
Immune-Enhancing Effect of Sargassum horneri on Cyclophosphamide-Induced Immunosuppression in BALB/c Mice and Primary Cultured Splenocytes


 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Results
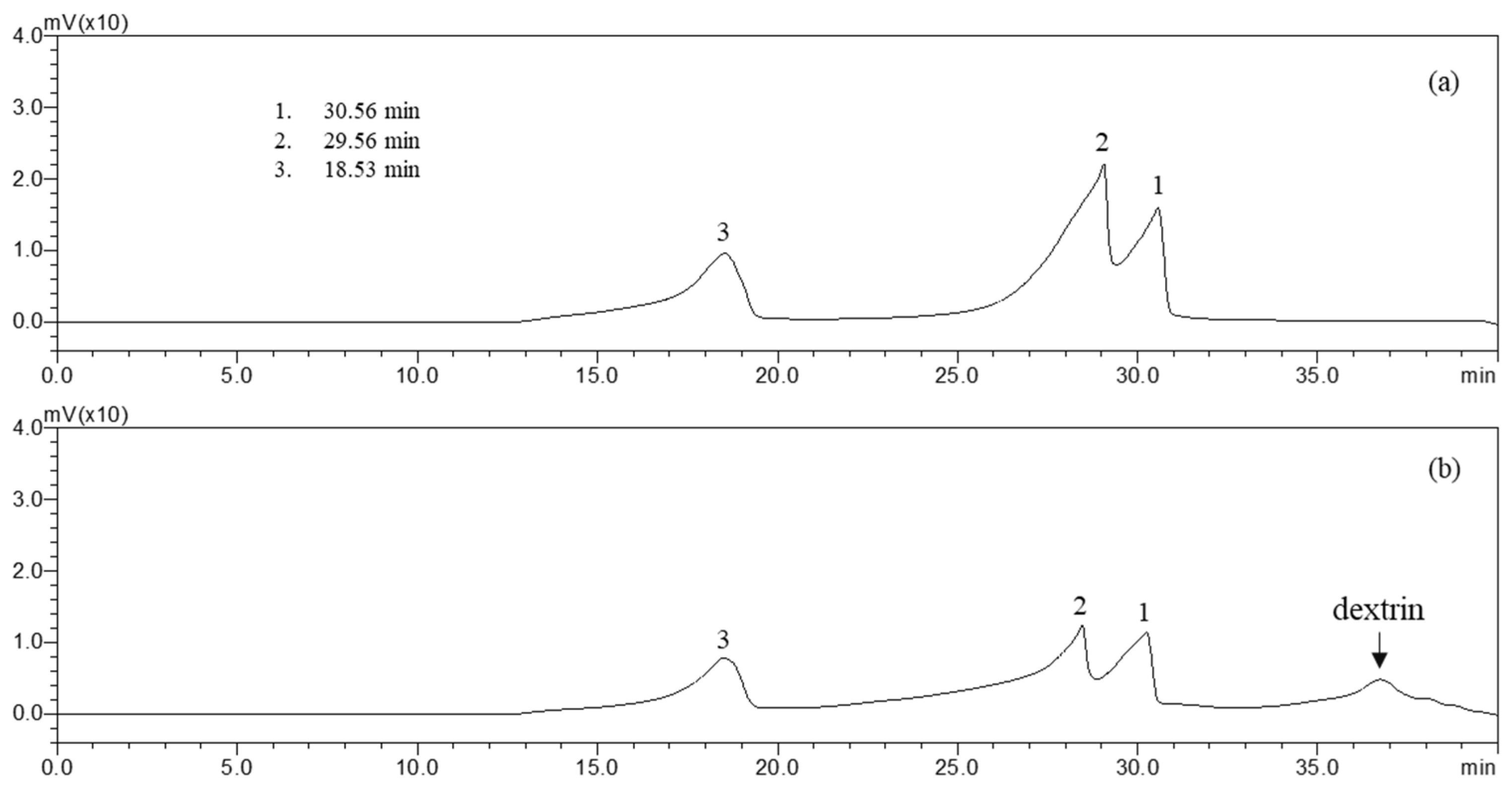
2.1. Total Sugar and Sulfate Group Contents of SH
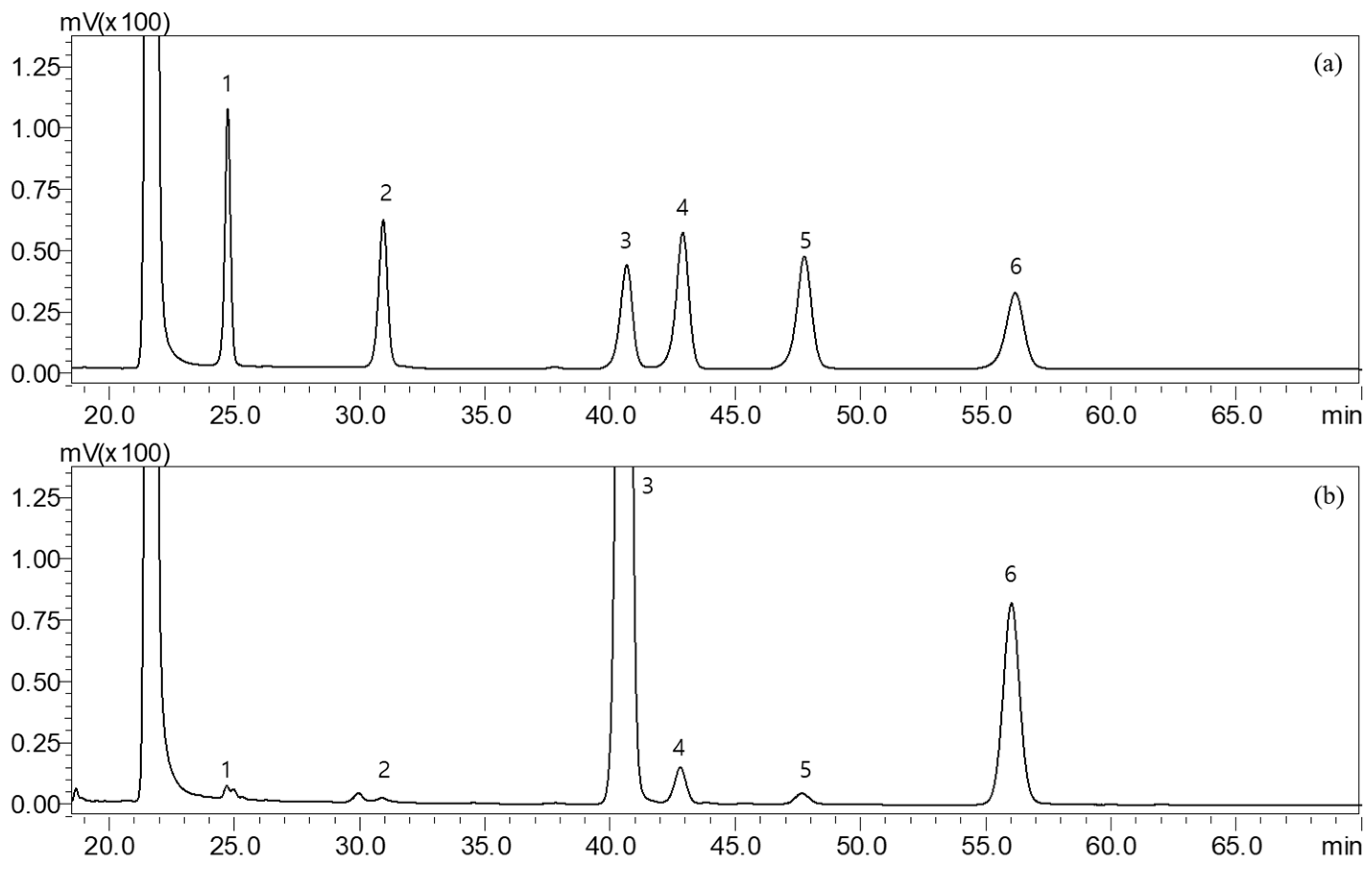
2.2. Monosaccharide Composition of SH
2.3. Molecular Weight Distribution
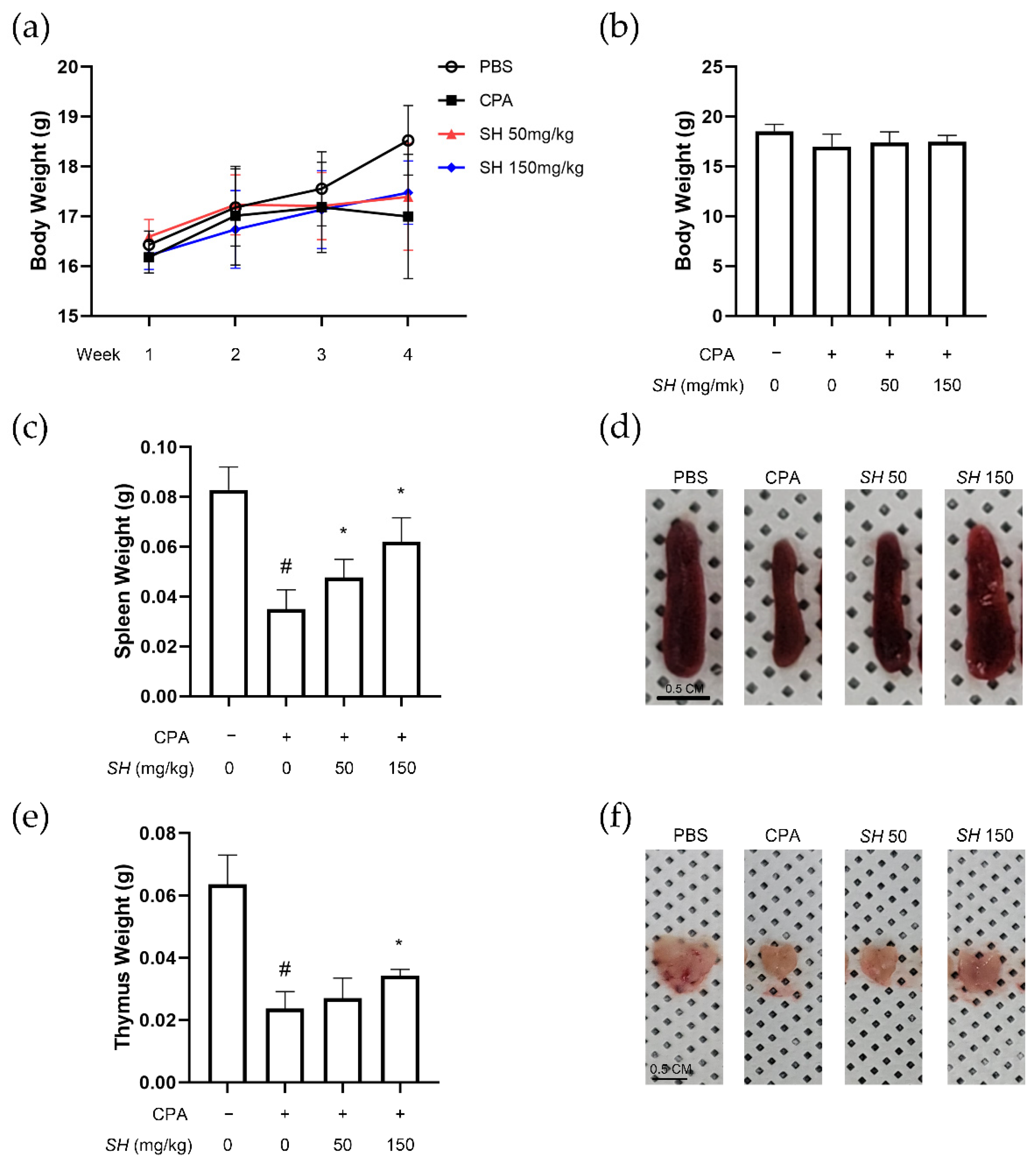
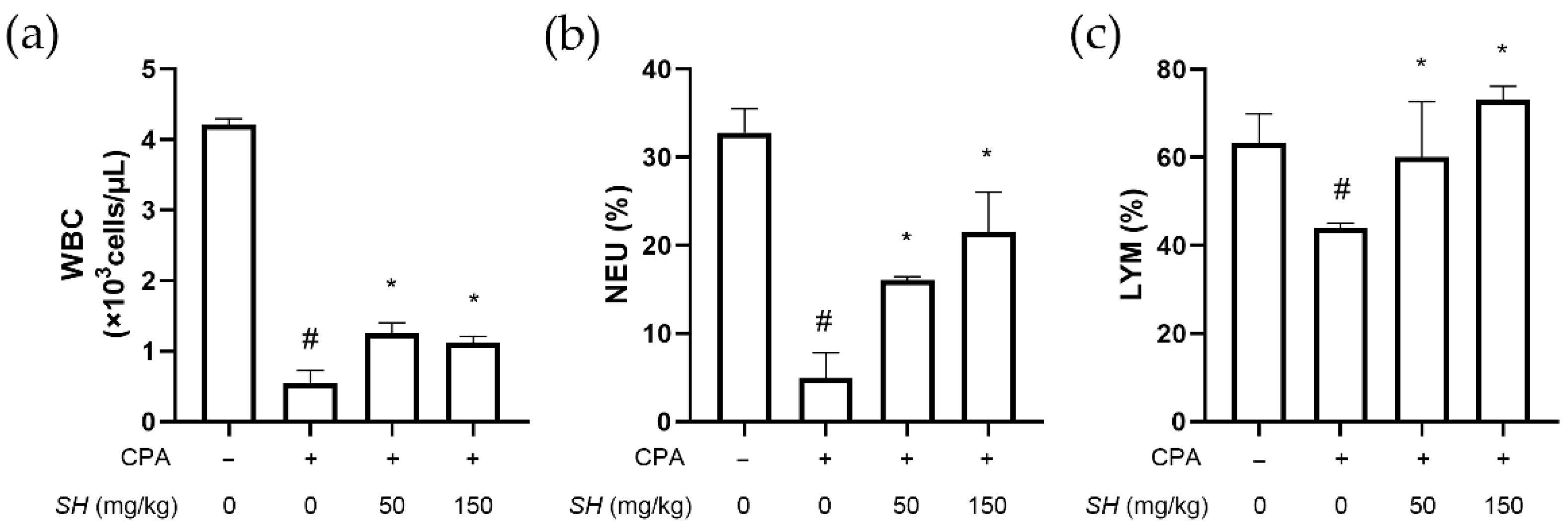
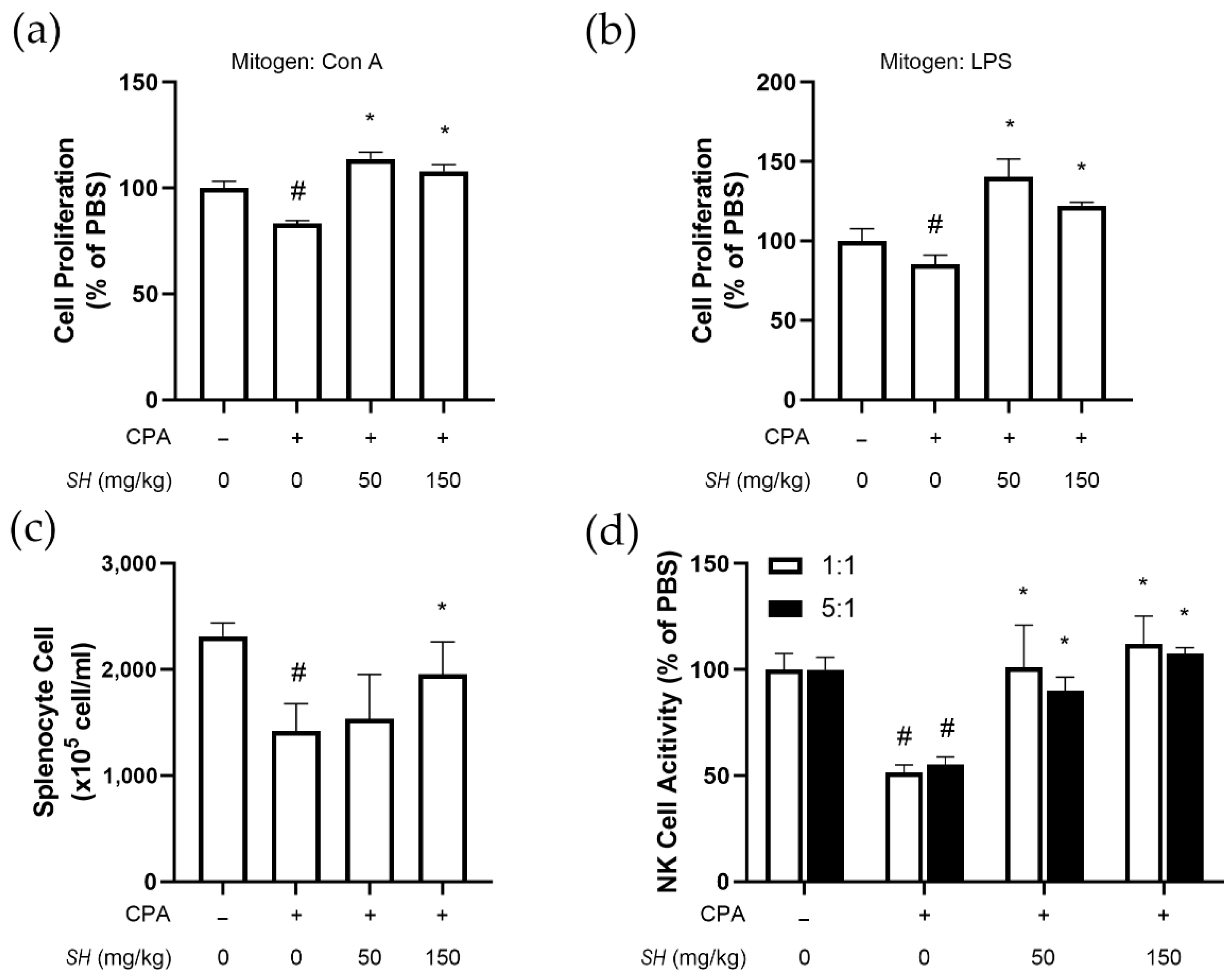
2.4. SH Enhances Immune Function in CPA-Induced Immunosuppressed BALB/c Mice
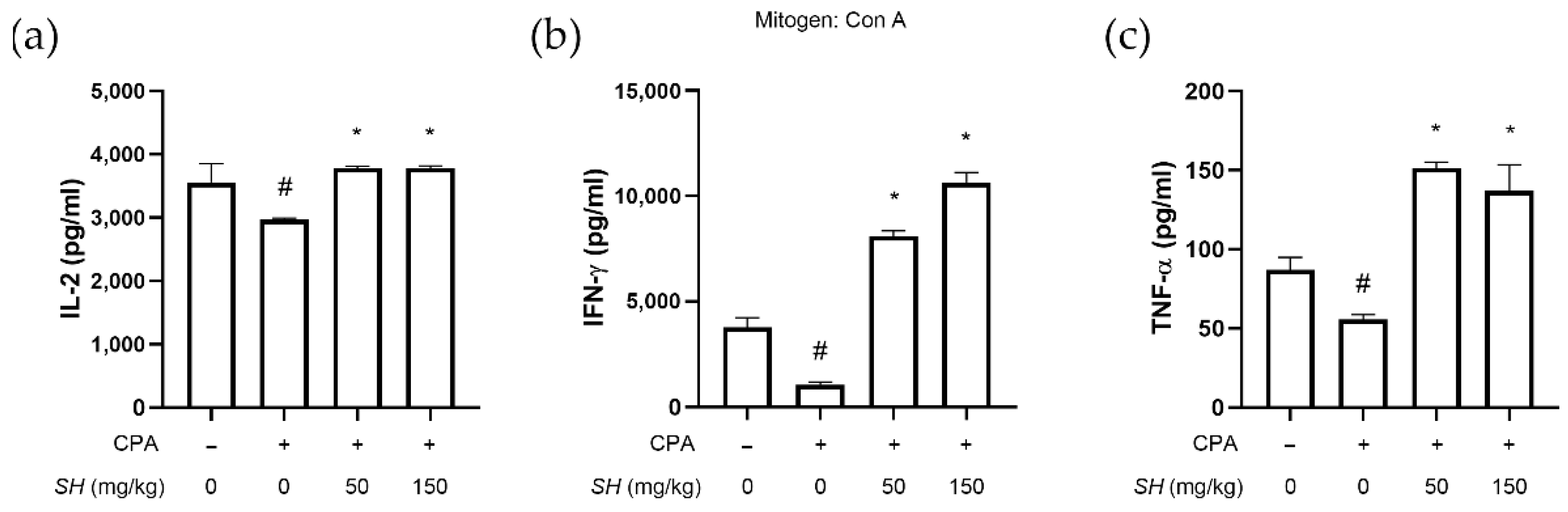
2.5. SH Induces Proliferation of Splenocytes, NK Cell Activity and Th1 Cytokine Expression In Vivo
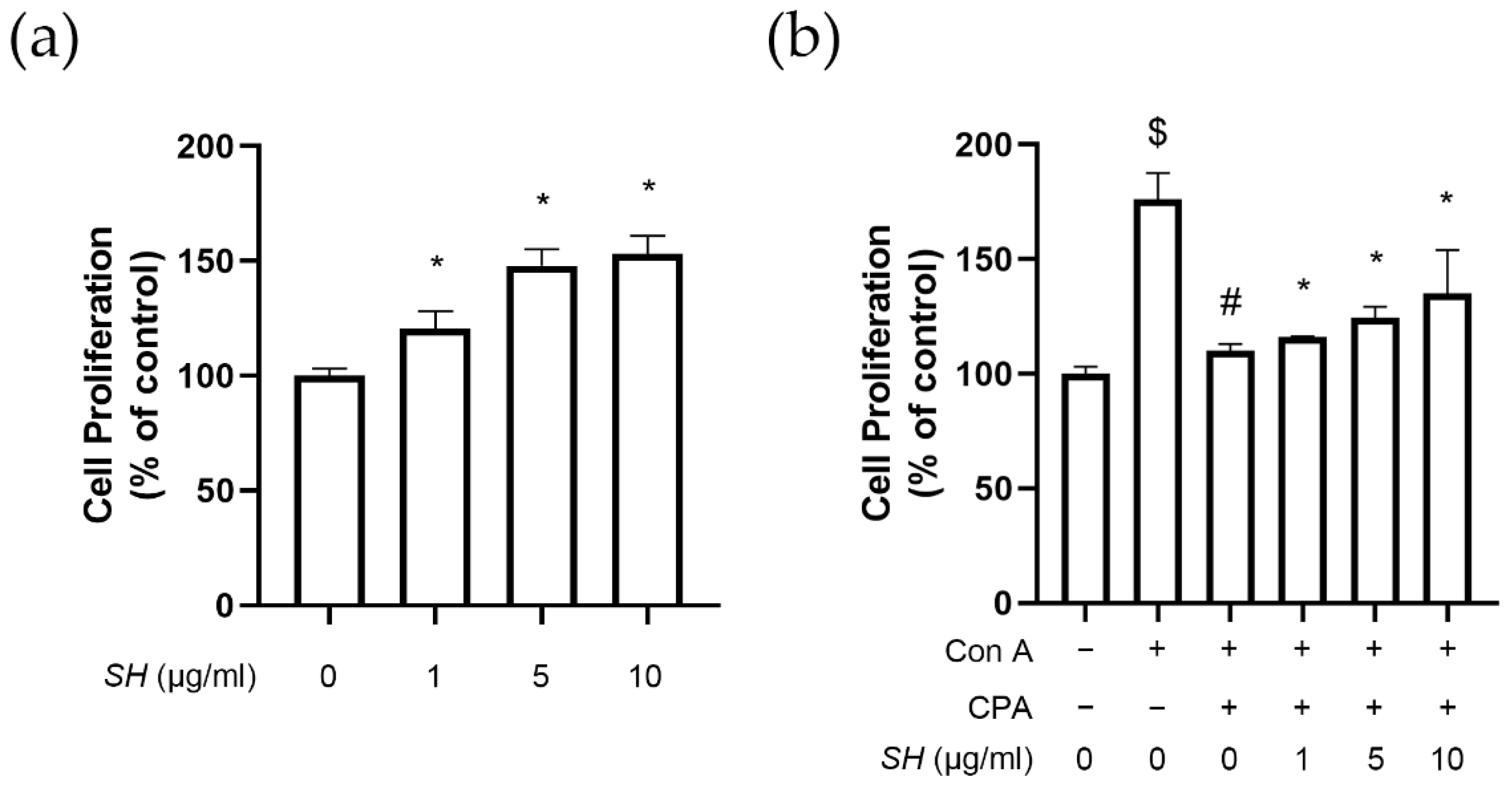
2.6. SH Induces Proliferation of Splenocytes
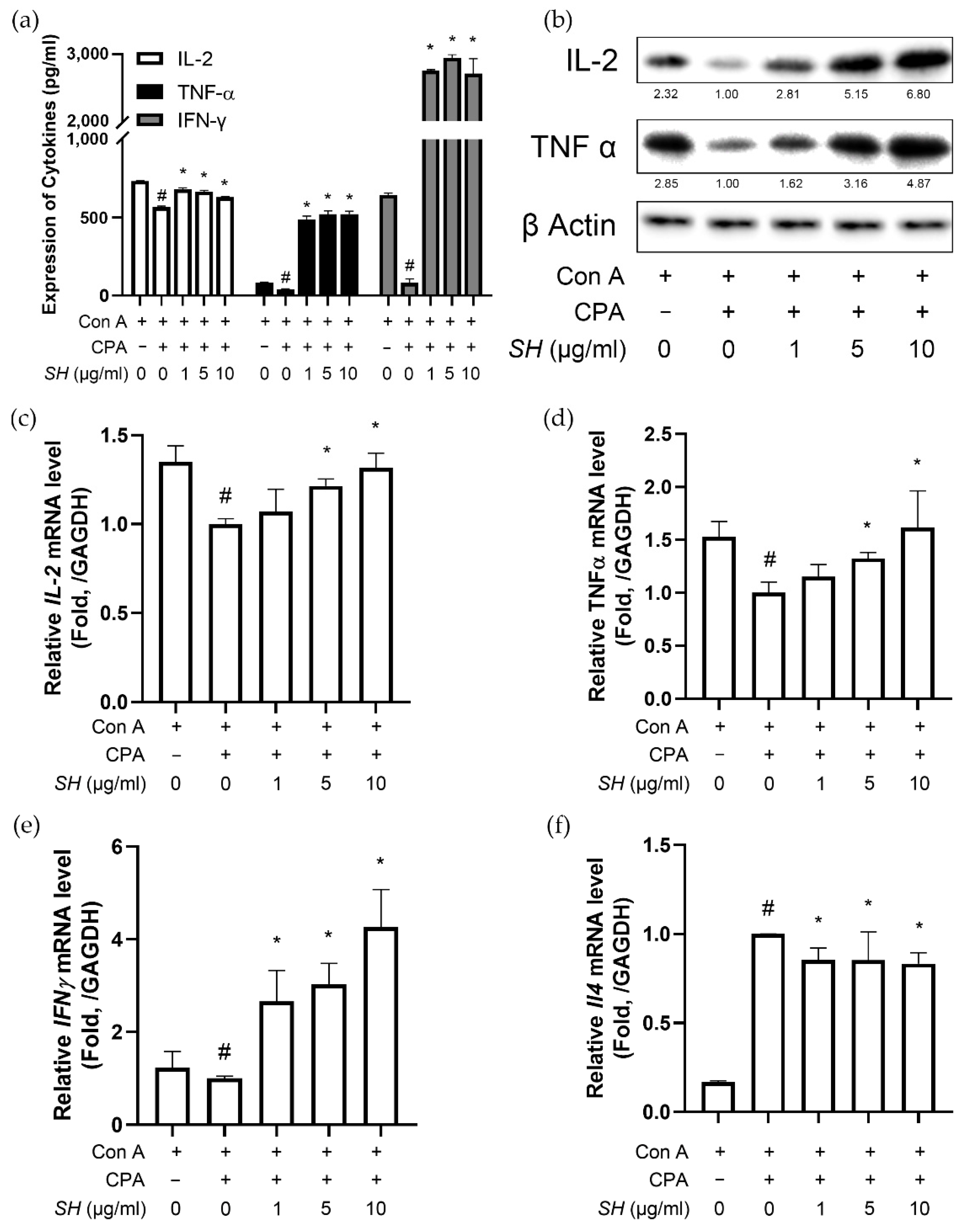
2.7. SH Increases Th1 Cytokine Expression in Splenocytes
3. Discussion
4. Materials and Methods
4.1. Sample Purchase and Extract Preparation
4.2. Measurement of Total Sugar Contents
4.3. Measurement of Sulfate Group Contents
4.4. Measurement of Monosaccharide Contents
4.5. HPLC-Diode Array Detector (DAD) Analysis
4.6. Molecular Weight Distribution
4.7. Animal Experiments
4.8. Isolation of Splenocytes
4.9. Splenocyte Cytotoxicity and Proliferation Assay
4.10. Flow Cytometry
4.11. Analysis of Blood Cell Count and Related Parameters
4.12. Determination of Cytokines
4.13. Determination of NK Activity
4.14. Western Blot Analysis
4.15. RNA Isolation and Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costa, M.; Cardoso, C.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Current knowledge and future perspectives of the use of seaweeds for livestock production and meat quality: A systematic review. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1075–1102. [Google Scholar] [CrossRef]
- Qiu, S.-M.; Aweya, J.J.; Liu, X.; Liu, Y.; Tang, S.; Zhang, W.; Cheong, K.-L. Bioactive polysaccharides from red seaweed as potent food supplements: A systematic review of their extraction, purification, and biological activities. Carbohydr. Polym. 2022, 275, 118696. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.J.; Zhao, R.X. Fucoidan: Structure and Bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Berteau, O. Sulfated fucans, fresh perspectives: Structures, functions, and biological properties of sulfated fucans and an overview of enzymes active toward this class of polysaccharide. Glycobiology 2003, 13, 29R–40R. