

Antioxidant Properties of Hemp Proteins: From Functional Food to Phytotherapy and Beyond





Abstract
:1. Introduction
2. Structure-Modifying Techniques
2.1. pH-Shift
2.2. Enzymatic Hydrolysis
3. Assessments of the Antioxidant Ability
3.1. Reactive Oxygen Species Scavenging Ability
3.2. Metal Ion Chelating Capacity
3.3. Assessments of Antioxidant Enzymatic and Non-Enzymatic Activities
4. Structure-Activity Relationship of Antioxidant Peptides
4.1. Molecular Weight
4.2. Composition of Amino Acids
4.3. Sequence of Amino Acids
5. Pre-Clinical Studies on Hemp Seed and Pathogenesis-Related Molecular Mechanisms
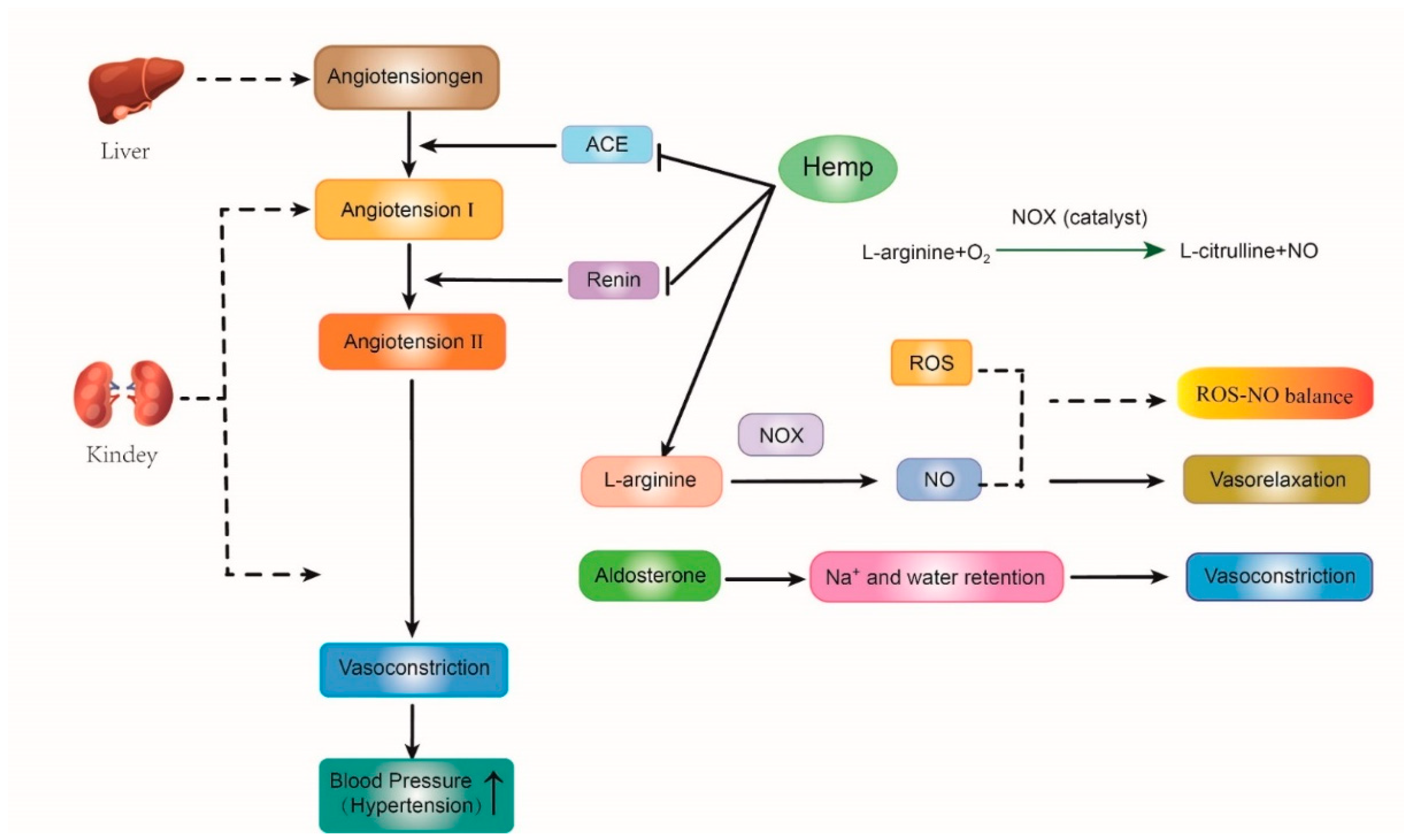
5.1. Anti-Hypertension Effect
5.2. Anti-Inflammatory Effect
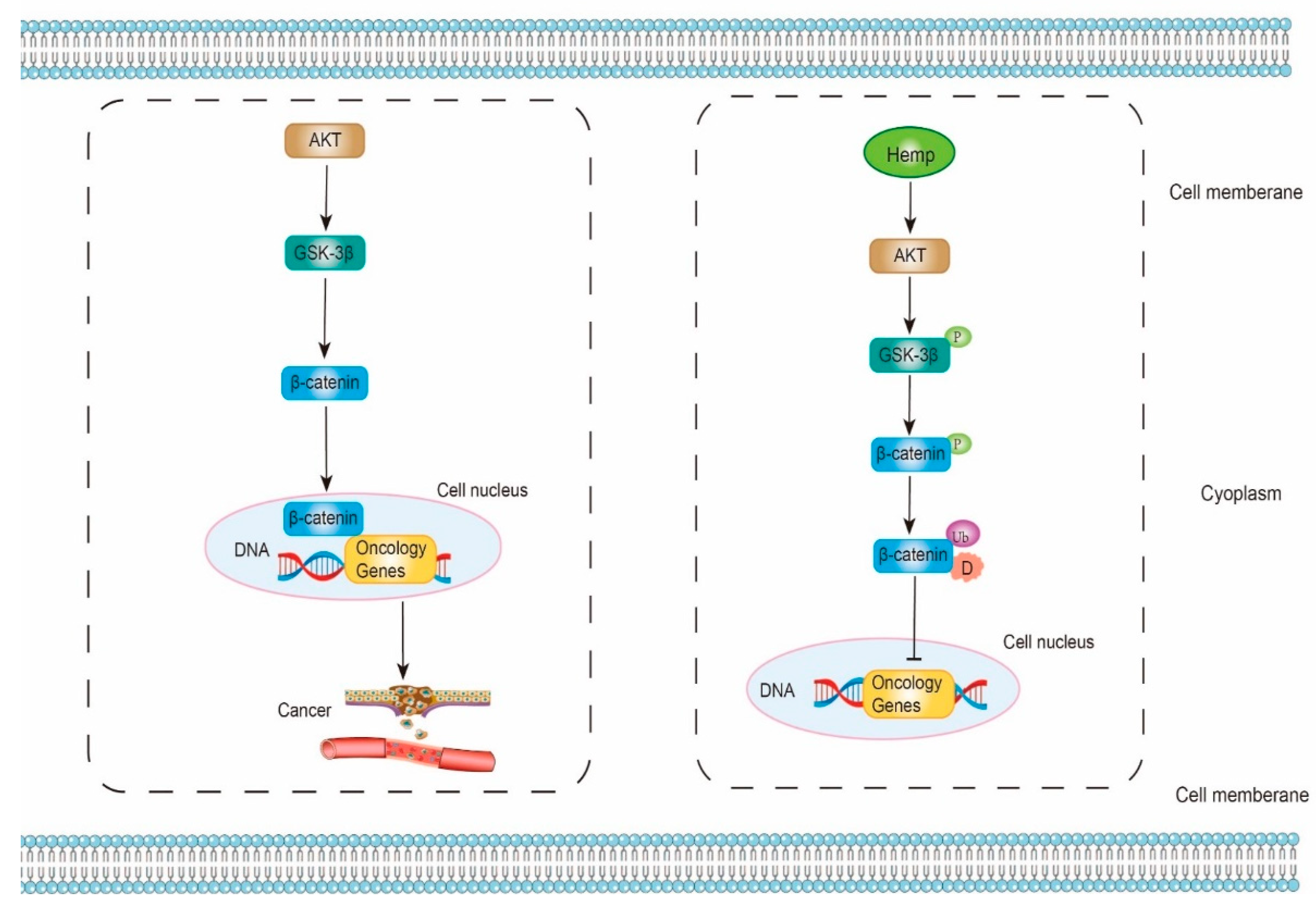
5.3. Anti-Cancer Effect
6. Hemp Seed Protein Product and Usage
7. Potential Hazard
8. Conclusions and Future
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gude, K.M.; Rajashekar, C.B.; Cunningham, B.; Kang, Q.; Wang, W.; Lee, M.; Rivard, C.L.; Pliakoni, E.D. Effect of High Tunnel Coverings on Antioxidants of Breaker and Light Red Tomatoes at Harvest and during Ripening. Agronomy 2020, 10, 1639. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Xu, Y.; Isenberg, J.S.; Bachowski, S.; Kolaja, K.L.; Jiang, J.; Stevenson, D.E.; Walborg, E.F. The role of oxidative stress in chemical carcinogenesis. Environ. Health Perspect. 1998, 106, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, A.; Priyadarsini, K.I. Free radicals, oxidative stress and importance of antioxidants in human health. J. Med. Allied Sci. 2011, 1, 53–60. [Google Scholar]
- Ramarathnam, N.; Osawa, T.; Ochi, H.; Kawakishi, S. The contribution of plant food antioxidants to human health. Trends Food Sci. Technol. 1995, 6, 75–82. [Google Scholar] [CrossRef]
- Pan, Y.; Li, H.; Zheng, S.-L.; Zhang, B.; Deng, Z.-Y. Implication of the Significance of Dietary Compatibility: Based on the Antioxidant and Anti-Inflammatory Interactions with Different Ratios of Hydrophilic and Lipophilic Antioxidants among Four Daily Agricultural Crops. J. Agric. Food Chem. 2018, 66, 7461–7474. [Google Scholar] [CrossRef]
- Dias, T.R.; Martin-Hidalgo, D.; Silva, B.M.; Oliveira, P.F.; Alves, M.G. Endogenous and Exogenous Antioxidants as a Tool to Ameliorate Male Infertility Induced by Reactive Oxygen Species. Antioxidants Redox Signal. 2020, 33, 767–785. [Google Scholar] [CrossRef]
- Wang, X.S.; Tang, C.H.; Yang, X.Q.; Gao, W.R. Characterization; amino acid composition and in vitro digestibility of hemp (Cannabis sativa L.) proteins. Food Chem. 2008, 107, 11–18. [Google Scholar] [CrossRef]
- Wang, W.; Ayella, A.; Jiang, Y.; Ouyang, P.; Qu, H. Wheat Lignans: Promising Cancer Preventive Agents; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 264–272. [Google Scholar]
- Wang, W.; Goodman, M.T. Antioxidant property of dietary phenolic agents in a human LDL-oxidation ex vivo model: Interaction of protein binding activity. Nutr. Res. 1999, 19, 191–202. [Google Scholar] [CrossRef]
- Lai, L.-S.; Chou, S.-T.; Chao, W.-W. Studies on the Antioxidative Activities of Hsian-tsao (Mesona procumbens Hemsl) Leaf Gum. J. Agric. Food Chem. 2001, 49, 963–968. [Google Scholar] [CrossRef]
- Tachakittirungrod, S.; Okonogi, S.; Chowwanapoonpohn, S. Study on antioxidant activity of certain plants in Thailand: Mechanism of antioxidant action of guava leaf extract. Food Chem. 2007, 103, 381–388. [Google Scholar] [CrossRef]
- Gülçin, I. The antioxidant and radical scavenging activities of black pepper (Piper nigrum) seeds. Int. J. Food Sci. Nutr. 2005, 56, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant and antiradical activities of l-carnitine. Life Sci. 2006, 78, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant activity of caffeic acid (3,4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef]
- Mizobuchi, M.; Ishidoh, K.; Kamemura, N. A comparison of cell death mechanisms of antioxidants, butylated hydroxyanisole and butylated hydroxytoluene. Drug Chem. Toxicol. 2022, 45, 1899–1906. [Google Scholar] [CrossRef]
