
Control Efficacy of Salicylic Acid Microcapsules against Postharvest Blue Mold in Apple Fruit
 and
and
Abstract
:1. Introduction
2. Results and Discussion
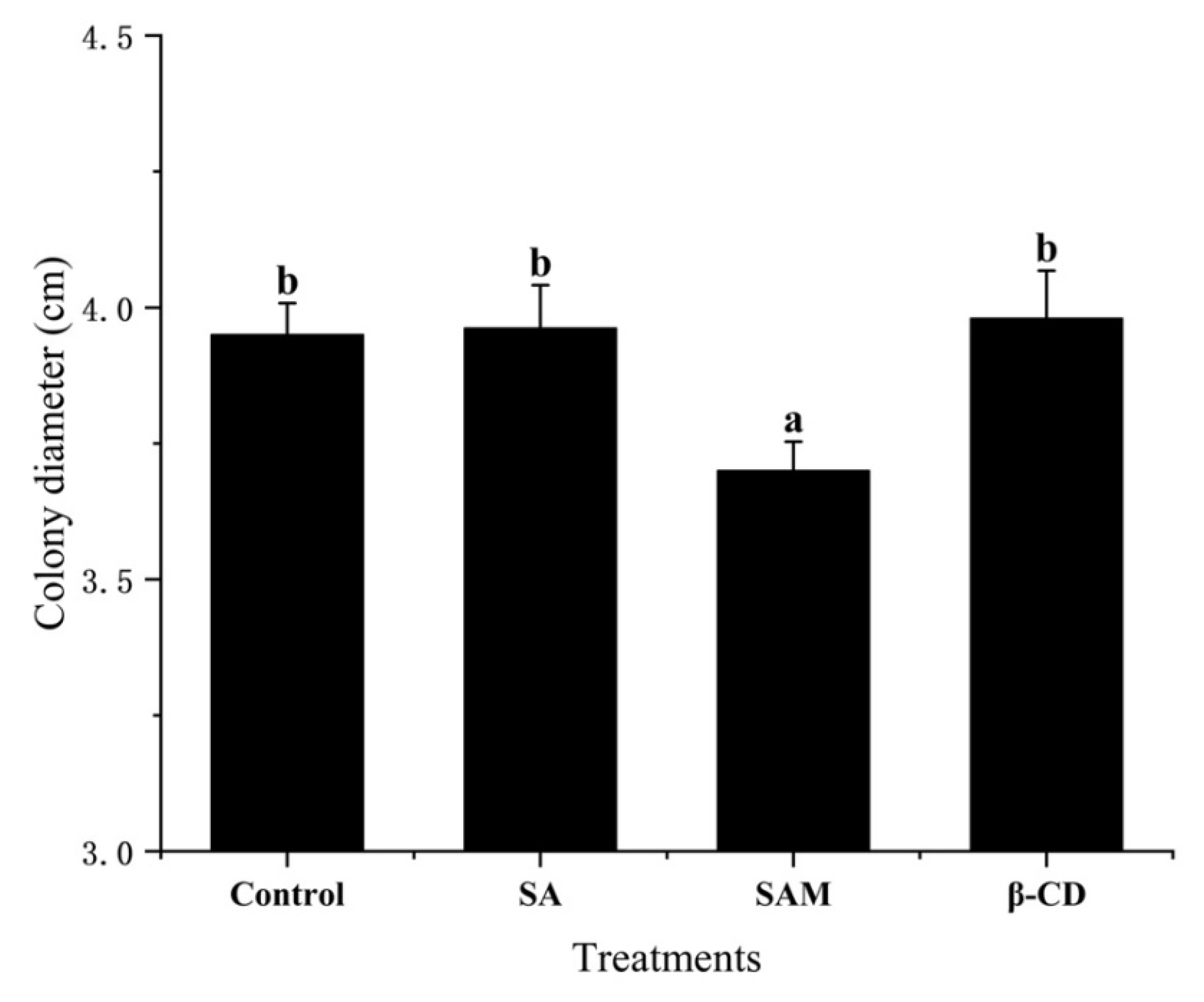
2.1. Effect of SAM on Control of P. expansum In Vitro
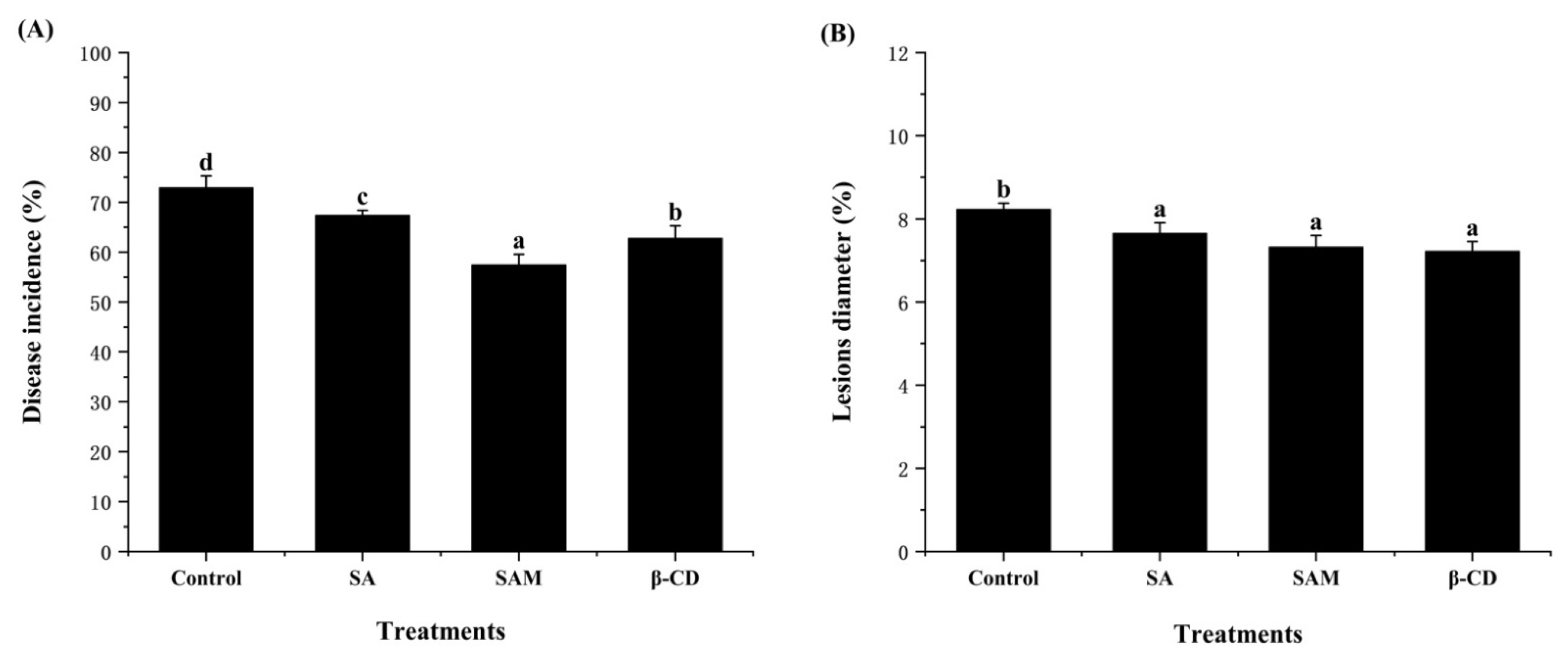
2.2. Effect of SAM on Control of Blue Mold in Apple Fruit
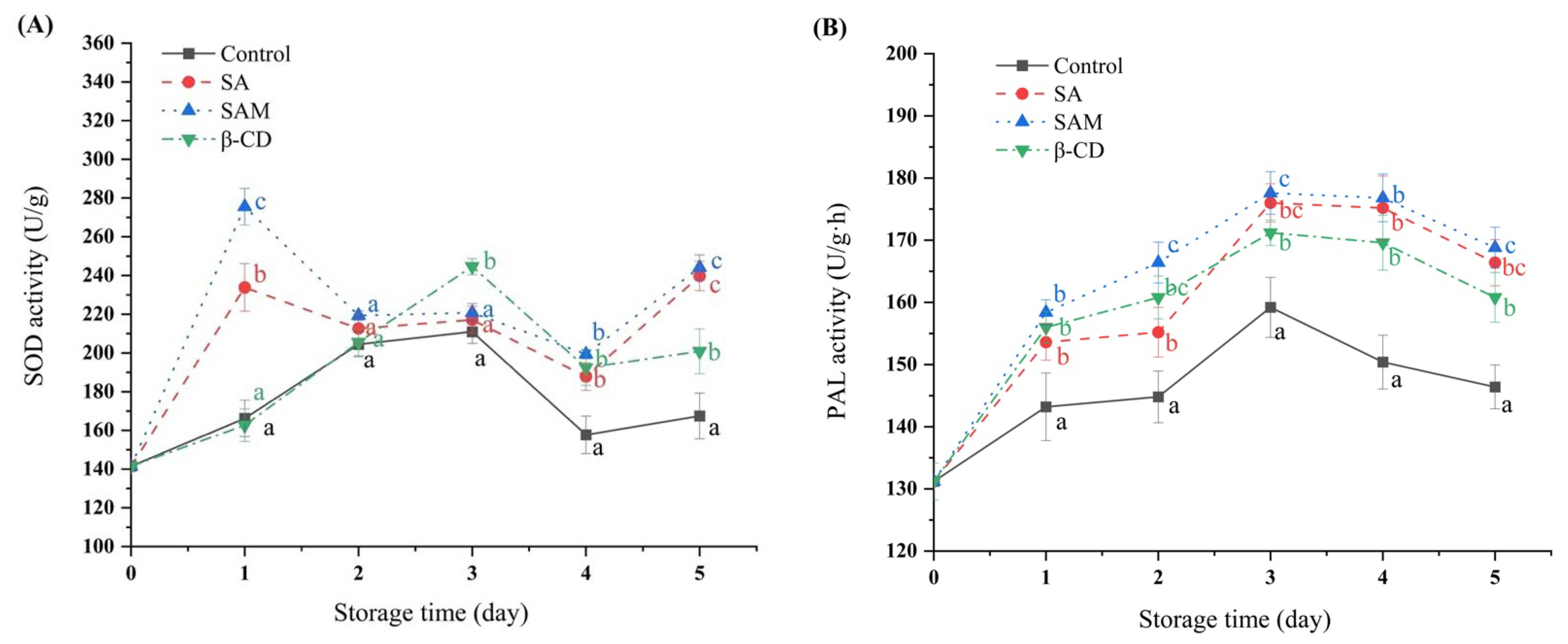
2.3. Effect of SAM on Antioxidant Enzyme Activities in Apple Fruit
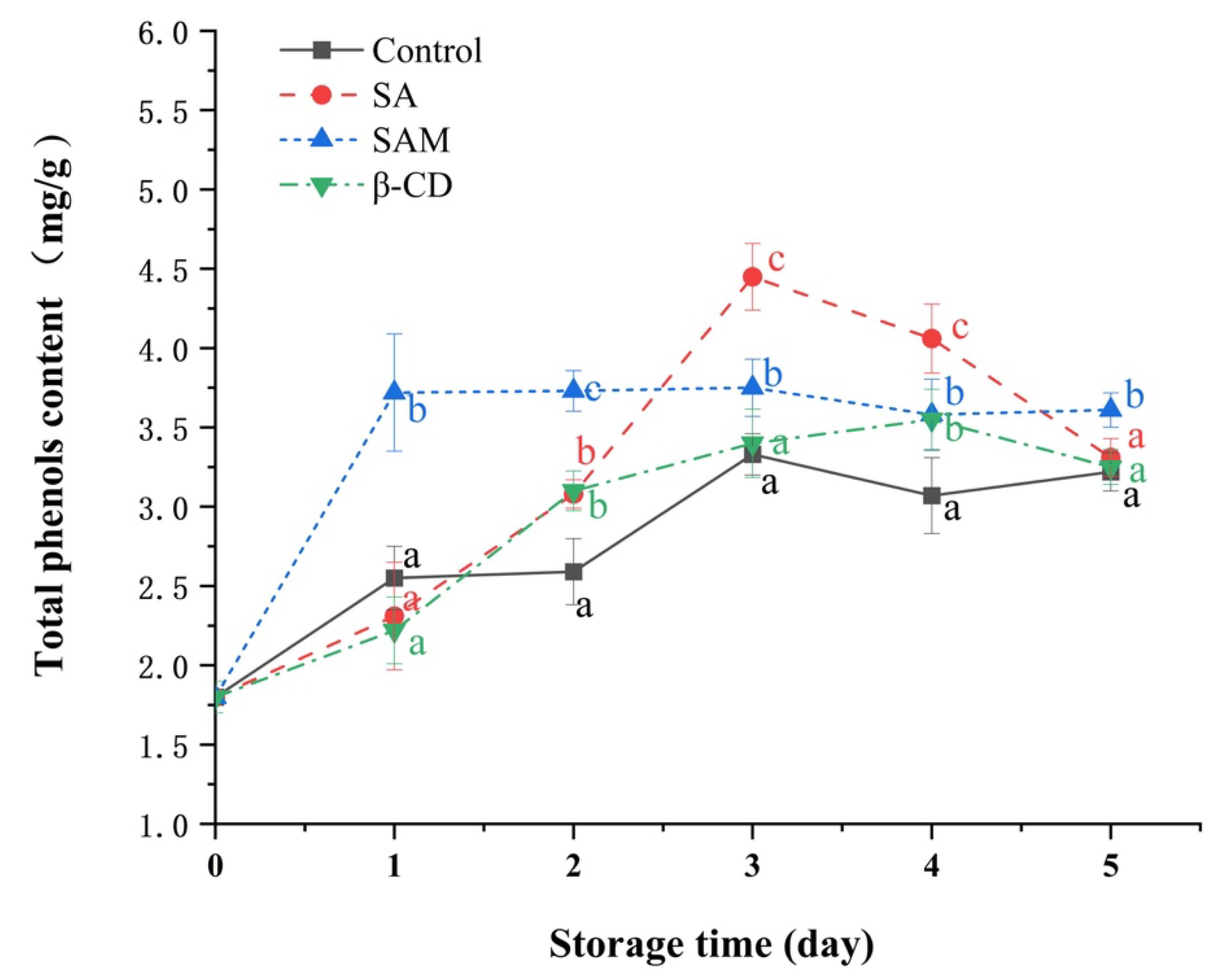
2.4. Effect of SAM on Total Phenolic Contents in Apple Fruit
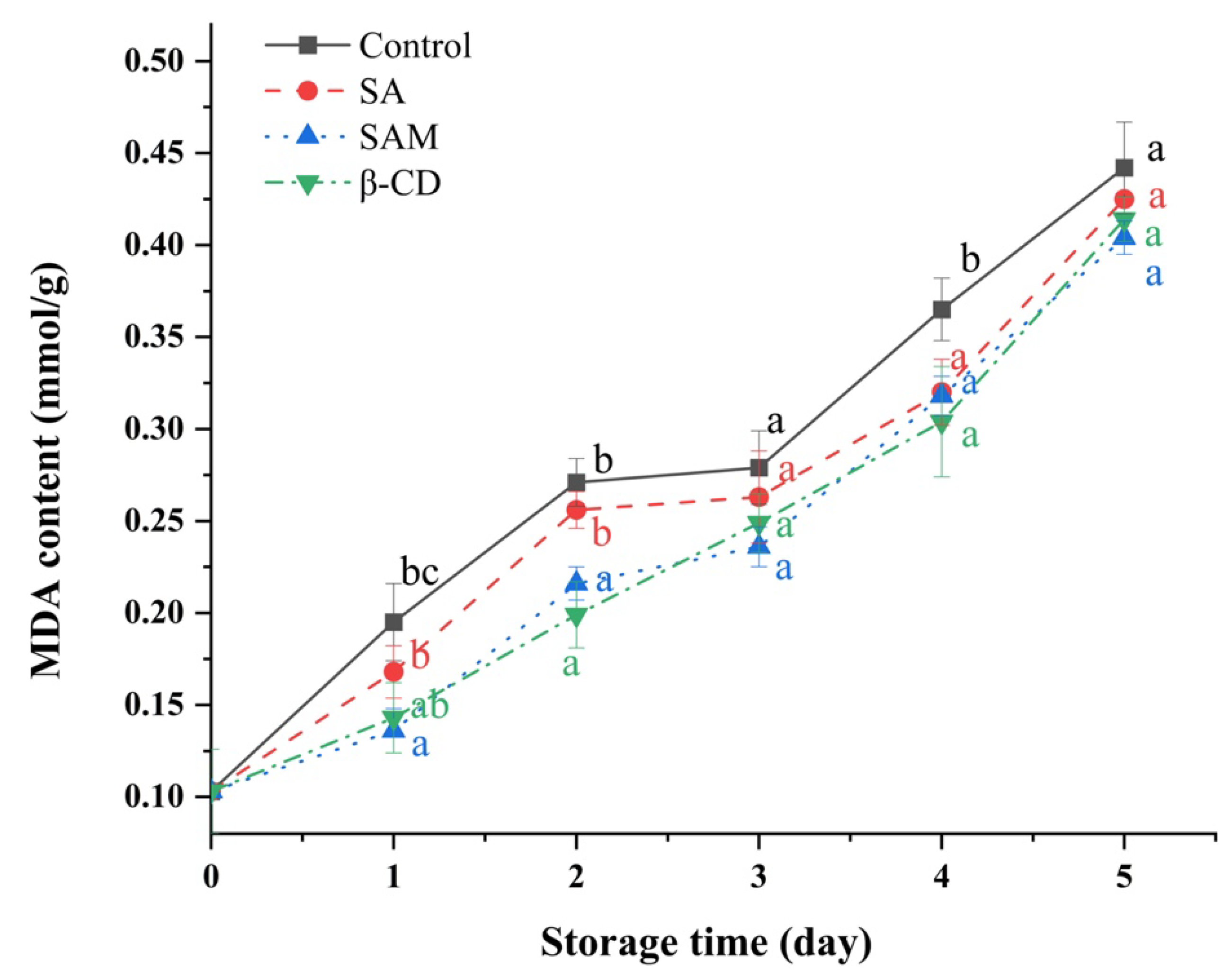
2.5. Influence of SAM on Malondialdehyde Contents in Apple Fruit
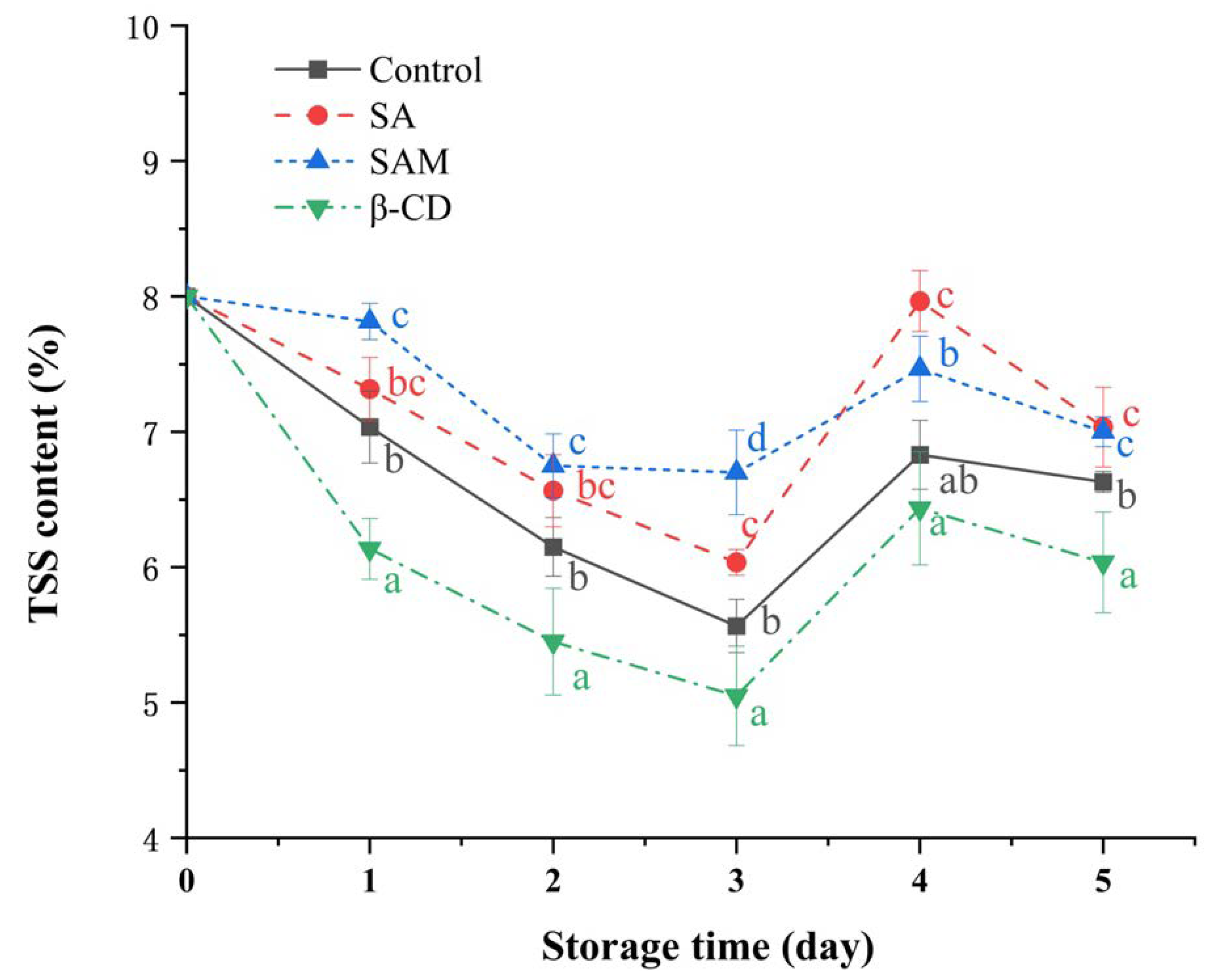
2.6. Impact of SAM on Postharvest Quality of Apple Fruit
3. Materials and Methods
3.1. Fruit Materials
3.2. Fungal Pathogens
3.3. Preparation of SAM
3.4. Effect of SAM on Control of P. expansum In Vitro
3.5. Effect of SAM on Control of P. expansum In Vivo
3.6. Effect of SAM on Enzyme Activities and Total Phenolic Contents in Apple Fruit
3.7. Influence of SAM on MDA Contents in Apple Fruit
3.8. Impact of SAM on Quality Parameters of Apple Fruit
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Abdel-Sattar, M.; Kotb, H.R.M. Nutritional status and productivity of Anna apple trees in the year following autumn irrigation determent. Agric. Water Manag. 2021, 252, 106882. [Google Scholar] [CrossRef]
- He, J.H.; Ma, L.J.; Wang, D.J.; Zhang, M.Y.; Zhou, H.L. Ferulic acid treatment reinforces the resistance of postharvest apple fruit during gray mold infection. J. Plant Pathol. 2019, 101, 503–511. [Google Scholar] [CrossRef]