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Shi, K.K.; Chen, S.; Wang, J.; Hassouna, A.; White, L.N.; Merien, F.; Xie, M.; Kong, Q.; Li, J.; et al. Fucoidan Extracted from the New Zealand Undaria pinnatifida—Physicochemical Comparison against Five Other Fucoidans: Unique Low Molecular Weight Fraction Bioactivity in Breast Cancer Cell Lines. Mar. Drugs 2018, 16, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.L.; Lee, B.-Y.; You, S.G. Relationship between Oversulfation and Conformation of Low and High Molecular Weight Fucoidans and Evaluation of Their in Vitro Anticancer Activity. Molecules 2010, 16, 291–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, P.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structural Features and Antiviral Activity of Sulphated Fucans from the Brown Seaweed Cystoseira Indica. Antivir. Chem. Chemother. 2007, 18, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Dinesh, S.; Menon, T.; Hanna, L.E.; Suresh, V.; Sathuvan, M.; Manikannan, M. In vitro anti-HIV-1 activity of fucoidan from Sargassum swartzii. Int. J. Biol. Macromol. 2016, 82, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Kawaguchi, M.; Kitamura, K.; Narumiya, S.; Kawamura, M.; Tengan, I.; Nishimoto, S.; Hanamure, Y.; Majima, Y.; Tsubura, S.; et al. An Exploratory Study on the Anti-inflammatory Effects of Fucoidan in Relation to Quality of Life in Advanced Cancer Patients. Integr. Cancer Ther. 2018, 17, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Xu, J.; Ge, K.; Tian, Q.; Zhao, P.; Guo, Y. Anti-inflammatory effect of low molecular weight fucoidan from Saccharina japonica on atherosclerosis in apoE-knockout mice. Int. J. Biol. Macromol. 2018, 118, 365–374. [Google Scholar] [CrossRef]
- Anisimova, N.Y.; Ustyuzhanina, N.E.; Donenko, F.V.; Bilan, M.I.; Ushakova, N.A.; Usov, A.I.; Nifantiev, N.E.; Kiselevskiy, M.V. Influence of fucoidans and their derivatives on antitumor and phagocytic activity of human blood leucocytes. Biochemistry 2015, 80, 925–933. [Google Scholar] [CrossRef]
- Hwang, P.-A.; Lin, H.-T.V.; Lin, H.-Y.; Lo, S.-K. Dietary Supplementation with Low-Molecular-Weight Fucoidan Enhances Innate and Adaptive Immune Responses and Protects against Mycoplasma pneumoniae Antigen Stimulation. Mar. Drugs 2019, 17, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decharneux, T.; Dubois, F.; Beauloye, C.; De Coninck, S.W.; Wattiaux, R. Effect of various flavonoids on lysosomes subjected to an oxidative or an osmotic stress. Biochem. Pharmacol. 1992, 44, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, M.; Liu, J.; Ding, X.; He, J.; Zhao, S.; Wu, L.; Gao, S.; Zhao, C.; Liu, D.; Zhang, J.; et al. Sargassum blooms in the East China Sea and Yellow Sea: Formation and management. Mar. Pollut. Bull. 2021, 162, 111845. [Google Scholar] [CrossRef]