- Moure, A.; Cruz, J.M.; Franco, D.; Domínguez, J.; Sineiro, J.; Domínguez, H.; Núñez, M.J.; Parajó, J. Natural antioxidants from residual sources. Food Chem. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Zengin, G.; Menghini, L.; Di Sotto, A.; Mancinelli, R.; Sisto, F.; Carradori, S.; Cesa, S.; Fraschetti, C.; Filippi, A.; Angiolella, L.; et al. Chromatographic Analyses, In Vitro Biological Activities, and Cytotoxicity of Cannabis sativa L. Essential Oil: A Multidisciplinary Study. Molecules 2018, 23, 3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schluttenhofer, C.; Yuan, L. Challenges towards Revitalizing Hemp: A Multifaceted Crop. Trends Plant Sci. 2017, 22, 917–929. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Hemp. History of Hemp in the U.S. Ministry of Hemp. Available online: https://ministryofhemp.com/hemp/history/ (accessed on 13 November 2022).
- Kleinhenz, M.D.; Magnin, G.; Lin, Z.; Griffin, J.; Kleinhenz, K.E.; Montgomery, S.; Curtis, A.; Martin, M.; Coetzee, J.F. Plasma concentrations of eleven cannabinoids in cattle following oral administration of industrial hemp (Cannabis sativa). Sci. Rep. 2020, 10, 12753–12760. [Google Scholar] [CrossRef]
- Kleinhenz, M.D.; Magnin, G.; Ensley, S.M.; Griffin, J.J.; Goeser, J.; Lynch, E.; Coetzee, J.F. Nutrient concentrations, digestibility, and cannabinoid concentrations of industrial hemp plant components. Appl. Anim. Sci. 2020, 36, 489–494. [Google Scholar] [CrossRef]
- Wagner, J.R.; Sorgentini, D.A.; Añón, M.C. Thermal and Electrophoretic Behavior, Hydrophobicity, and Some Functional Properties of Acid-Treated Soy Isolates. J. Agric. Food Chem. 1996, 44, 1881–1889. [Google Scholar] [CrossRef]
- Jiang, J.; Xiong, Y.L. Extreme pH treatments enhance the structure-reinforcement role of soy protein isolate and its emulsions in pork myofibrillar protein gels in the presence of microbial transglutaminase. Meat Sci. 2013, 93, 469–476. [Google Scholar] [CrossRef]
- Moure, A.; Domínguez, H.; Parajó, J.C. Antioxidant properties of ultrafiltration-recovered soy protein fractions from industrial effluents and their hydrolysates. Process Biochem. 2006, 41, 447–456. [Google Scholar] [CrossRef]
- Kim, S.-K.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Mannheim, A.; Cheryan, M. Enzyme-modified proteins from corn gluten meal: Preparation and functional properties. J. Am. Oil Chem. Soc. 1992, 69, 1163–1169. [Google Scholar] [CrossRef]
- Chen, C.; Chi, Y.J.; Zhao, M.Y.; Lv, L. Purification and identification of antioxidant peptides from egg white protein hydrolysate. Amino Acids 2012, 43, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Adler-Nissen, J. Enzymatic hydrolysis of soy protein for nutritional fortification of low pH food. Ann. Nutr. Aliment. 1978, 32, 205–216. [Google Scholar] [PubMed]
- Molnár-Perl, I. Advancement in the derivatizations of the amino groups with the o-phthaldehyde-thiol and with the 9-fluorenylmethyloxycarbonyl chloride reagents. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 1241–1269. [Google Scholar] [CrossRef]
- Aiello, G.; Lammi, C.; Boschin, G.; Zanoni, C.; Arnoldi, A. Exploration of Potentially Bioactive Peptides Generated from the Enzymatic Hydrolysis of Hempseed Proteins. J. Agric. Food Chem. 2017, 65, 10174–10184. [Google Scholar] [CrossRef]
- Tang, C.-H.; Wang, X.-S.; Yang, X.-Q. Enzymatic hydrolysis of hemp (Cannabis sativa L.) protein isolate by various proteases and antioxidant properties of the resulting hydrolysates. Food Chem. 2009, 114, 1484–1490. [Google Scholar] [CrossRef]
- Zou, T.-B.; He, T.-P.; Li, H.-B.; Tang, H.-W.; Xia, E.-Q. The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.; Li-Chan, E.C. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Zhang, M.; Mu, T.-H.; Sun, M.-J. Purification and identification of antioxidant peptides from sweet potato protein hydrolysates by Alcalase. J. Funct. Foods 2014, 7, 191–200. [Google Scholar] [CrossRef]
- Osakwe, O.N.; Siegel, A. A Novel Standardized Oxygen Radical Absorbance Assay for Evaluating Antioxidant Natural Products. J. AOAC Int. 2013, 96, 1365–1371. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Cao, G.; Alessio, H.M.; Cutler, R.G. Oxygen-radical absorbance capacity assay for antioxidants. Free Radic. Biol. Med. 1993, 14, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Wolfram Notebook. Simulating a Minimal ORAC Kinetic System. Available online: https://www.wolframcloud.com/objects/demonstrations/SimulatingAMinimalORACKineticSystem-source.nb (accessed on 13 November 2022).
