. 2020, 82, 5. [Google Scholar]
- Cao, G.; Su, P.; Zhang, S.; Guo, L.; Zhang, H.; Liang, Y.; Qin, C.; Zhang, W. Ginsenoside Re reduces Aβ production by activating PPARγ to inhibit BACE1 in N2a/APP695 cells. Eur. J. Pharmacol. 2016, 793, 101–108. [Google Scholar] [CrossRef]
- Ye, R.; Han, J.; Kong, X.; Zhao, L.; Cao, R.; Rao, Z.; Zhao, G. Protective effects of ginsenoside Rd on PC12 cells against hydrogen peroxide. Biol. Pharm. Bull. 2008, 31, 1923–1927. [Google Scholar] [CrossRef] [Green Version]
- Ye, R.; Li, N.; Han, J.; Kong, X.; Cao, R.; Rao, Z.; Zhao, G. Neuroprotective effects of ginsenoside Rd against oxygen-glucose deprivation in cultured hippocampal neurons. Neurosci. Res. 2009, 64, 306–310. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, A.; Zhou, Y.; San, X.; Jin, T.; Jin, Y. Panax ginseng ginsenoside-Rg2 protects memory impairment via anti-apoptosis in a rat model with vascular dementia. J. Ethnopharmacol. 2008, 115, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-F.; Fan, X.-J.; Li, X.; Peng, L.-L.; Wang, G.-H.; Ke, K.-F.; Jiang, Z.-L. Ginsenoside Rg1 protects neurons from hypoxic–ischemic injury possibly by inhibiting Ca2+ influx through NMDA receptors and L-type voltage-dependent Ca2+ channels. Eur. J. Pharmacol. 2008, 586, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-S.; Park, E.-M.; Kim, D.-H.; Jung, K.; Jung, J.-S.; Lee, E.-J.; Hyun, J.-W.; Kang, J.L.; Kim, H.-S. Anti-inflammatory mechanism of ginseng saponins in activated microglia. J. Neuroimmunol. 2009, 209, 40–49. [Google Scholar] [CrossRef]
- Zhou, Y.-Q.; Yang, Z.-L.; Xu, L.; Li, P.; Hu, Y.-Z. Akebia saponin D, a saponin component from Dipsacus asper Wall, protects PC 12 cells against amyloid-β induced cytotoxicity. Cell Biol. Int. 2009, 33, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Nakamichi, N.; Zhang, W.S.; Nakamura, Y.; Kambe, Y.; Fukumori, R.; Takuma, K.; Yamada, K.; Takarada, T.; Taniura, H. Possible protection by notoginsenoside R1 against glutamate neurotoxicity mediated by N-methyl-D-aspartate receptors composed of an NR1/NR2B subunit assembly. J. Neurosci. Res. 2009, 87, 2145–2156. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Zhu, S.; Meselhy, M.R.; Tohda, C.; Cai, S.; Komatsu, K. Dammarane-Type Saponins from Panax j aponicus and Their Neurite Outgrowth Activity in SK-N-SH Cells. J. Nat. Prod. 2002, 65, 1288–1292. [Google Scholar] [CrossRef]
- Choi, J.H.; Yoo, K.-Y.; Park, O.K.; Lee, C.H.; Won, M.-H.; Hwang, I.K.; Ryu, S.Y.; Kim, Y.S.; Yi, J.-S.; Bae, Y.-S. Platycodin D and 2 ″-o-acetyl-polygalacin D2 isolated from Platycodon grandiflorum protect ischemia/reperfusion injury in the gerbil hippocampus. Brain Res. 2009, 1279, 197–208. [Google Scholar] [CrossRef]
- Lee, M.R.; Yun, B.S.; In, O.H.; Sung, C.K. Comparative study of Korean white, red, and black ginseng extract on cholinesterase inhibitory activity and cholinergic function. J. Ginseng Res. 2011, 35, 421. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Kim, S.; Markelonis, G.; Oh, T. Ginsenosides Rb1 and Rg3 protect cultured rat cortical cells from glutamate-induced neurodegeneration. J Neurosci Res 53: 426-432. J. Neurosci. Res. 1998, 54, 123. [Google Scholar] [CrossRef]