- Byeon, S.; Cheon, K.-S.; Kim, S.; Yun, S.-H.; Oh, H.-J.; Park, S.; Kim, T.-H.; Kim, J.; Lee, H. Comparative Analysis of Sequence Polymorphism in Complete Organelle Genomes of the ‘Golden Tide’ Seaweed Sargassum horneri between Korean and Chinese Forms. Sustainability 2020, 12, 7280. [Google Scholar] [CrossRef]
- Liu, F.; Liu, X.; Wang, Y.; Jin, Z.; Moejes, F.W.; Sun, S. Insights on the Sargassum horneri golden tides in the Yellow Sea inferred from morphological and molecular data. Limnol. Oceanogr. 2018, 63, 1762–1773. [Google Scholar] [CrossRef]
- Lee, B.J.; Lee, S.-M.; Hyun, J.-h.; Kim, Y.Y. Durability Performances of Concrete Produced with Recycled Bio-Polymer Based on Sargassum Honeri. J. Korean Recycl. Constr. Resour. Inst. 2019, 7, 445–451. [Google Scholar]
- Madhavaraj, L.; Lim, H.-D.; Kim, K.-M.; Kim, D.-H.; Han, G.H. Influence of Sargassum horneri Mitigating Odorous Gas Emissions from Swine Manure Storage Facilities. Sustainability 2020, 12, 7587. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Lee, H.G.; Herath, K.H.I.N.M.; Jee, Y.; Jeon, Y.-J. The protective effect of Sargassum horneri against particulate matter-induced inflammation in lung tissues of an in vivo mouse asthma model. Food Funct. 2019, 10, 7995–8004. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, S.Y.; Lee, H.G.; Je, J.G.; Jee, Y.; Jeon, Y.J. Sargassum horneri (Turner) inhibit urban particulate matter-induced inflammation in MH-S lung macrophages via blocking TLRs mediated NF-κB and MAPK activation. J. Ethnopharmacol. 2020, 249, 112363. [Google Scholar] [CrossRef]
- Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, H.-S.; Jeon, Y.-J.; Jee, Y.; Kim, K.-N.; Lee, K.; Fernando, I.P.S.; Ahn, G. Sargassum horneri (Turner) C. Agardh ethanol extract attenuates fine dust-induced inflammatory responses and impaired skin barrier functions in HaCaT keratinocytes. J. Ethnopharmacol. 2021, 273, 114003. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Chen, X.; Sun, P. Chemical characterization, antioxidant and antitumor activity of sulfated polysaccharide from Sargassum horneri. Carbohydr. Polym. 2014, 105, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Liu, J.; Chen, X.; Fang, Z.; Sun, P. Structural features and antitumor activity of a purified polysaccharide extracted from Sargassum horneri. Int. J. Biol. Macromol. 2015, 73, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.; Lee, H.; Kim, N.; Jo, H.; Woo, E.-R.; Lee, K.; Han, Y.; Park, S.; Ahn, G.; Cheong, S.; et al. The Anti-Oxidative and Anti-Neuroinflammatory Effects of Sargassum horneri by Heme Oxygenase-1 Induction in BV2 and HT22 Cells. Antioxidants 2021, 10, 859. [Google Scholar] [CrossRef]