- Lu, R.-R.; Qian, P.; Sun, Z.; Zhou, X.-H.; Chen, T.-P.; He, J.-F.; Zhang, H.; Wu, J. Hempseed protein derived antioxidative peptides: Purification, identification and protection from hydrogen peroxide-induced apoptosis in PC12 cells. Food Chem. 2010, 123, 1210–1218. [Google Scholar] [CrossRef]
- Frassinetti, S.; Moccia, E.; Caltavuturo, L.; Gabriele, M.; Longo, V.; Bellani, L.; Giorgi, G.; Giorgetti, L. Nutraceutical potential of hemp (Cannabis sativa L.) seeds and sprouts. Food Chem. 2018, 262, 56–66. [Google Scholar] [CrossRef]
- Radočaj, O.; Dimić, E.; Tsao, R. Effects of Hemp (Cannabis sativa L.) Seed Oil Press-Cake and Decaffeinated Green Tea Leaves (Camellia sinensis) on Functional Characteristics of Gluten-Free Crackers. J. Food Sci. 2014, 79, C318–C325. [Google Scholar] [CrossRef]
- Megías, C.; Pedroche, J.; Yust, M.M.; Girón-Calle, J.; Alaiz, M.; Millán, F.; Vioque, J. Production of copper-chelating peptides after hydrolysis of sunflower proteins with pepsin and pancreatin. Food Sci. Technol. 2008, 41, 1973–1977. [Google Scholar] [CrossRef]
- Gulcin, I.; Alwasel, S.H. Metal Ions, Metal Chelators and Metal Chelating Assay as Antioxidant Method. Processes 2022, 10, 132. [Google Scholar] [CrossRef]
- Girgih, A.T.; Udenigwe, C.; Aluko, R.E. In Vitro Antioxidant Properties of Hemp Seed (Cannabis sativa L.) Protein Hydrolysate Fractions. J. Am. Oil Chem. Soc. 2010, 88, 381–389. [Google Scholar] [CrossRef]
- Wang, X.S.; Tang, C.H.; Chen, L.; Yang, X.Q. Characterization and Antioxidant Properties of Hemp Protein Hydrolysates Obtained with Neutrase. Food Technol. Biotechnol. 2009, 47, 428–434. Available online: https://hrcak.srce.hr/43902 (accessed on 13 November 2022).
- Borneo, R.; León, A.; Aguirre, A.; Ribotta, P.; Cantero, J. Antioxidant capacity of medicinal plants from the Province of Córdoba (Argentina) and their in vitro testing in a model food system. Food Chem. 2009, 112, 664–670. [Google Scholar] [CrossRef]
- Soares, A.A.; De Souza, C.G.M.; Daniel, F.M.; Ferrari, G.P.; Da Costa, S.M.G.; Peralta, R.M. Antioxidant activity and total phenolic content of Agaricus brasiliensis (Agaricus blazei Murril) in two stages of maturity. Food Chem. 2009, 112, 775–781. [Google Scholar] [CrossRef]
- Hong, S.; Sowndhararajan, K.; Joo, T.; Lim, C.; Cho, H.; Kim, S.; Kim, G.-Y.; Jhoo, J.-W. Ethanol and supercritical fluid extracts of hemp seed (Cannabis sativa L.) increase gene expression of antioxidant enzymes in HepG2 cells Antioxidant enzyme Cannabis sativa Hemp seed HepG2 cells Catalase Superoxide dismutase. Asian Pac. J. Reprod. 2015, 4, 147–152. [Google Scholar] [CrossRef]
- Girgih, A.T.; Alashi, A.M.; He, R.; Malomo, S.A.; Raj, P.D.; Netticadan, T.; Aluko, R.E. A Novel Hemp Seed Meal Protein Hydrolysate Reduces Oxidative Stress Factors in Spontaneously Hypertensive Rats. Nutrients 2014, 6, 5652–5666. [Google Scholar] [CrossRef] [Green Version]
- Kubiliene, A.; Mickute, K.; Baranauskaite, J.; Marksa, M.; Liekis, A.; Sadauskiene, I. The Effects of Cannabis sativa L. Extract on Oxidative Stress Markers In Vivo. Life 2021, 11, 647. [Google Scholar] [CrossRef]
- Wu, H.-Y.; Jan, T.-R. Cannabidiol hydroxyquinone-induced apoptosis of splenocytes is mediated predominantly by thiol depletion. Toxicol. Lett. 2010, 195, 68–74. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Flanagan, J.; Deemer, E.K.; Prior, R.L.; Huang, D. Novel Fluorometric Assay for Hydroxyl Radical Prevention Capacity Using Fluorescein as the Probe. J. Agric. Food Chem. 2002, 50, 2772–2777. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant Activity of Proteins and Peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A. A review of fish-derived antioxidant and antimicrobial peptides: Their production, assessment, and applications. Peptides 2012, 33, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Gong, Y.-D.; Li, Z.-R.; Yu, D.; Chi, C.-F.; Ma, J.-Y. Isolation and characterisation of five novel antioxidant peptides from ethanol-soluble proteins hydrolysate of spotless smoothhound (Mustelus griseus) muscle. J. Funct. Foods 2014, 6, 176–185. [Google Scholar] [CrossRef]
- Ren, Y.; Wu, H.; Li, X.; Lai, F.; Xiao, X. Purification and characterization of high antioxidant peptides from duck egg white protein hydrolysates. Biochem. Biophys. Res. Commun. 2014, 452, 888–894. [Google Scholar] [CrossRef]
- Kitrytė, V.; Bagdonaitė, D.; Rimantas Venskutonis, P. Biorefining of industrial hemp (Cannabis sativa L.) threshing residues into cannabinoid and antioxidant fractions by supercritical carbon dioxide; pressurized liquid and enzyme-assisted extractions. Food Chem. 2018, 267, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Chen, F.; Wang, X.; Ji, B.; Wu, Y. Isolation and identification of antioxidative peptides from porcine collagen hydrolysate by consecutive chromatography and electrospray ionization mass spectrometry. Food Chem. 2007, 102, 1135–1143. [Google Scholar] [CrossRef]
- Girgih, A.T.; Udenigwe, C.C.; Aluko, R.E. Reverse-phase HPLC Separation of Hemp Seed (Cannabis sativa L.) Protein Hydrolysate Produced Peptide Fractions with Enhanced Antioxidant Capacity. Mater. Veg. 2013, 68, 39–46. [Google Scholar] [CrossRef]
- Li, Y.-W.; Li, B.; He, J.; Qian, P. Structure-activity relationship study of antioxidative peptides by QSAR modeling: The amino acid next to C-terminus affects the activity. J. Pept. Sci. 2011, 17, 454–462. [Google Scholar] [CrossRef]
- Liu, P.; Long, W. Current Mathematical Methods Used in QSAR/QSPR Studies. Int. J. Mol. Sci. 2009, 10, 1978–1998. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Lin, Y.; Ding, Y.; Wang, Y.; Cheng, X.; Lin, Z. QSAR study on angiotensin-converting enzyme inhibitor oligopeptides based on a novel set of sequence information descriptors. J. Mol. Model. 2011, 17, 1599–1606. [Google Scholar] [CrossRef]
- Liang, G.; Liu, Y.; Shi, B.; Zhao, J.; Zheng, J. An Index for Characterization of Natural and Non-Natural Amino Acids for Peptidomimetics. PLoS ONE 2013, 8, e67844. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Li, Y. Review and perspective on bioactive peptides: A roadmap for research, development, and future opportunities. J. Agric. Food Res. 2022, 9, 100353. [Google Scholar] [CrossRef]