- Wang, K.L.; Ngea, G.L.N.; Godana, E.A.; Shi, Y.; Lanhuang, B.; Zhang, X.Y.; Zhao, L.N.; Yang, Q.Y.; Wang, S.Y.; Zhang, H.Y. Recent advances in Penicillium expansum infection mechanisms and current methods in controlling P. expansum in postharvest apples. Crit. Rev. Food Sci. Nutr. 2021. [Google Scholar] [CrossRef] [PubMed]
- Li, J.K.; Lei, H.H.; Song, H.M.; Lai, T.F.; Xu, X.B.; Shi, X.Q. 1-methylcyclopropene (1-MCP) suppressed postharvest blue mold of apple fruit by inhibiting the growth of Penicillium expansum. Postharvest Biol. Technol. 2017, 125, 59–64. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Wang, D.J.; Gao, X.X.; Yue, Z.Y.; Zhou, H.L. Exogenous caffeic acid and epicatechin enhance resistance against Botrytis cinerea through activation of the phenylpropanoid pathway in apples. Sci. Hortic. 2020, 268, 109348. [Google Scholar] [CrossRef]
- Ahima, J.; Zhang, X.Y.; Yang, Q.Y.; Zhao, L.N.; Tibiru, A.M.; Zhang, H.Y. Biocontrol activity of Rhodotorula mucilaginosa combined with salicylic acid against Penicillium digitatum infection in oranges. Biol. Control 2019, 135, 23–32. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Song, C.C.; Qi, S.N.; Lin, Q.; Duan, Y.Q. Jasmonic acid and salicylic acid induce the accumulation of sucrose and increase resistance to chilling injury in peach fruit. J. Sci. Food Agric. 2021, 101, 4250–4255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shi, X.P.; Li, B.H.; Zhang, Q.M.; Liang, W.X.; Wang, C.X. Salicylic acid confers enhanced resistance to Glomerella leaf spot in apple. Plant Physiol. Biochem. 2016, 106, 64–72. [Google Scholar] [CrossRef]
- Jiao, W.X.; Li, X.X.; Wang, X.M.; Cao, J.K.; Jiang, W.B. Chlorogenic acid induces resistance against Penicillium expansum in peach fruit by activating the salicylic acid signaling pathway. Food Chem. 2018, 260, 274–282. [Google Scholar] [CrossRef]