- Herath, K.H.I.N.M.; Cho, J.; Kim, A.; Kim, H.-S.; Han, E.J.; Kim, H.J.; Kim, M.S.; Ahn, G.; Jeon, Y.-J.; Jee, Y. Differential modulation of immune response and cytokine profiles of Sargassum horneri ethanol extract in murine spleen with or without Concanavalin A stimulation. Biomed. Pharmacother. 2018, 110, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Athukorala, Y.; Lee, K.-W.; Kim, S.-K.; Jeon, Y.-J. Anticoagulant activity of marine green and brown algae collected from Jeju Island in Korea. Bioresour. Technol. 2007, 98, 1711–1716. [Google Scholar] [CrossRef]
- Kim, D.-S.; Sung, N.-Y.; Park, S.-Y.; Kim, G.M.; Eom, J.; Yoo, J.-G.; Seo, I.-R.; Han, I.-J.; Cho, Y.-B.; Kim, K.-A. Immunomod-ulating activity of Sargassum horneri extracts in RAW264.7 macrophages. Korean J. Nutr. 2018, 51, 507–514. [Google Scholar]
- Yatim, K.M.; Lakkis, F.G. A Brief Journey through the Immune System. Clin. J. Am. Soc. Nephrol. 2015, 10, 1274–1281. [Google Scholar] [CrossRef] [Green Version]
- Gourbal, B.; Pinaud, S.; Beckers, G.J.M.; Van Der Meer, J.W.M.; Conrath, U.; Netea, M.G. Innate immune memory: An evolutionary perspective. Immunol. Rev. 2018, 283, 21–40. [Google Scholar] [CrossRef]
- Cooper, M.D.; Alder, M.N. The Evolution of Adaptive Immune Systems. Cell 2006, 124, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Cant, A.; Cole, T. Infections in the Immunocompromised. Adv. Exp. Med. Biol. 2010, 659, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.-H.; Park, K.-H.; Kim, S.-Y.; Oh, C.-S.; Bang, S.-I.; Chae, H.-J. Process Development for Deordorization of Fucoidan Using a Combined Method of Solvent Extraction and Spray Drying. KSBB J. 2011, 26, 49–56. [Google Scholar] [CrossRef]
- Lahaye, M. Marine algae as sources of fibres: Determination of soluble and insoluble dietary fibre contents in some ‘sea vegetables’. J. Sci. Food Agric. 1991, 54, 587–594. [Google Scholar] [CrossRef]
- Girard, J.P.; Marion, C.; Liutkus, M.; Boucard, M.; Rechencq, E.; Vidal, J.P.; Rossi, J.C. Hypotensive Constituents of Marine Algae; 1. Pharmacological Studies of Laminine. Planta Med. 1988, 54, 193–196. [Google Scholar] [CrossRef]
- Zayed, A.; El-Aasr, M.; Ibrahim, A.-R.S.; Ulber, R. Fucoidan Characterization: Determination of Purity and Physicochemical and Chemical Properties. Mar. Drugs 2020, 18, 571. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Y.; Zhang, Y.; Yang, Y.; Wang, P.; Imre, B.; Wong, A.C.Y.; Hsieh, Y.S.Y.; Wang, D. Brown Algae Carbohydrates: Structures, Pharmaceutical Properties, and Research Challenges. Mar. Drugs 2021, 19, 620. [Google Scholar] [CrossRef]