- Du, Z.; Wang, D.; Li, Y. Comprehensive Evaluation and Comparison of Machine Learning Methods in QSAR Modeling of Antioxidant Tripeptides. ACS Omega 2022, 7, 25760–25771. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Muramoto, K.; Yamauchi, F.; Nokihara, K. Antioxidant Activity of Designed Peptides Based on the Antioxidative Peptide Isolated from Digests of a Soybean Protein. J. Agric. Food Chem. 1996, 44, 2619–2623. [Google Scholar] [CrossRef]
- Erdmann, K.; Cheung, B.W.; Schröder, H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef]
- Möller, N.P.; Scholz-Ahrens, K.E.; Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: Indication for health effects. Eur. J. Nutr. 2008, 47, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Borges, R.S.; Júnior, J.B.; Viana, R.B.; Baetas, A.C.; Orestes, E.; Andrade, M.A.; Honório, K.M.; Da Silva, A.B.F. Understanding the Molecular Aspects of Tetrahydrocannabinol and Cannabidiol as Antioxidants. Molecules 2013, 18, 12663–12674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, R.; Honda, R.; Kanome, M.; Hagiwara, A.; Matsuda, Y.; Togitani, T.; Ikemoto, N.; Terashima, M. Designing antioxidant peptides based on the antioxidant properties of the amino acid side-chains. Food Chem. 2018, 245, 750–755. [Google Scholar] [CrossRef]
- Aguirre-Cruz, G.; León-López, A.; Cruz-Gómez, V.; Jiménez-Alvarado, R.; Aguirre-Álvarez, G. Collagen Hydrolysates for Skin Protection: Oral Administration and Topical Formulation. Antioxidants 2020, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- The Lecturio Medical Concept Library. Basics of Amino Acids. Available online: https://www.lecturio.com/concepts/basics-of-amino-acids/ (accessed on 13 November 2022).
- Chi, C.-F.; Hu, F.-Y.; Wang, B.; Li, Z.-R.; Luo, H.-Y. Influence of Amino Acid Compositions and Peptide Profiles on Antioxidant Capacities of Two Protein Hydrolysates from Skipjack Tuna (Katsuwonus pelamis) Dark Muscle. Mar. Drugs 2015, 13, 2580–2601. [Google Scholar] [CrossRef]
- Chi, C.F.; Wang, B.; Wang, Y.M.; Zhang, B.; Deng, S.J. Isolation and characterization of three antioxidant peptides from protein hydrolysate of bluefin leatherjacket (Navodon septentrionalis) heads. J. Funct. Foods 2015, 12, 1–10. [Google Scholar] [CrossRef]
- Tian, M.; Fang, B.; Jiang, L.; Guo, H.; Cui, J.; Ren, F. Structure-activity relationship of a series of antioxidant tripeptides derived from β-Lactoglobulin using QSAR modeling. Dairy Sci. Technol. 2015, 95, 451–463. [Google Scholar] [CrossRef]
- Rodriguez-Martin, N.M.; Toscano, R.; Villanueva, A.; Pedroche, J.; Millan, F.; la Paz, S.M.-D.; Millan-Linares, M.C. Neuroprotective protein hydrolysates from hemp (Cannabis sativa L.) seeds. Food Funct. 2019, 10, 6732–6739. [Google Scholar] [CrossRef] [PubMed]
- Teh, S.S.; Bekhit, A.E.D.; Carne, A.; Birch, J. Effect of the defatting process, acid and alkali extraction on the physicochemical and functional properties of hemp, flax and canola seed cake protein isolates. J. Food Meas. Charact. 2014, 8, 92–104. [Google Scholar] [CrossRef]
- Bougatef, A.; Nedjar-Arroume, N.; Manni, L.; Ravallec, R.; Barkia, A.; Guillochon, D.; Nasri, M. Purification and identification of novel antioxidant peptides from enzymatic hydrolysates of sardinelle (Sardinella aurita) by-products proteins. Food Chem. 2010, 118, 559–565. [Google Scholar] [CrossRef]
- Xu, L.; Li Xi Wu Xi Wang, H.; Hou Ru Huang Yi Zhang, X. Preparation and structural characterization of a new corn antioxidant Peptide. Chem. Res. Chin. Univ. 2004, 25, 466–469. [Google Scholar]
- Bamdad, F.; Ahmed, S.; Chen, L. Specifically designed peptide structures effectively suppressed oxidative reactions in chemical and cellular systems. J. Funct. Foods 2015, 18, 35–46. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Ren, X.; Zhang, X.; Wu, Z.; Liu, L. The positive correlation of antioxidant activity and prebiotic effect about naked oat phenolic compounds. Food Chem. 2023, 402, 134231. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Q.; Zhou, Y.; Zhang, Y.; Ren, Y.; Liu, L. Characterization and determination of bovine immunoglobulin G subtypes in milk and dairy products by UPLC-MS. Food Chem. 2022, 390, 133170. [Google Scholar] [CrossRef]
- Chen, Q.; Li, Y.; Dong, L.; Shi, R.; Wu, Z.; Liu, L.; Zhang, J.; Wu, Z.; Pan, D. Quantitative determination of Nε-(carboxymethyl)lysine in sterilized milk by isotope dilution UPLC-MS/MS method without derivatization and ion pair reagents. Food Chem. 2022, 385, 132697. [Google Scholar] [CrossRef]
- Iftikhar, A.; Zafar, U.; Ahmed, W.; Shabbir, M.A.; Sameen, A.; Sahar, A.; Bhat, Z.F.; Kowalczewski, P.; Jarzębski, M.; Aadil, R.M. Applications of Cannabis sativa L. in Food and Its Therapeutic Potential: From a Prohibited Drug to a Nutritional Supplement. Molecules 2021, 26, 7699. [Google Scholar] [CrossRef]
- Karabulut, G.; Feng, H.; Yemiş, O. Physicochemical and Antioxidant Properties of Industrial Hemp Seed Protein Isolate Treated by High-Intensity Ultrasound. Plant Foods Hum Nutr. 2022, 77, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Ukeda, H. Isolation an octaperptide which possesses active oxygen scavenging activity from peptic digest of sardine muscle. Nippon. Suisan Gakkaishi 1999, 65, 1096–1099. [Google Scholar] [CrossRef]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar] [CrossRef]
- Guo, H.; Kouzuma, Y.; Yonekura, M. Structures and properties of antioxidative peptides derived from royal jelly protein. Food Chem. 2009, 113, 238–245. [Google Scholar] [CrossRef]
- Dávalos, A.; Miguel, M.; Bartolomé, B.; López-Fandiño, R. Antioxidant Activity of Peptides Derived from Egg White Proteins by Enzymatic Hydrolysis. J. Food Prot. 2004, 67, 1939–1944. [Google Scholar] [CrossRef] [PubMed]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant Activity of Peptides Obtained from Porcine Myofibrillar Proteins by Protease Treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Wang, S.; Meckling, K.A.; Marcone, M.F.; Kakuda, Y.; Tsao, R. Can phytochemical antioxidant rich foods act as anti-cancer agents? Food Res. Int. 2011, 44, 2545–2554. [Google Scholar] [CrossRef]
- Zuardi, A.W.; Antonio, C.; Zuardi, W.; Candido, A.; Lima, P.; Recreio, J.D. Cannabidiol, from an inactive cannabinoid to a drug with wide spectrum of action. Rev. Bras. Psiquiatr. 2008, 30, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Fouad, A.A.; Albuali, W.H.; Mulhim, A.S.; Jresat, I. Cardioprotective effect of cannabidiol in rats exposed to doxorubicin toxicity. Environ. Toxicol. Pharmacol. 2013, 36, 347–357. [Google Scholar] [CrossRef]