- Haider, S.A.; Ahmad, S.; Khan, A.S.; Anjum, M.A.; Nasir, M.; Naz, S. Effects of salicylic acid on postharvest fruit quality of “Kinnow” mandarin under cold storage. Sci. Hortic. 2020, 259, 108843. [Google Scholar] [CrossRef]
- Mahattanadul, N.; Sunintaboon, P.; Sirithip, P.; Tuchinda, P. Chitosan-functionalised poly(2-hydroxyethyl methacrylate) core-shell microgels as drug delivery carriers: Salicylic acid loading and release. J. Microencapsul. 2016, 33, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.P.; Li, Y.C.; Long, H.T.; Liu, Z.T.; Huang, Y.; Zhang, M.; Wang, T.L.; Liu, Y.X.; Bi, Y.; Prusky, D.B. Preparation and use of trans-2-hexenal microcapsules to preserve ‘Zaosu’ pears. Sci. Hortic. 2021, 283, 110091. [Google Scholar] [CrossRef]
- Yan, L.; Zeng, F.Y.; Chen, Z.J.; Chen, S.; Lei, Y.F. Improvement of wood decay resistance by salicylic acid/silica microcapsule: Effects on the salicylic leaching, microscopic structure and decay resistance. Int. Biodeter. Biodegr. 2021, 156, 105134. [Google Scholar] [CrossRef]
- Yang, Z.; Fang, Y.; Ji, H. Controlled release and enhanced antibacterial activity of salicylic acid by hydrogen bonding with chitosan. Chin. J. Chem. Eng. 2016, 24, 421–426. [Google Scholar] [CrossRef]
- Yang, W.H.; Wang, L.; Ban, Z.J.; Yan, J.W.; Lu, H.Y.; Zhang, X.C.; Wu, Q.; Aghdam, M.S.; Luo, Z.S.; Li, L. Efficient microencapsulation of Syringa essential oil; the valuable potential on quality maintenance and storage behavior of peach. Food Hydrocolloid. 2019, 95, 177–185. [Google Scholar] [CrossRef]
- Dou, S.W.; Ouyang, Q.L.; You, K.Y.; Qian, J.J.; Tao, N.G. An inclusion complex of thymol into β-cyclodextrin and its antifungal activity against Geotrichum citri-aurantii. Postharvest Biol. Technol. 2018, 138, 31–36. [Google Scholar] [CrossRef]