- Lahrsen, E.; Liewert, I.; Alban, S. Gradual degradation of fucoidan from Fucus vesiculosus and its effect on structure, antioxidant and antiproliferative activities. Carbohydr. Polym. 2018, 192, 208–216. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. A highly regular fraction of a fucoidan from the brown seaweed Fucus distichus L. Carbohydr. Res. 2004, 339, 511–517. [Google Scholar] [CrossRef]
- Fletcher, H.; Biller, P.; Ross, A.; Adams, J. The seasonal variation of fucoidan within three species of brown macroalgae. Algal Res. 2017, 22, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Duarte, M.E.; Cardoso, M.A.; Noseda, M.D.; Cerezo, A.S. Structural studies on fucoidans from the brown seaweed Sargassum stenophyllum. Carbohydr. Res. 2001, 333, 281–293. [Google Scholar] [CrossRef]
- Malek, T.R. The main function of IL-2 is to promote the development of T regulatory cells. J. Leukoc. Biol. 2003, 74, 961–965. [Google Scholar] [CrossRef]
- Castro, F.; Cardoso, A.P.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef]
- Parameswaran, N.; Patial, S. Tumor Necrosis Factor-α Signaling in Macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef]
- Qi, Q.; Dong, Z.; Sun, Y.; Li, S.; Zhao, Z. Protective Effect of Bergenin against Cyclophosphamide-Induced Immunosuppression by Immunomodulatory Effect and Antioxidation in Balb/c Mice. Molecules 2018, 23, 2668. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Grabowski, K.A.; Xin, J.-P.; Coleman, J.; Huang, Z.; Espiritu, B.; Alkan, S.; Xie, H.B.; Zhu, Y.; White, F.A.; et al. IL-4 Induces Differentiation and Expansion of Th2 Cytokine-Producing Eosinophils. J. Immunol. 2004, 172, 2059–2066. [Google Scholar] [CrossRef] [Green Version]
- Han, E.J.; Kim, H.-S.; Sanjeewa, K.K.A.; Jung, K.; Jee, Y.; Jeon, Y.-J.; Fernando, I.P.S.; Ahn, G. Sargassum horneri as a Functional Food Ameliorated IgE/BSA-Induced Mast Cell Activation and Passive Cutaneous Anaphylaxis in Mice. Mar. Drugs 2020, 18, 594. [Google Scholar] [CrossRef]
- Ponce, N.M.A.; Stortz, C.A. A Comprehensive and Comparative Analysis of the Fucoidan Compositional Data Across the Phaeophyceae. Front. Plant Sci. 2020, 11, 556312. [Google Scholar] [CrossRef] [PubMed]
- Han, E.J.; Fernando, I.P.S.; Kim, H.-S.; Jeon, Y.-J.; Madusanka, D.M.D.; Dias, M.K.H.M.; Jee, Y.; Ahn, G. Oral Administration of Sargassum horneri Improves the HDM/DNCB-Induced Atopic Dermatitis in NC/Nga Mice. Nutrients 2020, 12, 2482. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, H.J.; Jee, Y.; Jeon, Y.-J.; Kim, H.J. Antioxidant potential of Sargassum horneri extract against urban particulate matter-induced oxidation. Food Sci. Biotechnol. 2020, 29, 855–865. [Google Scholar] [CrossRef]