- Chagas, M.H.N.; Zuardi, A.W.; Tumas, V.; Pena-Pereira, M.A.; Sobreira, E.T.; Bergamaschi, M.M.; Dos Santos, A.C.; Teixeira, A.L.; Hallak, J.E.C.; Crippa, J.A.S. Effects of cannabidiol in the treatment of patients with Parkinson’s disease: An exploratory double-blind trial. J. Psychopharmacol. 2014, 28, 1088–1098. [Google Scholar] [CrossRef]
- Xu, Y.; Li, J.; Zhao, J.; Wang, W.; Griffin, J.; Li, Y.; Bean, S.; Tilley, M.; Wang, D. Hempseed as a nutritious and healthy human food or animal feed source: A review. Int. J. Food Sci. Technol. 2020, 56, 530–543. [Google Scholar] [CrossRef]
- Aluko, R.E. Antihypertensive Peptides from Food Proteins. Annu. Rev. Food Sci. Technol. 2015, 6, 235–262. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Malomo, S.; Offengenden, M.; Wu, J.; Aluko, R.E. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Tain, Y.-L.; Hsu, C.-N. Oxidative Stress-Induced Hypertension of Developmental Origins: Preventive Aspects of Antioxidant Therapy. Antioxidants 2022, 11, 511. [Google Scholar] [CrossRef] [PubMed]
- Samaei, S.P.; Martini, S.; Tagliazucchi, D.; Gianotti, A.; Babini, E. Antioxidant and Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides Obtained from Alcalase Protein Hydrolysate Fractions of Hemp (Cannabis sativa L.) Bran. J. Agric. Food Chem. 2021, 69, 9220–9228. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; Udenigwe, C.C.; Li, H.; Adebiyi, A.P.; Aluko, R.E. Kinetics of Enzyme Inhibition and Antihypertensive Effects of Hemp Seed (Cannabis sativa L.) Protein Hydrolysates. J. Am. Oil Chem. Soc. 2011, 88, 1767–1774. [Google Scholar] [CrossRef]
- Surh, Y.-J.; Chun, K.-S.; Cha, H.-H.; Han, S.S.; Keum, Y.-S.; Park, K.-K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutat. Res. Mol. Mech. Mutagen. 2001, 480, 243–268. [Google Scholar] [CrossRef]
- Libby, P. Inflammatory Mechanisms: The Molecular Basis of Inflammation and Disease. Nutr. Rev. 2007, 65, S140–S146. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-κB, the first quarter-century: Remarkable progress and outstanding questions. Genes Dev. 2012, 26, 203–234. [Google Scholar] [CrossRef] [Green Version]
- Smale, S.T. Dimer-specific regulatory mechanisms within the NF-κB family of transcription factors. Immunol. Rev. 2012, 246, 193–204. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Shared Principles in NF-κB Signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Rasheed, Z.; Ahsan, H. Biochemical and cellular toxicology of peroxynitrite: Implications in cell death and autoimmune phenomenon. Immunopharmacol. Immunotoxicol. 2009, 31, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.G.; Oakley, F. Nuclear factor-κB1: Regulation and function. Int. J. Biochem. Cell Biol. 2008, 40, 1425–1430. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Park, S.H.; Han, J.H.; Hong, Y.K.; Hwang, S.; Lee, S.; Kim, D.; Han, S.Y.; Kim, E.S.; Cho, K.S. The effects of hempseed meal intake and linoleic acid on drosophila models of neurodegenerative diseases and hypercholesterolemia. Mol. Cells 2011, 31, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Yan, X.; Bobrovskaya, L.; Ji, M.; Yuan, H.; Lou, H.; Fan, P. Anti-neuroinflammatory effects of grossamide from hemp seed via suppression of TLR-4-mediated NF-κB signaling pathways in lipopolysaccharide-stimulated BV2 microglia cells. Mol. Cell Biochem. 2017, 428, 129–137. [Google Scholar] [CrossRef]
- Wen, Z.-S.; Xue, R.; Du, M.; Tang, Z.; Xiang, X.-W.; Zheng, B.; Qu, Y.-L. Hemp seed polysaccharides protect intestinal epithelial cells from hydrogen peroxide-induced oxidative stress. Int. J. Biol. Macromol. 2019, 135, 203–211. [Google Scholar] [CrossRef]
- Lee, J.-M.; Johnson, J.A. An important role of Nrf2-ARE pathway in the cellular defense mechanism. BMB Rep. 2004, 37, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-F.; Kuo, H.-P.; Liu, M.; Chou, C.-K.; Xia, W.; Du, Y.; Shen, J.; Chen, C.-T.; Huo, L.; Hsu, M.-C.; et al. KEAP1 E3 Ligase-Mediated Downregulation of NF-κB Signaling by Targeting IKKβ. Mol. Cell 2009, 36, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Zhao, L.; Zhao, Q.; Zhao, Y.; Sun, Y.; Zhang, Y.; Miao, H.; You, Q.-D.; Hu, R.; Guo, Q.-L. NF-κB and Nrf2 signaling pathways contribute to wogonin-mediated inhibition of inflammation-associated colorectal carcinogenesis. Cell Death Dis. 2014, 5, e1283. [Google Scholar] [CrossRef] [Green Version]
- Awuh, J.A.; Haug, M.; Mildenberger, J.; Marstad, A.; Do, C.P.N.; Louet, C.; Stenvik, J.; Steigedal, M.; Damås, J.K.; Halaas, Ø.; et al. Keap1 regulates inflammatory signaling in Mycobacterium avium -infected human macrophages. Proc. Natl. Acad. Sci. USA 2015, 112, E4272–E4280. [Google Scholar] [CrossRef] [Green Version]
- Jeong, W.-S.; Jun, M.; Kong, A.-N.T. Nrf2: A Potential Molecular Target for Cancer Chemoprevention by Natural Compounds. Antioxidants Redox. Signal. 2006, 8, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; King, B.; Ewert, E.; Su, X.; Mardiyati, N.; Zhao, Z.; Wang, W. Exercise Activates p53 and Negatively Regulates IGF,1 Pathway in epidermis within a skin cancer model. PLoS ONE 2016, 11, e0160939. Available online: http://hdl.handle.net/2097/34600 (accessed on 13 November 2022). [CrossRef] [PubMed] [Green Version]
- Wang, S.; Luo, Q.; Fan, P. Cannabisin F from Hemp (Cannabis sativa) Seed Suppresses Lipopolysaccharide-Induced Inflammatory Responses in BV2 Microglia as SIRT1 Modulator. Int. J. Mol. Sci. 2019, 20, 507. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, S.; Hsieh, C.-M.; Maemura, K.; Layne, M.D.; Yet, S.-F.; Lee, K.-H.; Matsui, T.; Rosenzweig, A.; Taylor, W.G.; Rubin, J.S.; et al. Akt Participation in the Wnt Signaling Pathway through Dishevelled. J. Biol. Chem. 2001, 276, 17479–17483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Song, H.; Yang, P.; Dai, X.; Li, Y.; Wang, L.; Du, J.; Pan, K.; Zhang, T. Silencing dishevelled-1 sensitizes paclitaxel-resistant human ovarian cancer cells via AKT/GSK-3β/β-catenin signalling. Cell Prolif. 2015, 48, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Feng, C.-C.; Chu, S.-J.; Zhang, R.; Lu, Y.-M.; Zhu, J.-S.; Zhang, J. Toosendanin inhibits growth and induces apoptosis in colorectal cancer cells through suppression of AKT/GSK-3β/β-catenin pathway. Int. J. Oncol. 2015, 47, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Miao, H.-C.; Li, W.-J.; Sun, Y.; Huang, S.-L.; Li, Z.-Y.; Guo, Q.-L. LW-213 induces G2/M cell cycle arrest through AKT/GSK3β/β-catenin signaling pathway in human breast cancer cells. Mol. Carcinog. 2016, 55, 778–792. [Google Scholar] [CrossRef]