- Wu, J.; Jeong, H.K.; Bulin, S.E.; Kwon, S.W.; Park, J.H.; Bezprozvanny, I. Ginsenosides protect striatal neurons in a cellular model of Huntington’s disease. J. Neurosci. Res. 2009, 87, 1904–1912. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.-L.; Wang, Q.-Z.; Sun, L.-M.; Li, X.-M.; Deng, J.-M.; Li, L.-F.; Zhang, J.; Xu, R.; Ma, S.-P. Asiaticoside: Attenuation of neurotoxicity induced by MPTP in a rat model of Parkinsonism via maintaining redox balance and up-regulating the ratio of Bcl-2/Bax. Pharmacol. Biochem. Behav. 2012, 100, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-C.; Zhu, Y.-G.; Zhu, L.-A.; Huang, C.; Chen, Y.; Chen, L.-M.; Fang, F.; Zhou, Y.-C.; Zhao, C.-H. Ginsenoside Rg1 attenuates dopamine-induced apoptosis in PC12 cells by suppressing oxidative stress. Eur. J. Pharmacol. 2003, 473, 1–7. [Google Scholar] [CrossRef]
- López, M.V.N.; Cuadrado, M.P.G.-S.; Ruiz-Poveda, O.M.P.; Del Fresno, A.M.V.; Accame, M.E.C. Neuroprotective effect of individual ginsenosides on astrocytes primary culture. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2007, 1770, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.-H.; Han, H.; Hu, X.-D.; Shi, L.-L. Protective effect of ginsenoside Rb1 on β-amyloid protein (1-42)-induced neurotoxicity in cortical neurons. Neurol. Res. 2009, 31, 663–667. [Google Scholar] [CrossRef]
- Shin, E.-J.; Koh, Y.H.; Kim, A.-Y.; Nah, S.-Y.; Jeong, J.H.; Chae, J.-S.; Kim, S.C.; Yen, T.P.H.; Yoon, H.-J.; Kim, W.-K. Ginsenosides attenuate kainic acid-induced synaptosomal oxidative stress via stimulation of adenosine A2A receptors in rat hippocampus. Behav. Brain Res. 2009, 197, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.-J.; Nabeshima, T.; Suh, H.-W.; Jhoo, W.-K.; Oh, K.-W.; Lim, Y.-K.; Kim, D.S.; Choi, K.H.; Kim, H.-C. Ginsenosides attenuate methamphetamine-induced behavioral side effects in mice via activation of adenosine A2A receptors: Possible involvements of the striatal reduction in AP-1 DNA binding activity and proenkephalin gene expression. Behav. Brain Res. 2005, 158, 143–157. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Yan, X.; Qin, K.; Shi, M.; Lin, T.; Zhu, Y.; Kang, T.; Zhao, G. Protective effects of ginsenoside Rd against okadaic acid-induced neurotoxicity in vivo and in vitro. J. Ethnopharmacol. 2011, 138, 135–141. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abduljawad, A.A.; Elawad, M.A.; Elkhalifa, M.E.M.; Ahmed, A.; Hamdoon, A.A.E.; Salim, L.H.M.; Ashraf, M.; Ayaz, M.; Hassan, S.S.u.; Bungau, S. Alzheimer’s Disease as a Major Public Health Concern: Role of Dietary Saponins in Mitigating Neurodegenerative Disorders and Their Underlying Mechanisms. Molecules 2022, 27, 6804. https://doi.org/10.3390/molecules27206804
Abduljawad AA, Elawad MA, Elkhalifa MEM, Ahmed A, Hamdoon AAE, Salim LHM, Ashraf M, Ayaz M, Hassan SSu, Bungau S. Alzheimer’s Disease as a Major Public Health Concern: Role of Dietary Saponins in Mitigating Neurodegenerative Disorders and Their Underlying Mechanisms. Molecules. 2022; 27(20):6804. https://doi.org/10.3390/molecules27206804
Chicago/Turabian StyleAbduljawad, Asaad A., Mohammed Ahmed Elawad, Modawy Elnour Modawy Elkhalifa, Alshebli Ahmed, Alashary Adam Eisa Hamdoon, Liga Hasan Mohammed Salim, Muhammad Ashraf, Muhammad Ayaz, Syed Shams ul Hassan, and Simona Bungau. 2022. "Alzheimer’s Disease as a Major Public Health Concern: Role of Dietary Saponins in Mitigating Neurodegenerative Disorders and Their Underlying Mechanisms" Molecules 27, no. 20: 6804. https://doi.org/10.3390/molecules27206804
APA StyleAbduljawad, A. A., Elawad, M. A., Elkhalifa, M. E. M., Ahmed, A., Hamdoon, A. A. E., Salim, L. H. M., Ashraf, M., Ayaz, M., Hassan, S. S. u., & Bungau, S. (2022). Alzheimer’s Disease as a Major Public Health Concern: Role of Dietary Saponins in Mitigating Neurodegenerative Disorders and Their Underlying Mechanisms. Molecules, 27(20), 6804. https://doi.org/10.3390/molecules27206804