- Murakami, S.; Hirazawa, C.; Ohya, T.; Yoshikawa, R.; Mizutani, T.; Ma, N.; Moriyama, M.; Ito, T.; Matsuzaki, C. The Edible Brown Seaweed Sargassum horneri (Turner) C. Agardh Ameliorates High-Fat Diet-Induced Obesity, Diabetes, and Hepatic Steatosis in Mice. Nutrients 2021, 13, 551. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; Jones, R.J.; Brodsky, R.A. Cyclophosphamide and cancer: Golden anniversary. Nat. Rev. Clin. Oncol. 2009, 6, 638–647. [Google Scholar] [CrossRef]
- Ahlmann, M.; Hempel, G. The effect of cyclophosphamide on the immune system: Implications for clinical cancer therapy. Cancer Chemother. Pharmacol. 2016, 78, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Swan, D.; Gurney, M.; Krawczyk, J.; Ryan, A.E.; O’Dwyer, M. Beyond DNA Damage: Exploring the Immunomodulatory Effects of Cyclophosphamide in Multiple Myeloma. HemaSphere 2020, 4, e350. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.-W.; Myers, R.; Greenslade, A.; Mader, E.K.; Greiner, S.; Federspiel, M.J.; Dispenzieri, A.; Russell, S.J. Using clinically approved cyclophosphamide regimens to control the humoral immune response to oncolytic viruses. Gene Ther. 2013, 20, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pukhalsky, A.; Toptygina, A.P.; Viktorov, V. Immunosuppressive action of cyclophosphamide in mice: Contribution of some factors to determination of strain differences. Int. J. Immunopharmacol. 1993, 15, 509–514. [Google Scholar] [CrossRef]
- Sharma, B.; Vaziri, N.D. Augmentation of human natural killer cell activity by cyclophosphamide in vitro. Cancer Res. 1984, 44, 3258–3261. [Google Scholar]
- Paul, S.; Lal, G. The Molecular Mechanism of Natural Killer Cells Function and Its Importance in Cancer Immunotherapy. Front. Immunol. 2017, 8, 1124. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, J.; Boisson-Dupuis, S.; Jouanguy, E.; Picard, C.; Puel, A.; Abel, L.; Casanova, J.-L. Novel primary immunodeficiencies revealed by the investigation of paediatric infectious diseases. Curr. Opin. Immunol. 2008, 20, 39–48. [Google Scholar] [CrossRef]
- Iannello, A.; Raulet, D.H. Immune Surveillance of Unhealthy Cells by Natural Killer Cells. Cold Spring Harb. Symp. Quant. Biol. 2013, 78, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Paust, S.; Gill, H.S.; Wang, B.-Z.; Flynn, M.P.; Moseman, E.A.; Senman, B.; Szczepanik, M.; Telenti, A.; Askenase, P.W.; Compans, R.; et al. Critical role for the chemokine receptor CXCR6 in NK cell–mediated antigen-specific memory of haptens and viruses. Nat. Immunol. 2010, 11, 1127–1135. [Google Scholar] [CrossRef]