- Mandal, S.; Mallick, N.; Mitra, A. Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. Plant Physiol. Biochem. 2009, 47, 642–649. [Google Scholar] [CrossRef]
- Li, D.H.; Wu, H.J.; Huang, W.; Guo, L.; Dou, H.T. Microcapsule of sweet orange essential oil encapsulated in beta-cyclodextrin improves the release behaviors in vitro and in vivo. Eur. J. Lipid Sci. Technol. 2018, 120, 1700521. [Google Scholar] [CrossRef]
- Lin, Y.; Huang, R.; Sun, X.; Yu, X.; Xiao, Y.; Wang, L.; Hu, W.; Zhong, T. The p-Anisaldehyde/β-cyclodextrin inclusion complexes as fumigation agent for control of postharvest decay and quality of strawberry. Food Control 2021, 130, 108346. [Google Scholar] [CrossRef]
- Gong, L.; Li, T.; Chen, F.; Duan, X.; Yuan, Y.; Zhang, D.; Jiang, Y. An inclusion complex of eugenol into β-cyclodextrin: Preparation, and physicochemical and antifungal characterization. Food Chem. 2016, 196, 324–330. [Google Scholar] [CrossRef]
- Wang, Y.; Bian, W.; Ren, X.; Song, X.; He, S. Microencapsulation of clove essential oil improves its antifungal activity against Penicillium digitatum in vitro and green mould on Navel oranges. J. Hortic. Sci. Biotechnol. 2018, 93, 159–166. [Google Scholar] [CrossRef]
- da Rocha Neto, A.C.; Luiz, C.; Maraschin, M.; Di Piero, R.M. Efficacy of salicylic acid to reduce Penicillium expansum inoculum and preserve apple fruits. Int. J. Food Microbiol. 2016, 221, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Li, S.E.; Xu, Y.H.; Bi, Y.; Zhang, B.; Shen, S.L.; Jiang, T.J.; Zheng, X.L. Melatonin treatment inhibits gray mold and induces disease resistance in cherry tomato fruit during postharvest. Postharvest Biol. Technol. 2019, 157, 110962. [Google Scholar] [CrossRef]