- Ozman, Z.; Iptec, B.O.; Sahin, E.; Eskiler, G.G.; Ozkan, A.D.; Kaleli, S. Regulation of valproic acid induced EMT by AKT/GSK3β/β-catenin signaling pathway in triple negative breast cancer. Mol. Biol. Rep. 2021, 48, 1335–1343. [Google Scholar] [CrossRef]
- Wei, L.; Dong, Y.; Sun, Y.; Mei, X.; Ma, X.; Shi, J.; Yang, Q.; Ji, Y.; Zhang, Z.; Sun, H.; et al. Anticancer property of Hemp Bioactive Peptides in Hep3B liver cancer cells through Akt/GSK3β/β-catenin signaling pathway. Food Sci. Nutr. 2021, 9, 1833–1841. [Google Scholar] [CrossRef]
- Liu, Y.; Duysen, E.; Yaktine, A.L.; Au, A.; Wang, W.; Birt, D.F. Dietary energy restriction inhibits ERK but not JNK or p38 activity in the epidermis of SENCAR mice. Carcinogenesis 2001, 22, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.-H.; Ten, Z.; Wang, X.-S.; Yang, X.-Q. Physicochemical and Functional Properties of Hemp (Cannabis sativa L.) Protein Isolate. J. Agric. Food Chem. 2006, 54, 8945–8950. [Google Scholar] [CrossRef] [PubMed]
- House, J.D.; Neufeld, J.; Leson, G. Evaluating the Quality of Protein from Hemp Seed (Cannabis sativa L.) Products Through the use of the Protein Digestibility-Corrected Amino Acid Score Method. J. Agric. Food Chem. 2010, 58, 11801–11807. [Google Scholar] [CrossRef] [PubMed]
- Callaway, J.C. Hempseed as a nutritional resource: An overview. Euphytica 2004, 140, 65–72. [Google Scholar] [CrossRef]
- Norajit, K.; Gu, B.-J.; Ryu, G.-H. Effects of the addition of hemp powder on the physicochemical properties and energy bar qualities of extruded rice. Food Chem. 2011, 129, 1919–1925. [Google Scholar] [CrossRef]
- Grasso, N.; Alonso-Miravalles, L.; O’Mahony, J.A. Composition, Physicochemical and Sensorial Properties of Commercial Plant-Based Yogurts. Foods 2020, 9, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pojić, M.; Hadnađev, T.D.; Hadnađev, M.; Rakita, S.; Brlek, T. Bread Supplementation with Hemp Seed Cake: A By-Product of Hemp Oil Processing. J. Food Qual. 2015, 38, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Zając, M.; Guzik, P.; Kulawik, P.; Tkaczewska, J.; Florkiewicz, A.; Migdał, W. The quality of pork loaves with the addition of hemp seeds, de-hulled hemp seeds, hemp protein and hemp flour. LWT 2019, 105, 190–199. [Google Scholar] [CrossRef]
- Korus, J.; Witczak, M.; Ziobro, R.; Juszczak, L. Hemp (Cannabis sativa subsp. sativa) flour and protein preparation as natural nutrients and structure forming agents in starch based gluten-free bread. Food Sci. Technol. 2017, 84, 143–150. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Baron, C.P.; Nielsen, N.S.; Otte, J.; Jacobsen, C. Antioxidant activity of yoghurt peptides, Part 2-Characterisation of peptide fractions. Food Chem 2010, 123, 1090–1097. [Google Scholar] [CrossRef]
- Dabija, A.; Codina, G.G.; Gâtlan A-MSănduleac, E.T.; Rusu, L. Effects of some vegetables proteins addition on yogurt quality. Food Ind. 2018, 13, 181–192. Available online: https://doaj.org/article/81642b004c9642a49dfea3df5e7a5cc4 (accessed on 13 November 2022).
- Douglas, K. Amino Acid Composition of an Organic Brown Rice Protein Concentrate and Isolate Compared to Soy and Whey Concentrates and Isolates. Foods 2014, 3, 394–402. [Google Scholar] [CrossRef]
- Gorissen, S.H.; Crombag, J.J.; Senden, J.M.; Waterval, W.A.; Bierau, J.; Verdijk, L.B.; van Loon, L.J. Protein content and amino acid composition of commercially available plant-based protein isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Aal ES, M.; Hucl, P. Amino acid composition and in vitro protein digestibility of selected ancient wheats and their end products. J. Food Compos. Anal. 2002, 15, 737–747. [Google Scholar] [CrossRef]
- Sampelayo, M.S.; Pérez, M.L.; Extremera, F.G.; Boza, J.J.; Boza, J. Use of different dietary protein sources for lactating goats: Milk production and composition as functions of protein degradability and amino acid composition. J. Dairy Sci. 1999, 82, 555–565. [Google Scholar] [CrossRef]
- Wang, Q.L.; Xiong, Y.L. Processing; Nutrition; and Functionality of Hempseed Protein: A Review. Compr. Rev. Food Sci. Food Safety 2019, 18, 936–952. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Jin, Y.; Xiong, Y.L. Heating-Aided pH Shifting Modifies Hemp Seed Protein Structure, Cross-Linking, and Emulsifying Properties. J. Agric. Food Chem. 2018, 66, 10827–10834. [Google Scholar] [CrossRef]
- Pérez-Ezquerra, P.R.; Sánchez-Morillas, L.; Davila-Ferandez, G.; Ruiz-Hornillos, F.; García, I.C.; Mañas, M.H.; Martínez, J.L.; Bartolomé, B. Contact urticaria to Cannabis sativa due to a lipid transfer protein (LTP). Allergol. mmunopathol. 2015, 43, 231–233. [Google Scholar] [CrossRef]
- Decuyper, I.; Ryckebosch, H.; Van Gasse, A.L.; Sabato, V.; Faber, M.; Bridts, C.H.; Ebo, D.G. Cannabis Allergy: What do We Know Anno 2015. Arch. Immunol. Ther. Exp. 2015, 63, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Decuyper, I.I.; Green, B.J.; Sussman, G.L.; Ebo, D.G.; Silvers, W.S.; Pacheco, K.; King, B.S.; Cohn, J.R.; Zeiger, R.S.; Zeiger, J.S.; et al. Occupational Allergies to Cannabis. J. Allergy Clin. Immunol. Pract. 2020, 8, 3331–3338. [Google Scholar] [CrossRef]
- Decuyper, I.I.; Van Gasse, A.L.; Cop, N.; Sabato, V.; Faber, M.A.; Mertens, C.; Bridts, C.H.; Hagendorens, M.M.; de Clerck, L.; Rihs, H.P.; et al. Cannabis sativa allergy, looking through the fog. Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-K.; Seo, J.-B.; Lee, M.-Y. Proteomic profiling of hempseed proteins from Cheungsam. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2012, 1824, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Fasoli, E.; Boschin, G.; Lammi, C.; Zanoni, C.; Citterio, A.; Arnoldi, A. Proteomic characterization of hempseed (Cannabis sativa L.). J. Proteom. 2016, 147, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Mamone, G.; Picariello, G.; Ramondo, A.; Nicolai, M.A.; Ferranti, P. Production, digestibility and allergenicity of hemp (Cannabis sativa L.) protein isolates. Food Res. Int. 2019, 115, 562–571. [Google Scholar] [CrossRef] [PubMed]

 represents promote or inactive;
represents promote or inactive;  represents inhibit or inactive.