- Bevington, S.L.; Cauchy, P.; Withers, D.R.; Lane, P.J.L.; Cockerill, P.N. T Cell Receptor and Cytokine Signaling Can Function at Different Stages to Establish and Maintain Transcriptional Memory and Enable T Helper Cell Differentiation. Front. Immunol. 2017, 8, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, D.; Zheng, L.; Lin, J.; Zhang, B.; Zhu, Y.; Li, N.; Xie, S.; Wang, Y.; Gao, N.; Huang, Z. Structural basis of assembly of the human T cell receptor–CD3 complex. Nature 2019, 573, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic self-tolerance maintained by activated T cells ex-pressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar] [PubMed]
- Kronenberg, M.; Davis, M.M.; Early, P.W.; Hood, L.E.; Watson, J.D. Helper and killer T cells do not express B cell immunoglobulin joining and constant region gene segments. J. Exp. Med. 1980, 152, 1745–1761. [Google Scholar] [CrossRef] [Green Version]
- Berger, A. Science commentary: Th1 and Th2 responses: What are they? BMJ 2000, 321, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, A.; Vargas, J.A.; Castejón, R.; Citores, M.J.; Romero, Y.; Millán, I.; Durántez, A. Production of intracellular IL-2, TNF-α, and IFN-γ by T cells in B-CLL. Cytom. Part B Clin. Cytom. 2003, 56B, 23–29. [Google Scholar] [CrossRef]
- Ruterbusch, M.; Pruner, K.B.; Shehata, L.; Pepper, M. In Vivo CD4+ T Cell Differentiation and Function: Revisiting the Th1/Th2 Paradigm. Annu. Rev. Immunol. 2020, 38, 705–725. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Dodgson, K.S. Determination of inorganic sulphate in studies on the enzymic and non-enzymic hydrolysis of carbohydrate and other sulphate esters. Biochem. J. 1961, 78, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.H.; Lee, M.J.; Moon, S.J.; Kim, D.S.; Nho, E.Y.; Choi, J.Y.; Kim, K.S. Analysis of monosaccharide composition of fu-coidan from Sargassum horneri using UPLC-DAD-TOF/MS combined with precolumn derivatization by 3-methyl-1-phenyl-5-pyrazolone. Korean J. Food Preserv. 2019, 26, 219–227. [Google Scholar] [CrossRef]
- Zhou, J.-R.; Yamada, R.; Huruiti, E.; Kitahara, N.; Nakamura, H.; Fang, J.; Nohara, T.; Yokomizo, K. Ripe Tomato Saponin Esculeoside A and Sapogenol Esculeogenin A Suppress CD4+ T Lymphocyte Activation by Modulation of Th2/Th1/Treg Differentiation. Nutrients 2022, 14, 2021. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Park, J.; Kim, H.; Sim, J.; Youn, D.; Kang, J.; Lim, S.; Jeong, M.; Yang, W.M.; Lee, S.; et al. Vanillic acid attenuates obesity via activation of the AMPK pathway and thermogenic factors in vivo and in vitro. FASEB J. 2018, 32, 1388–1402. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | Contents (mg/g Dry Weight) |