- Tang, T.T.; Xie, X.F.; Ren, X.; Wang, W.J.; Tang, X.M.; Zhang, J.; Wang, Z.D. A difference of enzymatic browning unrelated to PPO from physiology, targeted metabolomics and gene expression analysis in Fuji apples. Postharvest Biol. Technol. 2020, 170, 111323. [Google Scholar] [CrossRef]
- Patel, M.K.; Maurer, D.; Feygenberg, O.; Ovadia, A.; Elad, Y.; Oren-Shamir, M.; Alkan, N. Phenylalanine: A promising inducer of fruit resistance to postharvest pathogens. Foods 2020, 9, 646. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Wu, F.; Gu, N.; Yan, X.L.; Wang, K.L.; Dhanasekaran, S.; Gu, X.Y.; Zhao, L.N.; Zhang, H.Y. Postharvest biological control of Rhizopus rot and the mechanisms involved in induced disease resistance of peaches by Pichia membranefaciens. Postharvest Biol. Technol. 2020, 163, 111146. [Google Scholar] [CrossRef]
- Ma, L.J.; He, J.H.; Liu, H.; Zhou, H.L. The phenylpropanoid pathway affects apple fruit resistance to Botrytis cinerea. J. Phytopathol. 2018, 166, 206–215. [Google Scholar] [CrossRef]
- Buron-Moles, G.; Torres, R.; Amoako-Andoh, F.; Vinas, I.; Teixido, N.; Usall, J.; Keulemans, W.; Davey, M.W. Analysis of changes in protein abundance after wounding in ‘Golden Delicious’ apples. Postharvest Biol. Technol. 2014, 87, 51–60. [Google Scholar] [CrossRef]
- Moosa, A.; Sahi, S.T.; Khan, S.A.; Malik, A.U. Salicylic acid and jasmonic acid can suppress green and blue moulds of citrus fruit and induce the activity of polyphenol oxidase and peroxidase. Folia Hortic. 2019, 31, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.Q.; Li, Y.; Fan, H.F.; Xia, Q. Preparation and characterization of salicylic acid-loaded microcapsules as delivery systems for cosmetics. Integr. Ferroelectr. 2014, 152, 22–28. [Google Scholar] [CrossRef]