represents promote or inactive; represents inhibit or inactive.
represents inhibit or inactive.
represents promote or inactive; represents inhibit or inactive. represents promote or active; represents inhibit or inactive.
represents promote or active; represents inhibit or inactive.
represents promote or active; represents inhibit or inactive.
represents promote or active; represents inhibit or inactive. represents promote or active; represents inhibit or inactive.
represents promote or active; represents inhibit or inactive.
represents promote or active; represents inhibit or inactive.
represents promote or active; represents inhibit or inactive.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soybean Hydrolysate | Hemp Hydrolysate | p Value | |
|---|---|---|---|
| ORAC (μM trolox/g sample) | 38.78 ± 0.362 | 0.539 ± 0.124 | <0.001 |
| Metal iron chelating activity (%) | 42.71 ± 0.247 | 0.591 ± 0.152 | <0.05 |
| Amino | Defatted | Hemp Protein | Hemp Protein Isolate | Hemp Protein Fraction | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| His | 3.07 ± 0.01 | 2.78 ± 0.03 | 1.09 ± 0.33 | 2.23 ± 0.20 | 2.85 ± 0.05 | 2.81 ± 0.47 | 2.61 ± 0.06 | 2.47 ± 0.01 | 2.47 ± 0.03 | 2.49 ± 0.08 |
| Arg | 12.68 ± 0.21 | 14.07 ± 0.40 | 6.05 ± 0.56 | 10.64 ± 0.92 | 13.44 ± 0.03 | 9.91 ± 0.91 | 13.87 ± 0.11 | 12.96 ± 0.13 | 13.6 ± 0.25 | 13.31 ± 0.24 |
| Thr | 3.79 ± 0.08 | 3.68 ± 0.29 | 2.06 ± 0.18 | 3.36 ± 0.37 | 3.79 ± 0.10 | 4.57 ± 0.35 | 3.60 ± 0.04 | 3.77 ± 0.35 | 4.01 ± 0.19 | 4.00 ± 0.19 |
| Val | 5.27 ± 0.11 | 4.66 ± 0.02 | 2.51 ± 0.09 | 3.90 ± 0.12 | 3.77 ± 0.40 | 4.98 ± 0.13 | 5.67 ± 0.14 | 5.26 ± 0.11 | 4.45 ± 0.19 | 4.24 ± 0.14 |
| Met | 2.14 ± 0.11 | 1.81 ± 0.39 | 1.83 ± 0.15 | 1.63 ± 0.05 | 3.79 ± 0.10 | 1.39 ± 0.06 | 1.94 ± 0.04 | 2.03 ± 0.06 | 1.85 ± 0.14 | 1.80 ± 0.15 |
| Lys | 3.78 ± 0.09 | 2.97 ± 0.06 | 1.20 ± 0.23 | 1.97 ± 0.18 | 0.89 ± 0.34 | 4.16 ± 0.87 | 3.19 ± 0.11 | 2.94 ± 0.14 | 2.56 ± 0.03 | 2.51 ± 0.07 |
| Ile | 4.23 ± 0.08 | 3.84 ± 0.07 | 1.93 ± 0.02 | 3.12 ± 0.10 | 3.02 ± 0.40 | 3.99 ± 0.08 | 4.15 ± 0.13 | 4.16 ± 0.11 | 3.98 ± 0.05 | 3.90 ± 0.04 |
| Leu | 7.12 ± 0.05 | 6.75 ± 0.04 | 3.13 ± 0.08 | 4.90 ± 0.19 | 7.13 ± 0.11 | 6.63 ± 0.23 | 9.91 ± 0.06 | 7.26 ± 0.10 | 5.15 ± 0.04 | 4.82 ± 0.43 |
| Phe | 4.76 ± 0.09 | 4.60 ± 0.04 | 2.34 ± 0.19 | 3.86 ± 0.42 | 5.12 ± 0.02 | 4.57 ± 0.11 | 7.68 ± 0.05 | 5.01 ± 0.14 | 3.21 ± 0.02 | 2.85 ± 0.40 |
| Non-essential | ||||||||||
| Trp | 0.76 ± 0.05 | 1.23 ± 0.16 | 0.04 ± 0.00 | 0.02 ± 0.00 | 0.55 ± 0.02 | NA | 1.58 ± 0.01 | 1.44 ± 0.15 | 1.16 ± 0.12 | 1.11 ± 0.02 |
| Asp | 11.34 ± 0.37 | 11.39 ± 0.03 | 4.06 ± 0.40 | 6.55 ± 0.48 | 12.35 ± 0.34 | 9.41 ± 0.39 | 9.49 ± 0.06 | 11.70 ± 0.29 | 12.79 ± 0.47 | 12.70 ± 0.11 |
| Ser | 5.57 ± 0.11 | 4.63 ± 0.09 | 2.37 ± 0.23 | 3.90 ± 0.31 | 6.31 ± 0.21 | 5.18 ± 0.02 | 4.73 ± 0.21 | 4.79 ± 0.19 | 4.69 ± 0.07 | 4.47 ± 0.38 |
| Glu | 18.41 ± 0.13 | 20.06 ± 1.34 | 6.92 ± 0.45 | 11.46 ± 0.82 | 18.86 ± 0.35 | 16.14 ± 0.26 | 15.18 ± 0.96 | 19.31 ± 1.04 | 22.71 ± 1.58 | 22.87 ± 1.02 |
| Gly | 4.74 ± 0.07 | 4.29 ± 0.23 | 2.01 ± 0.24 | 3.19 ± 0.29 | 4.79 ± 0.23 | 3.99 ± 0.06 | 3.23 ± 0.06 | 3.93 ± 0.07 | 4.54 ± 0.20 | 4.71 ± 0.44 |
| Ala | 4.89 ± 0.10 | 4.47 ± 0.16 | 1.79 ± 0.04 | 2.73 ± 0.09 | 5.01 ± 0.14 | 4.50 ± 0.36 | 4.91 ± 0.06 | 4.77 ± 0.12 | 4.30 ± 0.19 | 4.12 ± 0.09 |
| Pro | 2.66 ± 0.69 | 4.00 ± 0.07 | 1.73 ± 0.07 | 2.66 ± 0.10 | 3.02 ± 0.00 | 4.53 ± 0.39 | 3.19 ± 0.33 | 4.04 ± 0.56 | 4.23 ± 0.11 | 4.89 ± 0.73 |
| Cys | 1.22 ± 0.02 | 1.32 ± 0.23 | 0.71 ± 0.02 | 0.99 ± 0.06 | 0.89 ± 0.34 | 0.17 ± 0.01 | 0.29 ± 0.13 | 0.66 ± 0.01 | 1.26 ± 0.07 | 1.58 ± 0.28 |
| Tyr | 3.79 ± 0.08 | 3.45 ± 0.00 | 1.51 ± 0.13 | 2.71 ± 0.26 | 3.79 ± 0.10 | 3.67 ± 0.23 | 4.78 ± 0.02 | 3.50 ± 0.02 | 3.06 ± 0.06 | 3.62 ± 0.96 |
| Reference | [76] | [59] | [77] | [77] | [76] | [7] | [59] | [59] | [59] | [59] |
| Protein Resource | Hydrolytic Enzyme | In Vitro Antioxidant Assays | Amino Acid Sequence | Reference |
|---|---|---|---|---|
| Sardine Muscle | Pepsin | Superoxide radical scavenging activity, hydroxyl radical scavenging activity. | Leu-Gln-Pro-Gly-Gln-Gly-Gln-Gln | [86] |