|---|---|---|
| 1 | Mannose | 1.91 |
| 2 | Rhamnose | 2.13 |
| 3 | Glucose | - |
| 4 | Galactose | 8.10 |
| 5 | Xylose | 4.29 |
| 6 | Fucose | 59.49 |
| PBS | CPA | SH 50 | SH 150 | |
|---|---|---|---|---|
| RBC (×106 cells/μL) | 11.15 ± 0.22 | 9.66 ± 0.23 # | 10.50 ± 0.23 | 11.60 ± 0.17 * |
| HGB (g/dL) | 16.6 ± 0.28 | 14.38 ± 0.33 # | 15.6 ± 0.35 | 17.48 ± 0.25 * |
| HCT (%) | 52.28 ± 1.04 | 43.93 ± 1.04 # | 48.70 ± 1.09 | 53.85 ± 0.85 * |
| MCV (fL) | 46.92 ± 0.31 | 45.48 ± 0.14 | 46.38 ± 0.3 | 46.43 ± 0.38 |
| MCH (pg) | 14.90 ± 0.05 | 14.88 ± 0.01 | 14.85 ± 0.07 | 15.08 ± 0.07 |
| MCHC (g/dL) | 31.77 ± 0.18 | 32.75 ± 0.09 | 32.02 ± 0.15 | 32.48 ± 0.13 |
| Monocytes (%) | 2 ± 0.20 | 15.35 ± 1.4 # | 11.92 ± 1.3 | 4.98 ± 1.2 * |
| Basophils (%) | 0.06 ± 0.04 | 1.34 ± 0.21 # | 1.72 ± 0.45 | 0.53 ± 0.25* |
| Eosinophils (%) | 13.13 ± 0.71 | 30.83 ± 3.09 # | 3.46 ± 0.54 * | 3.38 ± 0.79* |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.I.; Kim, D.-S.; Jung, Y.; Sung, N.-Y.; Kim, M.; Han, I.-J.; Nho, E.Y.; Hong, J.H.; Lee, J.-K.; Boo, M.; et al. Immune-Enhancing Effect of Sargassum horneri on Cyclophosphamide-Induced Immunosuppression in BALB/c Mice and Primary Cultured Splenocytes. Molecules 2022, 27, 8253. https://doi.org/10.3390/molecules27238253
Kim HI, Kim D-S, Jung Y, Sung N-Y, Kim M, Han I-J, Nho EY, Hong JH, Lee J-K, Boo M, et al. Immune-Enhancing Effect of Sargassum horneri on Cyclophosphamide-Induced Immunosuppression in BALB/c Mice and Primary Cultured Splenocytes. Molecules. 2022; 27(23):8253. https://doi.org/10.3390/molecules27238253
Chicago/Turabian StyleKim, Hyo In, Dong-Sub Kim, Yunu Jung, Nak-Yun Sung, Minjee Kim, In-Jun Han, Eun Yeong Nho, Joon Ho Hong, Jin-Kyu Lee, Mina Boo, and et al. 2022. "Immune-Enhancing Effect of Sargassum horneri on Cyclophosphamide-Induced Immunosuppression in BALB/c Mice and Primary Cultured Splenocytes" Molecules 27, no. 23: 8253. https://doi.org/10.3390/molecules27238253
APA StyleKim, H. I., Kim, D. -S., Jung, Y., Sung, N. -Y., Kim, M., Han, I. -J., Nho, E. Y., Hong, J. H., Lee, J. -K., Boo, M., Kim, H. -L., Baik, S., Jung, K. O., Lee, S., Kim, C. S., & Park, J. (2022). Immune-Enhancing Effect of Sargassum horneri on Cyclophosphamide-Induced Immunosuppression in BALB/c Mice and Primary Cultured Splenocytes. Molecules, 27(23), 8253. https://doi.org/10.3390/molecules27238253