- Wang, D.; Chen, Q.Y.; Chen, W.W.; Guo, Q.G.; Xia, Y.; Wu, D.; Jing, D.L.; Liang, G.L. Melatonin treatment maintains quality and delays lignification in loquat fruit during cold storage. Sci. Hortic. 2021, 284, 110126. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.X.; Li, F.F.; Hong, K.Q.; Yuan, D.B. Effects of procyanidin treatment on the ripening and softening of banana fruit during storage. Sci. Hortic. 2022, 292, 110644. [Google Scholar] [CrossRef]
- Wen, M.; Lin, X.; Yu, Y.S.; Wu, J.J.; Xu, Y.J.; Xiao, G.S. Natamycin treatment reduces the quality changes of postharvest mulberry fruit during storage. J. Food Biochem. 2019, 43, e12934. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Gong, D.; Liang, G.; Han, R.; Xie, J.; Li, W. Enhanced preservation effects of sugar apple fruits by salicylic acid treatment during post-harvest storage. J. Sci. Food Agric. 2008, 88, 2693–2699. [Google Scholar] [CrossRef]
- Oviedo-Mireles, J.C.; Soto-Parra, J.M.; Sanchez, E.; Yanez-Munoz, R.M.; Perez-Leal, R.; Noperi-Mosqueda, L.C. Salicylic acid and nutrient immersion to maintain apple quality and bioactive compounds in postharvest. Not. Bot. Horti Agrobot. 2021, 49, 12409. [Google Scholar] [CrossRef]
- Madanipour, S.; Alimohammadi, M.; Rezaie, S.; Nabizadeh, R.; Khaniki, G.J.; Hadi, M.; Yousefi, M.; Bidgoli, S.M.; Yousefzadeh, S. Influence of postharvest application of chitosan combined with ethanolic extract of liquorice on shelflife of apple fruit. J. Environ. Health Sci. Eng. 2019, 17, 331–336. [Google Scholar] [CrossRef]