| Casein | Pepsin | Superoxide radical scavenging activity, hydroxyl radical scavenging activity, DPPH radical scavenging activity. | Tyr-Phe-Tyr-Pro-Glu-Leu | [87] |
| Royal Jelly | Protease N | Superoxide radical scavenging activity, hydroxyl radical scavenging activity, hydrogen peroxide scavenging activity, metal chelating activity. | Ala-Leu, PheLys, Phe-Arg, lle-Arg, Lys-Phe, Lys-Leu, Lys-Tyr, Arg-Tyr, Tyr-Asp, Tyr-Tyr, Leu-Asp-Arg, Lys-Asn-Tyr-Pro | [88] |
| Egg White | Pepsin | ORAC, low-density lipoprotein lipid oxidation induced by Cu2+. | Tyr-Ala-Glu-Glu-Arg-Tyr-Pro-Ile-Leu | [89] |
| Porcine Myofibrillar | Actinase E, Papain | DPPH radical scavenging activity, metal ion chelating activity, hydroperoxides in a peroxidation system. | Asp-Ser-Gly-Val-Thr, Ile-Glu-Ala-Glu-Gly-Glu, Asp-Ala-Gln-Glu-Lys-Leu-Glu, Glu-Glu-Leu-Asp-Asn-Ala-Leu-Asn, Val-Pro-Ser-Ile-Asp-Asp-Gln-Glu-Glu-Leu-Met | [90] |
| Sardinelle | Alcalase | DPPH radical-scavenging assay, the lipid peroxidation inhibition activity, reducing power assay. | Leu-Ala-Arg-Leu, Gly-Gly-Glu, Leu-His-Tyr and Gly-Ala-Leu-Ala-Ala-His | [78] |
| Barley | Alcalase, Flvourzyme and Pepsin | DPPH radical scavenging assay, superoxide radical scavenging assay, hydroxyl radical scavenging assay, metal chelating activity, ORAC. | Pro-Tyr-Pro, Gln-Pro-Tyr-Pro-Gln, Gln-Gln-Pro-Tyr-Pro-Gln, Gln-Pro-Gln-Pro-Tyr-Pro-Gln, Thr-Gln-Gln-Pro-Tyr-Pro-Gln, Glu-Pro-Tyr-Pro-Glu | [80] |
| Corn | Alcalase | Superoxide dismutase activity. | Leu-Asp-Tyr-Glu | [79] |
| Amino Acids | Hemp Seed | Brown Rice | Soybean | Brown Rice | Oat | Pea | Potato | Wheat | Fababean | Corn | Infant (FAO/WHO/UNU) | Child 2–5 Years Old (FAO/WHO/UNU) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Essential | ||||||||||||
| His | 3.07 | 1.8 | 2.3 | 1.8 | 0.9 | 1.6 | 1.4 | 1.7 | 2.7 | 2.2 | 2.6 | 1.90 |
| Arg | 12.68 | 6.3 | 1.8 | 6.3 | 3.1 | 5.9 | 3.3 | 4.1 | 11.2 | 6.3 | NA | NA |
| Thr | 3.79 | 2.9 | 7.9 | 2.9 | 1.5 | 2.5 | 4.1 | 2.3 | 3.1 | 3.4 | 4.3 | 3.40 |
| Val | 5.27 | 4.6 | 5.9 | 4.6 | 2.0 | 2.7 | 3.7 | 3.6 | 6.8 | 7.3 | 5.5 | 3.50 |
| Met | 2.14 | 2.3 | 1.7 | 2.3 | 0.1 | 0.3 | 1.3 | 1.1 | 0.7 | 1.6 | NA | NA |
| Lys | 3.78 | 2.4 | 9.7 | 2.4 | 1.3 | 4.7 | 4.8 | 2.0 | 7.5 | 4.3 | 6.6 | 5.80 |
| Ile | 4.23 | 3.5 | 4.3 | 3.5 | 1.3 | 2.3 | 3.1 | 3.2 | 4.3 | 2.6 | 4.6 | 2.80 |
| Leu | 7.12 | 6.4 | 10.2 | 6.4 | 3.8 | 5.7 | 6.7 | 6.7 | 9.6 | 10.4 | 9.3 | 6.60 |
| Phe | 4.76 | 4.4 | 2.6 | 4.4 | 2.7 | 3.7 | 4.2 | 2.3 | 5.1 | 4.1 | NA | NA |
| Non-essential | ||||||||||||
| Trp | 0.76 | 1.2 | 1.1 | 1.2 | NA | NA | NA | 1.2 | NA | NA | NA | NA |
| Asp | 11.34 | 6.9 | 10.2 | 6.9 | NA | NA | NA | 4.5 | 7.6 | 7.1 | NA | N |
| Ser | 5.57 | 3.9 | 4.6 | 3.9 | 2.2 | 3.6 | 3.4 | 4.2 | 5.6 | 5.5 | NA | NA |
| Glu | 18.41 | 13.9 | 17.5 | 13.9 | 11.0 | 12.9 | 7.1 | 36.1 | 18.1 | 18.7 | NA | NA |
| Gly | 4.74 | 3.5 | 3.6 | 3.5 | 1.7 | 2.8 | 3.2 | 3.3 | 5.3 | 6.6 | NA | NA |
| Ala | 4.89 | 4.5 | 4.8 | 4.5 | 2.2 | 3.2 | 3.3 | 2.8 | 4.7 | 7.6 | NA | NA |
| Pro | 2.66 | 2.9 | 5.7 | 2.9 | 2.5 | 3.1 | 3.3 | 8.4 | 5.4 | 9.0 | NA | NA |
| Cys | 1.22 | 1.7 | 2.1 | 1.7 | 0.4 | 0.2 | 0.3 | 2.0 | 1.0 | 1.9 | NA | NA |
| Tyr | 3.79 | 4.3 | 1.1 | 4.3 | 1.5 | 2.6 | 3.8 | 4.0 | 1.3 | 1.4 | 1.7 | 1.10 |
| Reference | [76] | [135] | [135] | [135] | [136] | [136] | [136] | [137] | [138] | [138] | [139] | [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Griffin, J.; Li, Y.; Wang, D.; Wang, W. Antioxidant Properties of Hemp Proteins: From Functional Food to Phytotherapy and Beyond. Molecules 2022, 27, 7924. https://doi.org/10.3390/molecules27227924
Zhang J, Griffin J, Li Y, Wang D, Wang W. Antioxidant Properties of Hemp Proteins: From Functional Food to Phytotherapy and Beyond. Molecules. 2022; 27(22):7924. https://doi.org/10.3390/molecules27227924
Chicago/Turabian StyleZhang, Jiejia, Jason Griffin, Yonghui Li, Donghai Wang, and Weiqun Wang. 2022. "Antioxidant Properties of Hemp Proteins: From Functional Food to Phytotherapy and Beyond" Molecules 27, no. 22: 7924. https://doi.org/10.3390/molecules27227924
APA StyleZhang, J., Griffin, J., Li, Y., Wang, D., & Wang, W. (2022). Antioxidant Properties of Hemp Proteins: From Functional Food to Phytotherapy and Beyond. Molecules, 27(22), 7924. https://doi.org/10.3390/molecules27227924