- Matabura, V.V. Modelling of firmness variability of Jonagold apple during postharvest storage. J. Food Sci. Technol. 2022, 59, 1487–1498. [Google Scholar] [CrossRef]
- Beghi, R.; Giovanelli, G.; Malegori, C.; Giovenzana, V.; Guidetti, R. Testing of a VIS-NIR system for the monitoring of long-term apple storage. Food Bioprocess Technol. 2014, 7, 2134–2143. [Google Scholar] [CrossRef]
- Lu, H.; Lu, L.; Zeng, L.; Fu, D.; Xiang, H.; Yu, T.; Zheng, X. Effect of chitin on the antagonistic activity of Rhodosporidium paludigenum against Penicillium expansum in apple fruit. Postharvest Biol. Technol. 2014, 92, 9–15. [Google Scholar] [CrossRef]
- Song, X.; Li, R.; Zhang, Q.; He, S.; Wang, Y. Antibacterial effect and possible mechanism of salicylic acid microcapsules against Escherichia coli and Staphylococcus aureus. Int. J. Environ. Res. Public Health 2022, 19, 12761. [Google Scholar] [CrossRef]
- Li, X.; Fu, H. Fungal spore aerosolization at different positions of a growing colony blown by airflow. Aerosol Air Qual. Res. 2020, 20, 2826–2833. [Google Scholar] [CrossRef]
- Mirshekari, A.; Madani, B.; Yahia, E.M.; Golding, J.B.; Vand, S.H. Postharvest melatonin treatment reduces chilling injury in sapota fruit. J. Sci. Food Agric. 2020, 100, 1897–1903. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.P.; Kahramanoglu, I.; Chen, J.Y.; Gan, Z.Y.; Chen, C.Y. Effects of hot air treatments on postharvest storage of Newhall navel orange. Plants 2020, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.L.; Zhao, H.D.; Zhang, J.; Sheng, Z.T.; Cao, J.K.; Jiang, W.B. Different molecular weights chitosan coatings delay the senescence of postharvest nectarine fruit in relation to changes of redox state and respiratory pathway metabolism. Food Chem. 2019, 289, 160–168. [Google Scholar] [CrossRef]
- He, S.; Zhan, Z.; Shi, C.; Wang, S.; Shi, X. Ethanol at subinhibitory concentrations enhances biofilm formation in Salmonella Enteritidis. Foods 2022, 11, 2237. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Day | Firmness (N) | |||
|---|---|---|---|---|
| Control | SA | SAM | β-CD | |
| 0 | 2.82 ± 0.03 a | 2.82 ± 0.03 a | 2.82 ± 0.03 a | 2.82 ± 0.03a |
| 1 | 2.69 ± 0.20 a | 2.78 ± 0.05 a | 2.80 ± 0.18 b | 2.80 ± 0.06 a |
| 2 | 2.53 ± 0.13 a | 2.76 ± 0.09 b | 2.79 ± 0.13 b | 2.62 ± 0.09 a |
| 3 | 2.68 ± 0.13 a | 2.84 ± 0.15 b | 2.72±0.06 ab | 2.72±0.16 ab |
| 4 | 2.58 ± 0.08 a | 2.73 ± 0.15 b | 2.93 ± 0.05 c | 2.71 ± 0.15 b |
| 5 | 2.55 ± 0.11 a | 2.58 ± 0.10 a | 2.77 ± 0.04 c | 2.67 ± 0.10 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Chen, J.; Bian, W.; Yang, X.; Ye, L.; He, S.; Song, X. Control Efficacy of Salicylic Acid Microcapsules against Postharvest Blue Mold in Apple Fruit. Molecules 2022, 27, 8108. https://doi.org/10.3390/molecules27228108
Wang Y, Chen J, Bian W, Yang X, Ye L, He S, Song X. Control Efficacy of Salicylic Acid Microcapsules against Postharvest Blue Mold in Apple Fruit. Molecules. 2022; 27(22):8108. https://doi.org/10.3390/molecules27228108
Chicago/Turabian StyleWang, Yifei, Jiahao Chen, Wenyi Bian, Xiaobo Yang, Lin Ye, Shoukui He, and Xiaoqiu Song. 2022. "Control Efficacy of Salicylic Acid Microcapsules against Postharvest Blue Mold in Apple Fruit" Molecules 27, no. 22: 8108. https://doi.org/10.3390/molecules27228108
APA StyleWang, Y., Chen, J., Bian, W., Yang, X., Ye, L., He, S., & Song, X. (2022). Control Efficacy of Salicylic Acid Microcapsules against Postharvest Blue Mold in Apple Fruit. Molecules, 27(22), 8108. https://doi.org/10.3390/molecules27228108